湖南高望界国家级自然保护区雪峰山梭罗种群结构与动态特征
2022-11-29王春晖王本忠徐展宏
王春晖,陈 昕*,王本忠,徐展宏
(1.南京林业大学,南方现代林业协同创新中心,江苏 南京 210037;2.南京林业大学生物与环境学院,江苏 南京 210037; 3.湖南高望界国家级自然保护区管理局,湖南 古丈 416300;4.南京林业大学林学院,江苏 南京 210037)
种群是物种存在的基本单位,种群结构影响群落结构[1-2]。基于绘制生命表及生存曲线等科学研究方法,可揭示种群与环境之间的相融度和发展趋势[3]。通过分析种群的年龄结构、静态生命表和生存状况,发现不同年龄阶段个体在群落中的生存特点,揭示种群历史动态特征与受干扰状况,也能预测未来动态及演变趋势[4-6]。因此,研究种群结构与数量动态特征可以揭示种群的生存状况,对种群资源的保护有重要意义。
雪峰山梭罗(Reevesiapubescensvar.xuefengensis)属锦葵科(Malvaceae)梭罗树属(Reevesia),是中国特有树种,1981年在湖南雪峰山第一次发现,数量较少,零星分布在湖南省西南部一带[7]; 2018年王本忠等[8]在湖南高望界保护区内发现雪峰山梭罗群落。为了解雪峰山梭罗种群的生态特征,通过对样地内雪峰山梭罗种群进行结构划分、数量动态量化、生存分析、时间序列分析等研究,深入探究高望界保护区雪峰山梭罗种群的基本特征,以及该种群的发展趋势,为雪峰山梭罗的有效保护提供科学参考。
1 材料与方法
1.1 研究区概况
高望界国家级自然保护区位于湖南省湘西土家族苗族自治州古丈县境内(109°58′28″~110°14′38″E,28°36′32″~28°45′39″N),总面积17 169.8 hm2,海拔190.0~1 146.2 m[9]。该区域降雨量大、光照充足、小气候显著,为亚热带山地季风气候[10]。研究区内的乔木层除雪峰山梭罗,主要还有青冈(Cyclobalanopsisglauca)、锥栗(Castaneahenryi)、马尾松 (Pinusmassoniana)等;灌木层有马银花(Rhododendronovatum)、紫果槭(Acercordatum)、水丝梨(Sycopsissinensis)等;草本层有蛇根草(Ophiorrhizamungos)、两色鳞毛蕨(Dryopterissetosa)等。
1.2 研究方法
1.2.1 样地设置与野外调查
2020年11月,在高望界自然保护区内实地踏查,采用典型样地法,选取集中覆盖雪峰山梭罗种群的地段,设置1块250 m × 350 m的样地,位于山脊线附近,半阳坡,海拔755 m,坡度39°,逐木调查所有目标树种,详细记录其株高、胸径、冠幅、枝下高等指标。通过GPS定位,记录目标树种经纬度、海拔等地形因子。
1.2.2 种群结构划分
从雪峰山梭罗种群的野外实地调查数据中发现雪峰山梭罗个体数量较少,分布零星,真实树龄较难测定,故采用“空间代替时间”的研究方法,以结构级代替年龄级[11-13]。根据雪峰山梭罗的生活史特点将其胸径(DBH)、基径(BD)划分为10级,高度(H)为9级,冠幅(C)为8级(表1)。
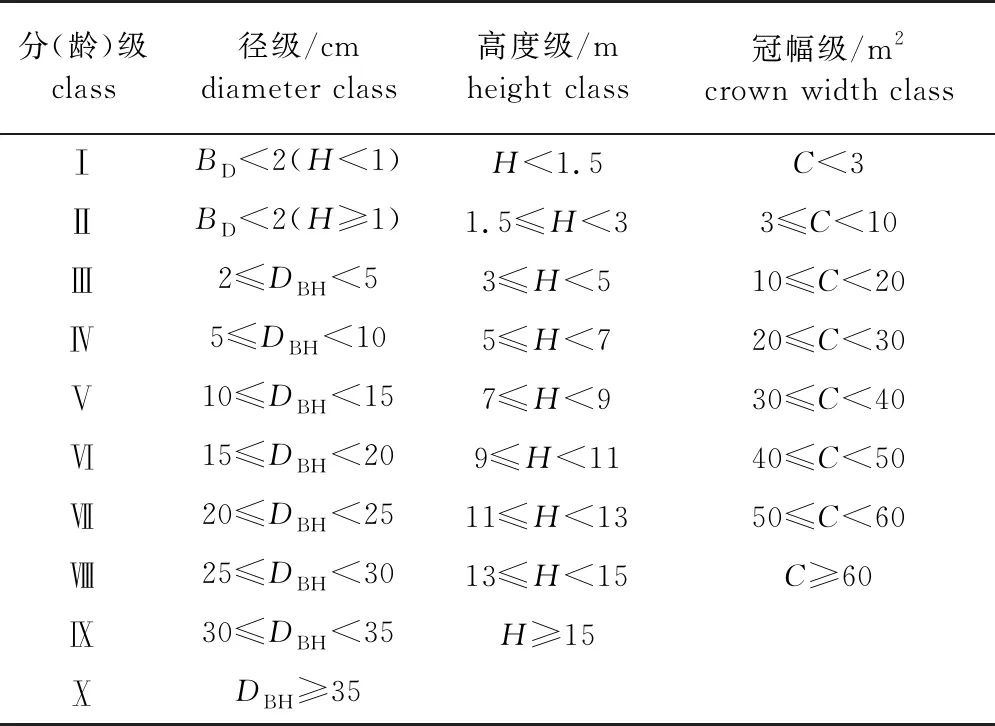
表1 雪峰山梭罗种群结构级划分
1.2.3 种群数量动态量化
采用陈晓德[14]的动态量化分析方法研究雪峰山梭罗种群动态变化,方法如下:

种群对外界干扰所承担的风险概率Pmax=1/[kmin(S1,S2,…,Sk)]。式中,P取值为最大时才会对种群动态V′pi构成最大影响[16]。
1.2.4 静态生命表编制和生存分析
静态生命表描述出种群的死亡过程与环境之间的关系,计算公式及各参数如下:
x龄级开始时标准化存活个体数lx=ax/a0×1 000。式中:a0为固定值,第Ⅰ龄级现有个体数;ax指x龄级内现有个体数。
x至x+1龄级内的标准化死亡数dx=lx-lx+1。
x至x+1龄级内的死亡率qx=dx/lx。
x至x+1龄级内的平均存活的个体数Lx=(lx+lx+1)/2。
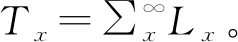
进入x龄级个体的平均期望寿命ex=Tx/lx。
各龄级内的消失率Kx=lnlx- lnlx+1。
引入存活曲线来描述特定年龄的死亡率,以龄级为横坐标,以标准化存活个体数的自然对数值(lnlx)为纵坐标绘制雪峰山梭罗的存活曲线,可反映雪峰山梭罗种群动态特征及各年龄阶段的存活状况[17]。Deevey[18]将存活曲线划分为3种基础类型:Ⅰ型为凸曲线,即种群个体几乎能达到该物种的寿命,在种群幼期状态时死亡率比较小;Ⅱ型为对角线,即种群各个年龄期的死亡率是基本相等的;Ⅲ型为凹曲线,即幼体死亡率高,到达某个年龄阶段后死亡率低而稳定[19-20]。参考Hett等[21]提出的两种数学模型来检验雪峰山梭罗种群的存活状况属于Deevey-Ⅱ型或Deevey-Ⅲ型存活曲线。
因为静态生命表编制的前提是假设种群所经历的环境条件不发生变化,所以在实际中具有误差和局限性,不能准确显示由于环境影响而产生的具体变化[22-23]。为了能得到在某时刻种群的动态结构特征,利用生存函数进行分析[24-25],参考杨凤翔等[26]的公式,引入以下4个函数来分析其种群的动态特征:
生存率(S(i))=S1×S2×…×Si;
累积死亡率(F(i))=1 -S(i);

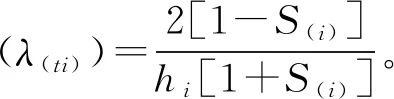
以龄级为横坐标,4个函数值为纵坐标绘制雪峰山梭罗种群的生存率曲线、累计死亡率曲线、死亡密度曲线和危险率曲线。
1.2.5 时间序列分析
运用一次移动平均法进行时间序列分析[27]对雪峰山梭罗种群年龄结构进行预测,其目的是对未来经过2、4、6、8个龄级后的雪峰山梭罗种群各龄级的个体数量进行预测。具体公式为:
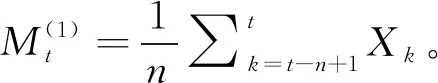
2 结果与分析
2.1 雪峰山梭罗种群结构特征
2.1.1 径级结构
雪峰山梭罗种群径级结构分布如图1所示,在样地内种群的个体数共有182株。第Ⅰ龄级是幼苗,占总数28.57%,林分密度为5.94株/hm2;第Ⅱ龄级是幼树,占总数的25.27%,林分密度为5.26株/hm2;第Ⅲ龄级是小树,占总数15.38%,林分密度为3.2株/hm2。从总体看第Ⅰ—Ⅲ径级个体数量较多,其中从幼树到小树的过程中,数量减少较快,个体死亡增多;第Ⅳ—Ⅴ龄级是中树,占总数18.14%,林分密度为3.77株/hm2,由此可知中树各龄级的数量平稳下降;第Ⅵ—Ⅷ龄级是大树,大树的个体数量仅19株,较少,林分密度为2.17株/hm2;最后2个龄级属于老树,该阶段仅4株,单株胸径最大达到49.30 cm,胸径超过40 cm的雪峰山梭罗有2株。

图1 雪峰山梭罗种群径级结构
2.1.2 高度结构
雪峰山梭罗种群个体数随高度级分布如图2所示,可知第Ⅰ高度级的个体数最多,占总数36.26%,林分密度是7.54株/hm2;第Ⅱ高度级的个体数迅速下降到35株,占总数19.23%,林分密度是4.00株/hm2;第Ⅸ高度级的个体数最少,占总数2.20%,林分密度为0.46株/hm2,单株高度最高可达16 m,高度超过15 m的雪峰山梭罗有4株。种群高度级结构较完整,9个高度级均有个体分布。
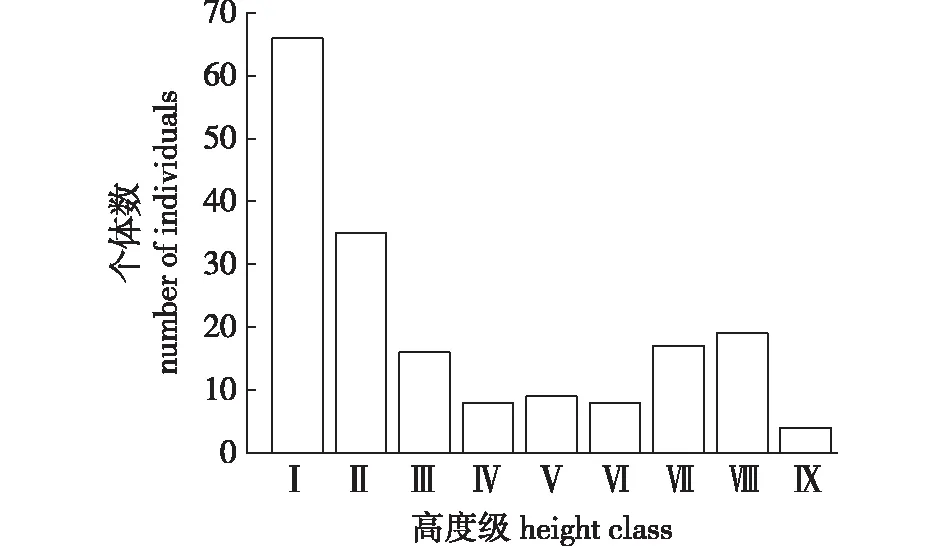
图2 雪峰山梭罗高度级结构
2.1.3 冠幅结构
幼苗、幼树的个体较小,不适合进行冠幅计算统计,故除去98株幼苗、幼树,参与统计的雪峰山梭罗个体数为84株,冠幅的测量指标以树冠投影圆的面积大小统计,冠幅级结构如图3所示。第Ⅰ冠幅级的数量为5株,占总数的5.95%,林分密度为0.57株/hm2;第Ⅱ级的个体数上升到最大,个体数为28株,占总数的33.33%,林分密度为3.20株/hm2;第Ⅲ—Ⅵ级个体数不断下降,第Ⅵ级的个体数仅2株,占总数的2.38%;第Ⅶ级的个体数上升到10株,第Ⅷ级的个体数为7株,单株冠幅最大可达95.03 m2,冠幅超过70 m2的雪峰山梭罗有7株,占总数的8.33%。
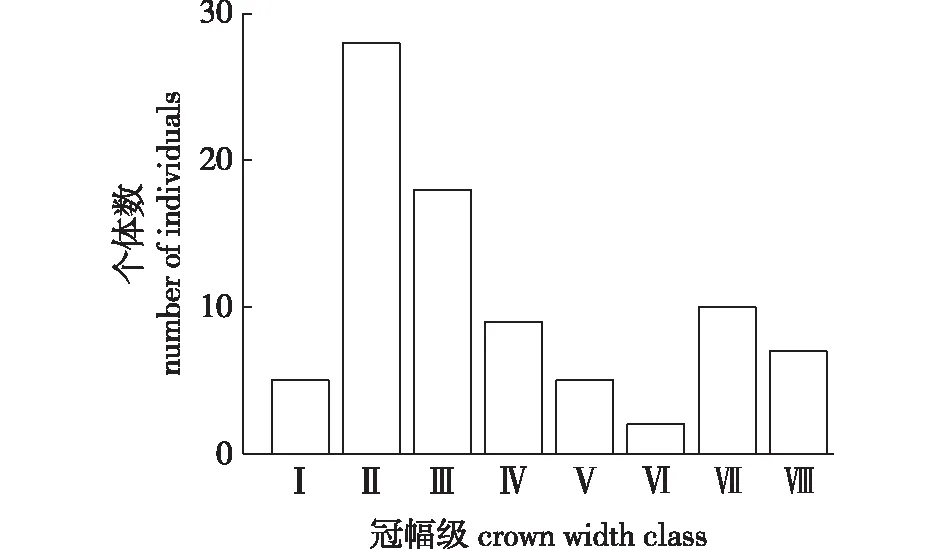
图3 雪峰山梭罗冠幅级结构
2.2 雪峰山梭罗种群数量动态
在种群数量动态量化分析过程中,使用径级结构数据将会出现风险概率Pmax公式中分母为零的情况,无法进行计算,故利用高度级结构能更合理地对雪峰山梭罗种群数量进行动态量化分析。经分析(表2)可知,雪峰山梭罗种群动态指数V4、V6、V7均小于0,表明在第Ⅳ、Ⅵ、Ⅶ龄级出现衰退的结构的动态关系,第Ⅲ龄级和第Ⅴ龄级分别向第Ⅳ龄级和第Ⅵ、Ⅶ龄级转化时种群增长受到影响[28];V1—V3、V5和V8均大于0,表示第Ⅰ—Ⅲ、Ⅴ和Ⅷ龄级的种群结构动态关系出现增长趋势;不考虑外部干扰时Vpi为37.69%,考虑外部干扰时V′pi为0.52%趋近于0,种群对完全随机干扰所承担的最大风险概率(Pmax)为1.39%。
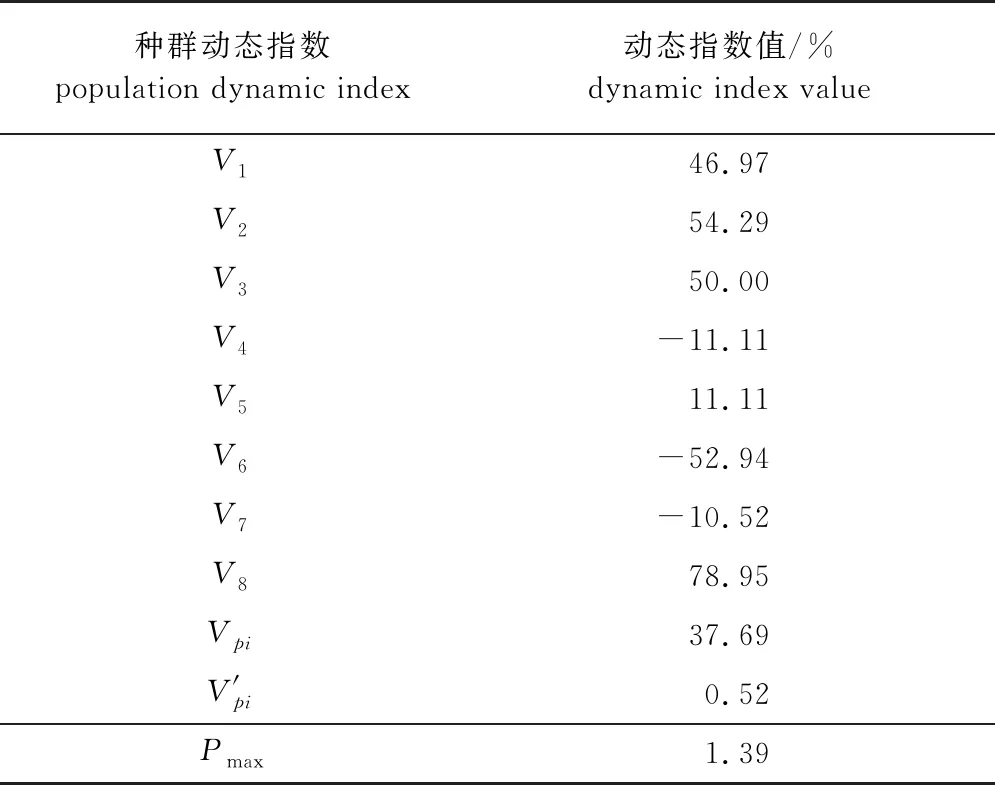
表2 雪峰山梭罗种群动态变化指数
2.3 雪峰山梭罗种群静态生命表及存活曲线
经分析可知,第Ⅵ龄级和第Ⅸ龄级相比第Ⅶ龄级和第Ⅹ龄级的存活数减少,依据特定时间生命表的假设前提,龄级变化是稳定的。由此,对第Ⅵ—Ⅹ龄级使用匀滑处理,经修正后得ax,Ⅵ—Ⅹ龄级的存活数分别为9、7、5、3、1。
由此得出雪峰山梭罗种群静态生命表(表3),经分析可知雪峰山梭罗种群存活数(lx)随龄级增加而减少。个体期望寿命(ex)随龄级的增加呈现先减后增再减,具有一定的波动性。期望寿命(ex)可以表现出各龄级内个体的平均生存能力[29],在Ⅰ、Ⅱ龄级中,最小期望寿命在第Ⅸ龄级处(ex=0.667),最大期望寿命在第Ⅰ龄级处(ex=3.202),整体趋势是逐渐降低。
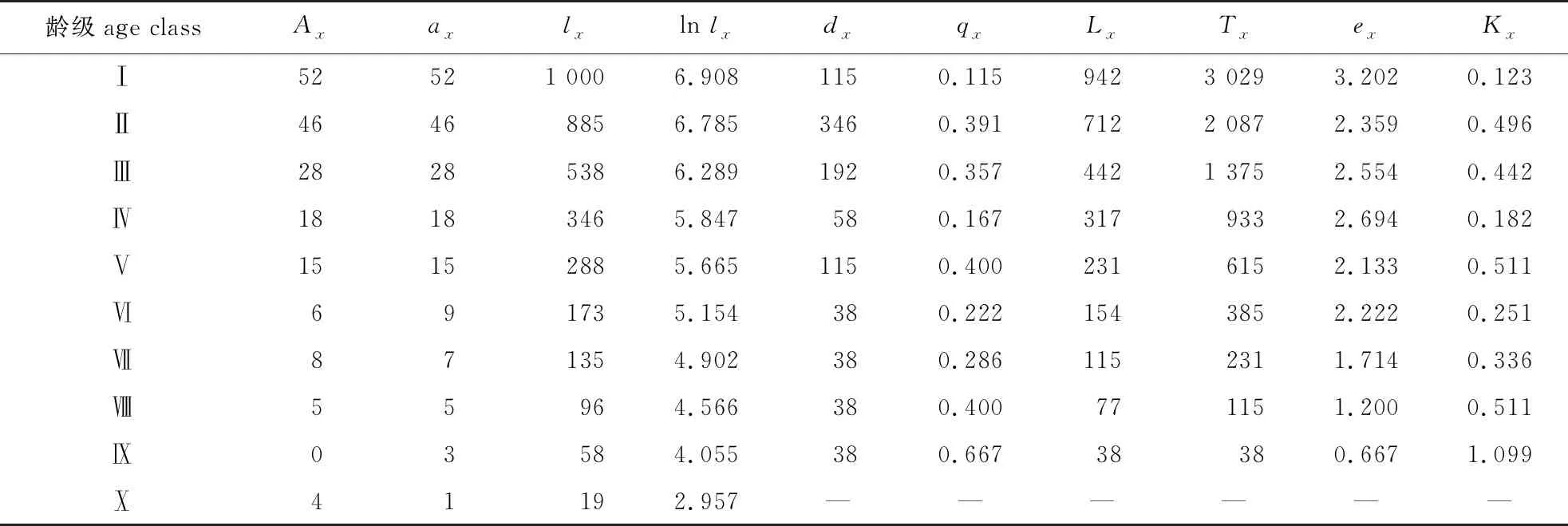
表3 雪峰山梭罗种群静态生命表
雪峰山梭罗种群存活曲线见图4。根据Deevey理论可以判定,雪峰山梭罗种群的存活曲线可能是Deevey-Ⅱ型或Deevey-Ⅲ型。应用Hett等数学模型对雪峰山梭罗种群存活曲线类型进行检验,建立其相应模型:Nx,Ⅱ=7.795 37e-0.073 59x(F=1 265.231 14,R2=0.924 76);Nx,Ⅲ=7.608 56x-0.247 08(F=340.220 67,R2=0.722 56)。指数模型的F值和R2值均比幂函数模型的大,故雪峰山梭罗种群存活曲线更趋于Deevey-Ⅱ型。
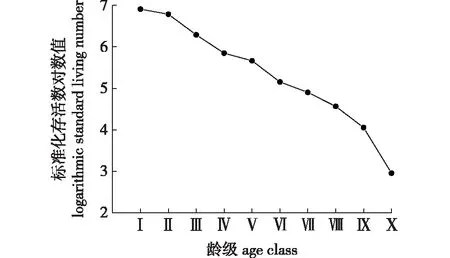
图4 雪峰山梭罗种群存活曲线
雪峰山梭罗种群的死亡率(qx)和消失率(Kx)曲线见图5。
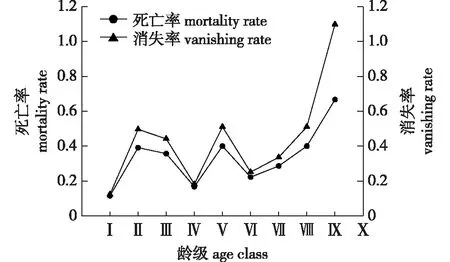
图5 雪峰山梭罗种群死亡率和消失率曲线
由图5可知,qx与Kx的变化趋势基本相同,变化幅度有所差别。在第Ⅱ龄级时出现qx和Kx的第一个峰值,分别为0.391和0.496,由于幼苗阶段个体对养分需求增大,种间竞争激烈,导致此阶段个体大量死亡;此后,qx和Kx波动下降,在Ⅵ龄级达到低点,分别为0.222和0.251,说明雪峰山梭罗种群经历了环境筛选和竞争后,适应能力增强;随后qx和Kx缓慢上升,在Ⅸ龄级时达到最大值,分别为0.667和1.099,说明此时基本上达到了生理寿命。
2.4 雪峰山梭罗种群生存分析
雪峰山梭罗种群生存曲线见图6,可知在第Ⅴ龄级以后,变化趋势变平缓。分析发现,雪峰山梭罗种群随龄级增高累计死亡率(F(i))单调上升,生存率(S(i))单调下降。第Ⅰ—Ⅲ龄级生存率明显下降累计死亡率大幅上升,生存率与累计死亡率在第Ⅱ龄级与第Ⅲ龄级间达到平衡状态。表明种群在低龄阶段向下一阶段生长过程中死亡数量多;死亡密度(f(ti))最大值出现在第Ⅱ龄级,危险率(λ(ti))最大值出现在第Ⅹ龄级。整体来看,f(ti)和λ(ti)曲线变化趋势均为先上升后下降再上升(图6)。
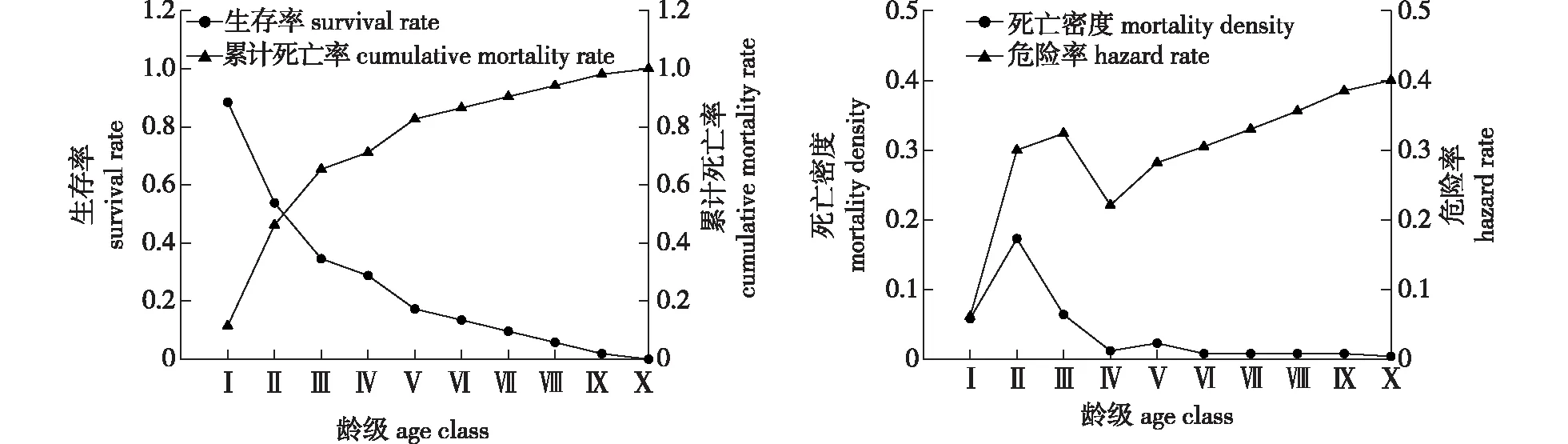
图6 雪峰山梭罗种群生存率和累计死亡率、死亡密度和危险率曲线
2.5 雪峰山梭罗种群时间序列预测
种群的时间序列预测结果见表4。通过预测结果可知,除了经过2个龄级时间后的第Ⅶ龄级外,其余龄级雪峰山梭罗种群个体数均在增加。第Ⅱ、Ⅳ、Ⅵ、Ⅷ龄级在未来2、4、6、8个龄级时间后,增加比例分别为6.52%、56.52%、64.71%、69.23%。
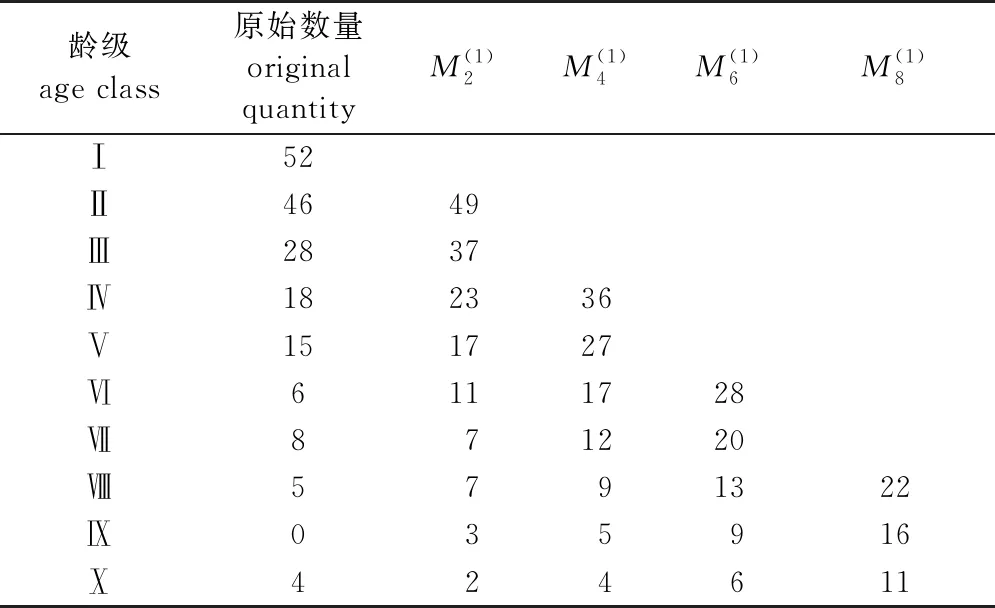
表4 雪峰山梭罗种群动态变化的时间序列预测
3 讨 论
生存环境之间存在相互作用会对种群的结构与动态产生影响。径级结构作为种群龄级划分的重要参数可以反映种群的空间分布[30],高度结构反映种群的垂直空间结构及个体间的资源竞争力差异[31],冠幅结构反映个体对生存空间的使用能力[32]。种群结构分析表明,雪峰山梭罗种群低龄级个体占优势,随着龄级增加优势逐级弱化,虽然幼苗幼树的数量多,但生存率较低,原因是幼苗幼树对于环境要求较高,适应能力较弱,低龄级向中龄级成长过程中个体大量死亡,而后保持相对稳定,种群总体呈增长趋势。除了受自然或人为因素干扰,高龄级植物自身生理代谢能力减弱、对营养需求较高,导致高龄级个体数量稀少[33]。
从种群结构数量动态变化指数可以看出,雪峰山梭罗种群在遭受外界环境干扰情况下,对外界干扰较为敏感,抗干扰能力弱,且种群稳定性差,这些特点对种群个体数量增长有一定影响。在实地调查中也发现,雪峰山梭罗幼苗常因多株较为密集的生长在高大乔木周围,导致林内光照条件较差,不能充足得到土壤养分,同层植物间的生存竞争变得激烈,所以在向下一阶段龄级生长时自然更新能力不足。由静态生命表可看出,雪峰山梭罗种群有3次死亡高峰。第1次死亡高峰出现在第Ⅱ龄级,此阶段大量幼树向小树成长,对外界不利环境的抵抗力较弱,经过激烈的环境筛选和种内竞争,死亡率增高;第2次死亡高峰出现在第Ⅴ龄级,此阶段的个体生存能力和竞争力增强,但受密度制约效应的影响[34],有限的生存空间不能满足此阶段所有个体的生长需求,结果导致种内竞争再次增强,死亡率也出现增高;第Ⅸ龄级出现第3次死亡高峰,致死原因是老树生命活动减弱对环境的适应能力下降,表示该阶段接近物种生理寿命,逐渐死亡。雪峰山梭罗种群符合Deevey-Ⅱ型种群存活曲线,表明该种群有相对稳定的增长趋势。结合雪峰山梭罗死亡率和消失率曲线分析可得,如不能提高低龄级个体存活率,随时间推移,中龄级个体数得不到补充,则种群将有可能进入衰退的发展趋势。生存率函数和累计死亡率函数曲线变化趋势相反,整体变化特征可总结为前期大幅减少、中期较为稳定、后期逐渐衰退,这也与种群径级结构、静态生命表和存活曲线的分析结果相统一。通过分析时间序列预测结果可知,整体来看雪峰山梭罗个体数是增加的,幼龄个体数较多,可以良好的补充中后期种群,同时老龄树逐渐增多,雪峰山梭罗种群在未来会有增长趋势。这也与种群径级结构、静态生命表及存活曲线的分析结果一致。
本研究区内乔木密度较高,林分郁闭度较高,林内光照不足,对幼苗、幼树向小树过渡产生影响。从长远分析,该种群对环境敏感度高,种群稳定性差,可能会制约该种群的自然更新和发展。鉴于雪峰山梭罗种群幼龄死亡率高的特点,可采用适度人工辅助,适当去除林下同层植物,为幼苗生长创造条件,促进幼苗幼树向小树过渡,后将分布较密集的小树移栽到适宜该种群生长的林内较空旷处,同时加强对该树种的保护,禁止乱砍乱伐和破坏自然生境的人为行为。基于雪峰山梭罗生物学特性,探究种群更新机制,可促进种群的稳定增长。
