岩溶生态系统中的植物
2022-11-30曹建华袁道先朱同彬梁建宏周孟霞罗劬侃
曹建华 ,袁道先 ,杨 慧 ,黄 芬 ,朱同彬 ,梁建宏 ,周孟霞 ,罗劬侃 ,吴 夏
(1. 中国地质科学院岩溶地质研究所/自然资源部、广西岩溶动力学重点实验室, 广西 桂林 541004;2. 联合国教科文组织国际岩溶研究中心, 广西 桂林 541004)
0 引 言
岩溶生态系统是受岩溶环境制约的生态系统,支撑的岩石是碳酸盐岩,其由无机环境和生命系统两个部分组成。无机环境有两个特点:一是碳酸盐岩的富钙性,成土物质先天不足,土壤资源短缺、富钙偏碱性;二是地表、地下双层结构,水资源以地下水资源为主。而无机环境的两个特征诱发生命系统的两个特征:一是旱生、石生和喜钙的岩溶植被;二是无光、潮湿,在相对恒温条件下的地下生物群[1-2]。碳酸盐岩的化学组成和岩溶地质结构深刻影响岩溶生态系统的生产者、消费者和分解者,影响养分、水分的迁移和转化,具明显的地质印迹[3-4]。
生态系统是指一个生物群落及其周边地质地理环境相互作用的自然系统,生态系统由无机环境、生产者、消费者和分解者4 部分组成,生物群落适应周边环境而生存、发育和演化[5]。植物是生态系统的生产者,对维持生态系统的物质循环、群落演替起到至关重要作用,长期进化和自然选择产生的岩溶植被独具特色[6]。本文从岩溶生态系统中植物的喜钙性、石生性、旱生性、养分生态计量、多样性和特有性、生态系统碳汇功能的协同性等方面进行论述,最后介绍两种特有的天坑植物群落和洞口植物群落,以期深入认识岩溶植被特点、形成机制,对脆弱岩溶生态系统的保护和修复、功能的发挥起到促进作用[7]。
1 岩溶生态系统植物的喜钙性
岩溶生态系统的富钙属性,起源于碳酸盐岩的富钙性,虽然不同地质时代的碳酸盐岩化学组分有一定差异(其CaO 的含量为27.3%~54.33%,MgO 的含量为0.49%~19.66%,酸不溶物的含量为0.41%~10.53%)[8],但与玄武岩、花岗岩相比具有一个共同特征,那就是钙、镁含量高,在碳酸盐岩上孕育的岩溶生态系统中,无论是大气降雨、河水-泉水-地下水、土壤和植物体中,其钙含量均为硅酸盐岩上孕育的地带性生态系统类型中的几倍至十几倍[9-11]。
钙在植物生长发育的需求上为大量元素,但在细胞水平上属于微量元素[12]。植物体内钙的含量为0.1%~0.5%,大部分存在于细胞壁上,Ca2+对细胞壁的稳定性、细胞的完整性,以及对细胞反应中的调控作用,在植物个体生长发育过程中起着不可替代的作用[13-15]。
植 物 细 胞 外的Ca2+是mmol·L-1水 平,细 胞 内Ca2+约是μmol·L-1水平或更低,细胞内的钙主要分布在液泡中,细胞质中钙含量较少(以防钙与磷酸形成磷酸钙沉淀),其次是细胞器(线粒体、内质网和叶绿体),这些钙素处于动态平衡状态,以调节细胞外的离子环境与质膜的结构与功能[13-17]。
根据植物生长对土壤钙质的依赖程度,可定性地将植物划分为5 类:①嗜钙植物(Calciphile):只分布在富含碳酸钙的基质上;②喜钙植物(Calcicole):在多钙土壤上才能生长正常,较少生长在酸性土壤;③嫌钙植物(Calcifuge):在酸性土壤生长良好,土中含钙稍多即受毒害作用;④亚嫌钙性植物:在酸性土壤上生长茂盛,很少见于石灰岩或强钙性土壤上;⑤中间型(不择土性植物):对土壤含钙量多少没有明显反应[18-19]。根据调查资料的系统整理,依据《中国植被志》的分类原则,在群系尺度上编制中国岩溶植被分类系统,可分为:3 个植被型组、13 个植被型和554 个群系[7]。
贵州茂兰自然保护区是亚热带岩溶森林保护区,是研究岩溶植被群落特征理想的野外场所,其森林群落结构为常绿落叶阔叶林,其不同于亚热带常绿阔叶林,异于北亚热带和亚热带中常绿落叶混交林[20],采集岩溶区、非岩溶区植物品种各13 种,其中岩溶区与非岩溶区共有品种7 种中间型植物。通过测定其植物叶片的钙总量、不同钙素形态含量、亚细胞组分钙含量及根际土土壤钙含量和pH 等多项指标,揭示喜钙植物、嫌钙植物和中间植物品种间的定量差异性(表1)。
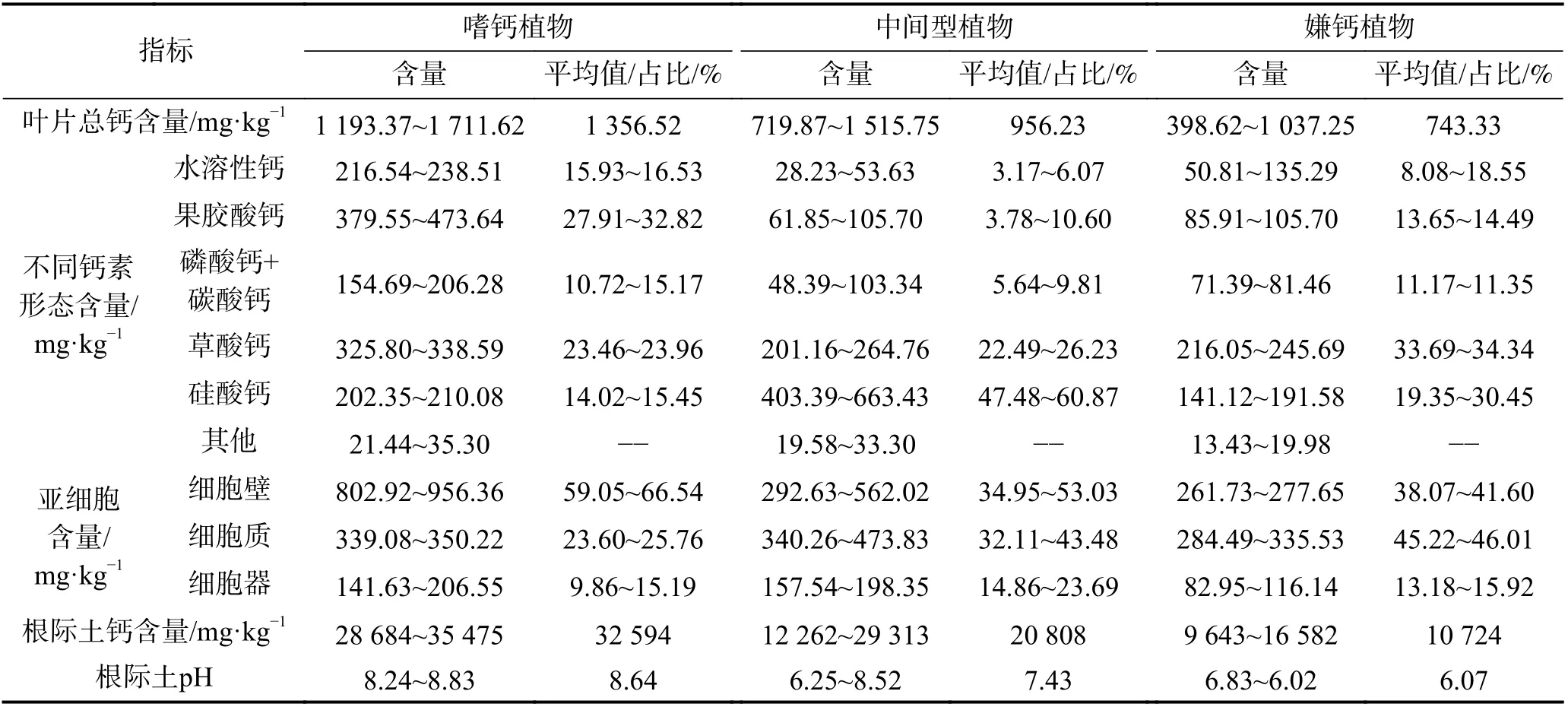
表1 嗜钙植物、嫌钙植物和中间植物与钙相关指标的对比[15]Table 1 Comparison of calcium-related indexes among Calciphile, Calcifuge and intermediate plants
嗜钙植物叶片钙质含量平均为1 356.52 mg·kg-1,比嫌钙植物高出82.49%;嗜钙植物叶片中钙以果胶酸钙形态为主,其含量占总钙量的27.91%~32.82%,其次是草酸钙(23.46%~23.96%),嫌钙植物叶片中钙主要以草酸钙为主,其含量占总钙量的33.69%~34.34%,其次是硅酸钙(19.35%~30.45%);嗜钙植物叶片中钙主要赋存在细胞壁,其含量占总钙量59.05%~66.54%,其次是细胞质(23.60%~25.76%),嫌钙植物叶片中钙主要赋存在细胞质中,其含量占总钙量45.22%~46.01%,其次是细胞壁(38.07%~41.60%)。
嗜钙植物叶片中果胶酸钙、草酸钙是两大主要形态,果胶酸钙是活性钙,有利于钙离子的迁移、转化和吸收,在提高细胞壁刚性,维持细胞膜的稳定性方面具有重要作用[21-22],草酸钙是惰性钙,可在液胞内形成异晶细胞,也可在细胞间隙形成针状、柱状等各种形状的晶体沉积[23],这与高钙岩溶环境和嗜钙植物生理特性是吻合的,有利于植被吸收大量的钙、有利于调控细胞内外钙素的平衡关系。嫌钙型植物叶片中主要以草酸钙、硅酸钙为主,草酸钙、硅酸钙均为惰性钙,难以转移、吸收和再利用,这或许就是嫌钙型植物缓解过量钙产生毒害的调节策略。
根据已有研究结果揭示,岩溶植物对高钙岩溶环境的适应策略主要包括耐受(容忍)、排除和阻断。耐受机制是植物细胞间隙存在相对较高的游离羧基,具有较高的阳离子交换量,使得植物可容纳较高含量的钙量[23];排除机制有两方面,一是提高钙调蛋白酶的活性,如细胞质膜上Ca2+-ATPase(腺苷三磷酸酶)活性增强,能将细胞质中的钙运送出细胞,在细胞壁中储存[12,24],二是钙通道的特异性,形成泌钙腺体、甚至气孔[25-26],将多余的钙素排出细胞外[27-28];阻断机制是指植物将从土壤中吸收的过量钙固定在根际周围,形成钙化根,限制其向地上部运输[20,29]。
由于岩溶生态系统中的钙库巨大,影响着岩溶生态系统的系列特征。土壤钙对有害重金属存在拮抗作用,酸沉降可缓解有害金属的影响,有利于维持生态系统的健康[30];钙素对岩溶生态系统中的养分循环具有强烈影响,土壤钙甚至被认为对岩溶生态系统退化过程具有指示作用[31]。例如,美国南部亚特兰大(Atlanta)州Panola 山区森林生态系统中的钙每年流失量为12.76 kg·ha-1,以此速度80 a 后,森林生态系统的钙营养将失衡。
2 岩溶生态系统中植物的石生性
岩溶生态系统被认为是生长在碳酸盐岩之上的生态系统[11],这意味着岩溶植物群落系统具有发达、丰富的地下生物量以支撑地上生物体。贵州茂兰自然保护区岩溶植被的根系生物量调查结果显示:地上生物量比同纬度的非岩溶区森林低,仅相当于温带森林区;但地下生物量则不仅比同纬度非岩溶区森林的高,而且比温带、热带非岩溶区森林的都高,达57.49~58.15 mg·hm-2,绝对量是温带、热带非岩溶区森林的近2 倍[32]。
植物根系越发达,从土壤、岩石深层吸收的水分和养分越多,越能适应干旱和贫瘠环境[32-33]。低等植物壳状地衣是常见的殖居在碳酸盐岩浅表层的先锋植物,它是藻类和真菌的结合体,藻类主要生存在最表层1 mm,通过光合作用向地衣提供养分,真菌则可在浅表层1~2 cm 内生存,汲取更多水分,互利共生。而地衣殖居可使碳酸盐岩风化速率增强24%~64%[34],同时提高岩石浅表层的持水率和岩石表面养分的富集程度,在改造微生境的同时,诱导高等植物萌发[34]。
高等植物群落具有相似的功能性状。通过对亚热带桂林岩溶区16 个洞穴调查,可观察到植物的根系沿岩隙或湿润的岩壁下延达数十米深,采集81 个洞穴植物根系样本,经DNA 测序表明根系样品分属38 个植物品种,其中有12 种植物根系下扎深度大于10 m。而黄连木(Pistacia chinensisBunge)、乌桕(Sapium sebiferum(Linn.) Roxb.)拥有最长根,长度可达29 m[35-36]。深根系植物可利用岩石裂隙水或地下河水,而浅根系植物主要利用表层带水和土壤水;在同一群落中植物根系结构和分布差异对水分吸收和再分配具有重要作用,存在互利共存机制[37],从而维持自然生态系统中物种多样性和群落稳定性。
岩溶植物的石生性还表现在岩石类型、构造特征对植被群落结构组成的强烈影响上。因钙、镁含量的差异性,碳酸盐岩可分为石灰岩、白云岩两大类,在溶解风化过程中,碳酸盐岩90%以上的物质会随水带走,残留下来形成土壤的一般在4%~10%,因岩石钙、镁和酸不溶物含量的小量差异,在形成土壤过程中得到了重组和放大,对植物群落组成产生明显影响[38]。贵州荔波县茂兰岩溶森林区4 000 m2范围内共记录到维管植物84 科180 属344 种:纯灰岩区主要以槭树科、樟科、胡桃科为主,十个重要值最大的科占总重要值的60.69%;纯白云岩区则以榛科、壳斗科、槭树科和山茱萸科为主,十个重要值最大的科占总重要值的66.10%;不纯碳酸盐岩区以漆树科、壳斗科、樟科为主[39](表2)。
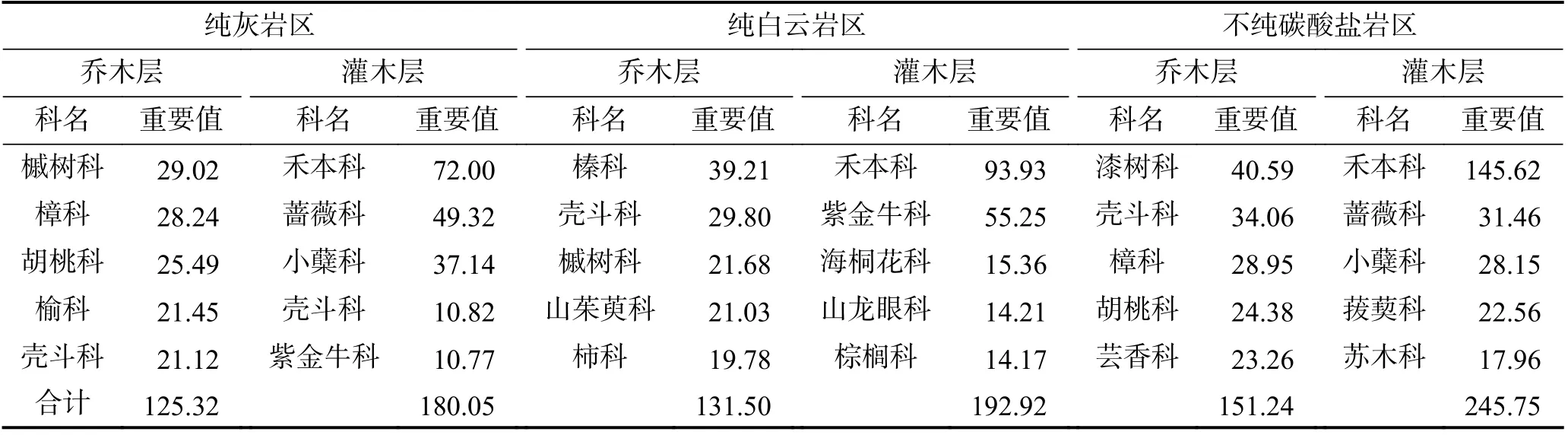
表2 贵州荔波县茂兰岩溶森林区群落乔木层和灌木层主要科的重要值[39]Table 2 Important values of main families in arbor layer and shrub layer in Maolan karst forest area, Libo county, Guizhou Province
石灰岩的矿物组分为斜六边体的方解石,白云岩的矿物组分是正六面体的白云石,石灰岩更易溶解风化、更易形成溶蚀裂隙和管道,且向地下具有更大的延伸,趋于不均匀风化,而岩石深部的溶蚀裂隙可为木本植物根系的发育提供更大的物理空间,获取深层地下水;白云岩抗溶解风化能力较强,趋于均匀风化,地表的土壤更趋均匀覆盖,有利于浅根系植物的发育。贵州省普定县调查统计结果显示:石灰岩为主的地区木本植物覆盖度高、植被活动强, 但生产力水平较低;而白云岩为主的地区草本植物覆盖度高、植被活动弱,但生产力水平较高[40]。
3 岩溶生态系统中植物的旱生性
中国南方岩溶生态系统的干旱是岩溶水文地质结构性干旱。岩溶含水介质在垂向上可依次发育为:岩溶表层带、包气垂直渗流带、季节波动径流带、饱水潜流带[41]。表层岩溶带是地表强烈岩溶作用过程的产物,表层岩溶带溶隙率、孔隙率可达20%~60%,厚度多在几米至十几米[42-43]。表层岩溶带的密集溶隙率、孔隙率,加上竖井、落水洞、漏斗等的发育导致降雨迅速垂直入渗、汇入地下,很少产生坡面径流[44]。桂林岩溶区能产生坡面流的最低日降雨量为64 mm,全年坡面径流量约占流域年径流量的1%,表层带径流量约占流域年径流量的8%[45]。
地下含水空间以溶蚀裂隙(含孔隙)为主,岩溶洞穴或岩溶管道主要起传输作用,如贵州普定后寨地下河流域六谷、老黑潭等水文站流量实测资料进行的统测分析显示,六谷站汇水范围的岩溶管道容积占储水空间的7.47%~9.40%,而溶蚀裂隙的容积占92.53%;老黑潭站汇水范围的岩溶管道占储水空间的7.49%~11.60%,溶蚀裂隙的容积占88.4%~92.5%[46]。
而水是限制植物生长和分布的最主要因素[31,47]。地下水丰富、地表水短缺,成为岩溶区植物在旱季生态需水量不足的地质原因。岩溶区的植物主要通过生态、生理的调控来适应岩溶干旱。
植物通过对生态性状的调控,以提高适应岩溶区干旱能力。一是植物根(系)生态性状的调控,延长根(系)长度和量,获取更深层的水分[33];二是植物木质部生态性状的调控,木质部主要由导管、薄壁组织和纤维组成,导管输送水分、薄壁组织储存水分、纤维机械支撑[48],缩小导管直径、丰富薄壁组织,能够在缺水环境下维持水分运输,从而维持叶片和枝条的生理功能作用[49-50],甚至有些岩溶植物(如薄叶羊蹄甲Bauhinia tenuiflora)的木质部导管具有独特的“二元性” ,即同时具有粗大的导管(>400 μm)和众多细小导管(<30 μm) ,以适应不同水分条件下,植物水分传输、利用方式[51];三是植物叶片的生态性状的调控,岩溶植物的叶片、尤其是干旱条件更为严重的山顶植物,以植物叶片变小、角质层增厚(甚至发育蜡质层)、气孔指数降低,发达的表皮毛、栅栏组织等来减少水分丧失[52]。
广西马山弄拉鸡蛋堡的山顶和山腰上均生长青冈栎,两者的高度相差仅为几十米,但是其辐射、气温、水分或湿度等条件截然不同。山顶条件恶劣,因此,山顶植物叶片宽度、长度仅为山腰的57.30%、54.49%,叶片的厚度、角质层增厚增加39.70%、84.67%,气孔指数降低22.22%[53](表3)。

表3 不同地貌条件下青冈栎的抗旱性特征比较[53]Table 3 Comparison of drought resistance characteristics of Cyclobalanopsis glauca under different geomorphic conditions
广西弄岗岩溶峰丛洼地森林区,年均气温为22 ℃,年降水量为1 150~1 550 mm,选择同一植物品种苹婆(Sterculia monosperma),对比不同生境植物群落水分利用效率,分布于洼地的植物水分利用效率(WUE)为 28.12±16.30 μmol·mol-1,山腰的为44.26±7.04 μmol·mol-1、山顶的为46.52±8.60 μmol·mol-1,从洼地到山顶,苹婆水分利用效率显著提高。4 种山顶优势植物的水分利用效率比洼地的高65%[54](表4)。

表4 广西弄岗峰丛洼地不同生境植物群落水分利用效率[54]Table 4 Water use efficiency of plant communities in different habitats in Nonggang peak cluster depression, Guangxi
通过植物分泌生物酶、激素等,调控植物生理过程,提高适应岩溶区干旱能力。例如,对亚热带岩溶区常见植物青冈(Cyclobalanopsis glaucea)、圆叶乌桕(Sapium rotundifoliumHemsl. )、火棘(Pyracantha fortuneana(Maxim.) Li)和小果蔷薇(Rosa cymosa)的研究发现,随着干旱加剧,这些植物通过积累脯氨酸、可溶性糖等有机渗透调节剂调控质膜渗透势,维护细胞膜稳定,减少水分胁迫对细胞的伤害,同时累积钠、钾等无机离子调节液泡渗透势,维持细胞膨压[55]。岩溶区不同演替阶段代表性植物蚬木(Excentrodendron hsienmu(Chun et How) H. T. Chang et R. H.Miau)、构树(Broussonetia papyrifeera)和火棘等在适度干旱条件下依靠增加体内超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的含量,以清除活性氧自由基降低膜脂过氧化程度,避免干旱对其造成伤害,达到缓解植物干旱状况的目的[56-57],提高岩溶植物叶片碳酸酐酶活性,将细胞内碳酸氢根离子转化成水和CO2,补充干旱引起的叶片水分不足和胞间CO2减少,在提高植物水力安全的同时维持光合产物的合成,提高岩溶植物的干旱适应性[58]。
4 岩溶生态系统中植物的多样性
岩溶环境的生境异质性在不同的空间尺度上均存在,不同岩溶景观类型、不同地貌部位、不同坡向、小生境水分和养分的差异性等,生境的异质性可影响植物多样性的形成,甚至成为植物多样性维持的主导因子[59-60]。
岩溶生态系统中的植物多样性常表现为少属科、寡种属。云南西双版纳热带岩溶区面积约为3 600 km2,占土地面积的19%,海拔为600~1 600 m;岩溶区森林调查结果显示:拥有维管束植物153 科、640 属、1 394 种,分别占区系总科数的77.7%、属数的56.1%和种数的37.9%[61]。
广西弄岗北热带岩溶森林区面积100 km2的深入调查结果显示,维管束植被有184 科、810 属、1 752种[62-63],分别占广西植物区系的80.7%、72.3%、58.7%[6];广西木论中亚热带岩溶森林区植物区系维管束植被有175 科、528 属、906 种,分别占广西植物区系的76.8%、47.1%、30.4%[64];广西岩溶区面积约8.2 万km2,占土地面积的34.8%,早期调查揭示广西岩溶植被的区系由175 科、662 属、1 500 种组成[65],占 广 西 植 物 区 系(228 科、1 120 属、2 983 种)的76.8%、59.1%、50.3%[6]。
荔波县茂兰中亚热带岩溶森林区面积200 km2,维管束植物有154 科、514 属、1 203 种[20],占贵州省植物区系的61.6%、33.1%、21.1%[66];贵州岩溶区面积约11.6 万km2,占土地面积的61.2%,已有调查结果显示贵州共有维管束植物250 科、1 551 属以及5 691 种[67]。
贵州、广西、云南三省(区)岩溶地区的维管束植物共有195 科、1 213 属、4 287 种,其中包括许多特有属和特有种[61,68-69],植物种属比为3.8,显著低于全国8.5 的水平,反映出岩溶生态系统中植物种属的多样性。
岩溶生态系统中植物多样性的另一个重要特征是物种的特有性。如果将岩溶区生长发育的植物称为岩溶植物,则岩溶植物的组成可分为三种:特有种或专有种,即只能在岩溶区生存,在非岩溶区则无法生存;适宜或适应岩溶环境的适宜种,植物在岩溶区生长很好,在非岩溶区生长不好、但能生存;随遇种,即植物品种具有很强的适应能力,在岩溶区和非岩溶区均能良好生长。
根据植物学家的调查分类,西双版纳岩溶区已有调查的植被品种1 394 种,特有种141 种、适宜种170 种[70],分别占区系植物种数的10.1%和12.2%;广西弄岗北热带岩溶森林区已有调查的植被品种1 752种,特有种278 种、适宜种350 种[63],分别占区系植物种数的15.9%、20.0%;换言之,具有岩溶特色的植物品种占区系品种30%以上,如果岩溶生境遭遇破坏,则30%的具有特色的植物品种首先丧失,且恢复难度十分困难。例如,贵州荔波县茂兰岩溶森林保护区,被列入IUCN 物种名录和中国物种红色名录的植物种类涉及27 科、67 属、121 种,在121 种中,濒危植物有83 种,占总数的68.6%,包括贵州山核桃(Carya Kweichowensis) 、 狭 叶 含 笑(Michelia angustioblonga)、荔波瘤果茶(Camellia rubinuricata)、白花兜兰(Paphiopedium emersonii)等[71]。
5 岩溶生态系统中两类特有植物群落
5.1 岩溶洞穴弱光带植物
岩溶洞穴植物,是指岩溶洞穴生境中自然分布的植物。根据光照强度和外部气候影响程度,洞穴可被分为3 个区域:洞口带(cave entrance zone)、弱光带(cave twilight zone)和黑暗带(cave dark zone)[72]。由于黑暗带的光照强度几乎为0,仅能生长极少的、缺乏叶绿素的植物和一些真菌、细菌等,因此岩溶洞穴植物大多分布于洞口带和弱光带。
据不完全统计,中国从2001-2021 年间洞穴植物新种发表超过了80 种,其中描述最多新物种的前4人(第一定名人)分别是张丽兵(23 种蕨类植物)、温放(20 种种子植物)、王文采(17 种种子植物)、刘演(11 种种子植物)。
调查中国西南地区的60 多个岩溶洞穴,发现83科186 属418 种维管束植物,其中31 种(7%)为洞穴特有种、37%为中国特有种。88%的种类为草本、8%为灌木、4%为藤本,其中物种最丰富的科包括:荨麻科(73 种)、苦苣苔科(37 种)、秋海棠科(22 种)、凤尾蕨科(20 种)、鳞毛蕨科(20 种);最丰富的属包括:楼梯草属(42 种)、秋海棠属(22 种)、耳蕨属(19 种)、报春苣苔属(19 种)、冷水花属(13 种)[73]。
广西植物研究所植物多样性研究团队研究了岩溶洞穴代表类群报春苣苔属植物对低光环境的分子适应机制,发现与光合作用相关的光敏色素家族PHYE基因的两个位点受到正向选择,因而推测该功能基因在其适应洞穴环境的过程中可能起到关键作用,岩溶洞穴为植物物种的形成和适应性进化提供有利条件的场所[74]。洞穴植物叶片元素含量对洞穴环境是趋同适应[75]。
5.2 岩溶天坑植物群
天坑是岩溶地下河大厅顶板崩塌,由天窗逐渐演变而来,天坑的深度通常大于100 m,天坑环境相对封闭,底部与地下河相连,能更好地调节天坑内的温度、湿度,维持小气候,使其较少受外界气候变化干扰,为植物生存、发育提供良好、稳定的环境,能保育更多过去遗留下来的植物群落成分。经调查统计,广西乐业天坑植物群落种子植物有137 科、445 属、863 种,温带科和热带科之比为1∶2.31,温带属和热带属之比为1∶1.5;与坑外植被群落(177科945 属2 406 种;温带科和热带科之比为1∶2.79;温带属和热带属之比为1∶2.18)相比,多样性在科、属、种水平上分别降低22.59%、52.91%、64.13%,但坑内植物群落中温带成分比例更高,暗示着随着全球气候变暖,地表植被群落中热带组分不断增加,而坑内植物群落热带组分增加滞后。这为研究区域植物群落演替提供了更多样本[76-77]。
天坑植物区系的演化相比天坑地质发育滞后:天坑发育前期,地下大厅开始崩塌,坑口面积小于坑底面积,天坑形状呈倒置漏斗状,透入的光照少、空气潮湿,光照强度由坑口向坑内逐渐递减。天坑内植被区系起源经历苔藓植物阶段、藤类植物阶段和耐阴草本-灌木阶段;天坑发育中期,天坑口与天坑底面积相近,此时天坑保温、保湿效果较好,从天坑口透入的光照增多、空气潮湿、土壤加厚。此时天坑植物区系具有强烈的热带性,热带性质的属的比例高达70%,比地表植物区系热带属的比例高10%,相反,温带属的比例比天坑外温带属的比例小7%;天坑发育后期,天坑退化,悬崖不断崩落,坑底不断被垫,形状呈漏斗状,坑口部面积大于底部面积,天坑所产生的湿热小环境特征变得不明显,环境变得和地表趋于一致。此时,植被群落热带属的比例为65%,但是仍然比地表植物区系热带属的比例要高5个百分点。而温带属的比例升高,达到25%,逐渐接近外界植物区系中温带属比例,但仍要小3 个百分点[76, 78-79]。
6 展 望
(1)学科交叉,完善岩溶生态系统理论 影响生态系统形成、演变的因素中,气候因素、水文因素是塑造生态系统长期的、缓慢的驱动力,而地质、地貌因素则是生态系统得以存在和发展的载体和物质基础。岩溶生态系统明显受地质条件的制约,植物作为生态系统的初级生产者,其生产力关乎整个生态系统的物质和能量供给。植物生存发育的前提条件是具有足够的水分和养分,而岩溶区是缺土少水、富钙偏碱的岩溶环境,深入了解植物适应岩溶环境的内在机制,从元素生物地球化学循环的角度揭示岩石-土壤-植物间水分、养分、元素迁移和富集规律,不仅要将焦点关注岩溶表层带岩石-土壤-根系融合体系对植物生产者的支撑作用,更要加深揭示植物-岩石间相互作用所带来的生理、生态特性,这就需要生态工作者与地学工作者密切合作。多学科交叉渗透,尤其是充分发挥中国岩溶研究的地域优势,尽快获得中国不同纬度、不同海拔植物对岩溶环境的适应机制和其生理生态特性,以充分利用岩溶生态系统结构特征,认识同一地区植物群落演替过程和内在规律,尤其是当前全球气候变暖、极端天气频发的形势下,完善中国岩溶生态系统理论。
(2)退化岩溶生态系统修复技术亟需研发 岩溶生态系统是脆弱的生态系统,在人为干扰下,在20世纪末至21 世纪初,中国西南岩溶区出现大面积的石漠化现象,土地生产力严重下降,“一方水土养活不了一方人”。为实践“绿水青山就是金山银山”的生态文明理念,退化岩溶生态系统(极端表现形式是重度石漠化)面临两大科学问题:一是在地质-气候不同配置下,揭示其植物生态修复瓶颈技术的科学基础,以及仿自然植被群落快速恢复人工培育技术的研发。二是筛选适应性、适宜性经济植物,尤其富钙经济植物的培育,发展富钙食品经济,揭示其对地质-气候不同配置下的生长、发育和演替更新过程和机制;研发消除障碍、特色生态肥料、延长其生态经济“生命周期”的系列技术;研发“互利共赢”的套种、间种技术,探索可持续发展途径。
中国西南岩溶区保存有贵州茂兰、广西木论和弄岗、云南西双版纳等原始的岩溶森林保护区,其为探索脆弱岩溶生态系统中发展具仿自然特色的生态产业,改善当地居民生产生活条件,实践“绿水青山就是金山银山”道路,提供了植物“种源库”和“基因库”,鉴于岩溶生境强烈的空间异质性,水土资源空间配置的差异性,还有大量的科学问题、技术问题亟需解决,更多岩溶区生态经济协调协同、且可持续途径亟需探索。
致谢:本文撰写时间比较仓促,撰写过程中得到多位志同道合的专家的支持,尤其感谢广西植物研究所李先琨、韦毅钢等老师、南宁师范大学黄玉清老师提供了相关的素材和数据。
