lncRNA GAS5调控Notch通路对炎症环境下牙髓干细胞成骨分化的影响
2022-11-28何龙云蔓吴薇薇翁海勇王琼超
何龙 云蔓 吴薇薇 翁海勇 王琼超
牙髓干细胞(dental pulp stem cells,DPSCs)具有与骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMSCs)相似的免疫表型和多能分化特征[1]。据报道,炎性DPSCs的成骨能力低于正常DPSCs[2],但对其机制的了解仍然有限。明确炎症环境下DPSCs的成骨分化受到抑制的分子机制,对寻找有效的牙齿再生的药物至关重要。长链非编码RNA生长阻滞特异性转录本5(long non-coding RNA growth arrest-specific transcript 5,lncRNA GAS5)在多种癌症中受到抑制[3]。研究发现lncRNA GAS5的过表达通过调节microRNA-498上调RUNX2的表达来促进hMSCs的成骨分化[4];可促进人牙周膜干细胞(human periodontal ligament stem cells,hPDLSCs)的成骨分化。在成人牙齿龋坏或受伤的牙髓区域的间充质细胞中,Notch信号被上调[5]。Notch通路的激活可以抑制DPSCs的成牙本质细胞分化,并抑制间充质祖细胞的成骨细胞分化[6]。而lncRNA GAS5是与Notch信号相互作用的lncRNA之一[7],GAS5的敲低与Notch通路的激活和细胞炎性损伤有关[8]。推测lncRNA GAS5可能调控Notch通路影响人牙髓干细胞(hDPSCs)成骨分化。本研究采用脂多糖(lipopolysaccharide,LPS)溶液和过表达lncRNA GAS5的慢病毒载体同时处理hDPSCs,探讨其对成骨分化的影响,并初步探讨其机制。
1 材料与方法
1.1 实验材料
hDPSCs(BFN60810649,上海青旗生物技术发展有限公司);胎牛血清、DMEM培养基、胰蛋白酶(Gbico公司,美国);LPS(L5293)(Sigma-Aldrich,美国);RIPA裂解液(P0013B)(上海碧云天生物科技公司);Trizol试剂(9108-1);过表达lncRNA GAS5的慢病毒载体及其对照空载体(上海Gene Pharma公司);Jagged-1(Notch通路的激动剂,HY-P1846A)(MedChemExpress公司,美国);骨钙素(osteocalcin,OCN)(ab93876)、Hey1(hairy and enhancer of split-related with YRPW motif 1)(ab154077)、发状分裂相关增强子1(hairy and enhancer of split 1,Hes1)(ab108937)、GAPDH(ab9485)、山羊抗兔IgG H&L(HRP)(ab205718)(Abcam公司,英国)。细胞培养箱、NanoDrop 2000分光光度计(Thermo Fisher Scientific公司,美国);ABI Prism®7300型荧光定量PCR系统(ABI公司,美国);IX73倒置荧光显微镜(Olympus公司,日本);iMark680多功能酶标仪、蛋白转膜装置、凝胶成像系统(Bio-Rad公司,美国)。
1.2 实验方法
1.2.1 细胞培养 hDPSCs在含有10%胎牛血清和1%青霉素-链霉素溶液的DMEM培养基中,于37 ℃、5%CO2的湿润培养箱中培养和传代。每3 d补充新鲜培养基,待细胞达到80%融合时进行传代。取第3代细胞用于后续实验。
1.2.2 细胞分组与处理 将hDPSCs(第3代)以每孔1×105个细胞的密度接种在6 孔板中,分为(1):正常对照组:细胞在含有10%胎牛血清的培养基中正常培养;(2)成骨诱导组:用成骨诱导培养基(含有10%胎牛血清、100 nmol/L地塞米松、10 mmol/L β-甘油磷酸、50 mg/mL抗坏血酸)培养hDPSCs;(3)LPS组:用含10 μg/mL LPS的成骨诱导培养基培养hDPSCs;(4)NC+LPS组:慢病毒对照空载体感染hDPSCs 72 h,然后用含10 μg/mL LPS的成骨诱导培养基培养hDPSCs;(5)GAS5过表达组:含有过表达lncRNA GAS5的慢病毒载体感染hDPSCs 72 h,然后用含10 μg/mL LPS的成骨诱导培养基培养hDPSCs;(6)Jagged-1组:用含10 μg/mL LPS和10 μmol/L Jagged-1的成骨诱导培养基培养hDPSCs;(7)GAS5过表达+Jagged-1组:在过表达lncRNA GAS5的慢病毒载体感染hDPSCs 72 h后,用含10 μg/mL LPS和10 μmol/L Jagged-1的成骨诱导培养基培养hDPSCs。每隔3 d更换新鲜培养基。倒置显微镜下观察hDPSCs形态变化。
1.2.3 CCK-8检测细胞增殖活性 将hDPSCs以每孔2×103个细胞接种在96 孔培养板中,在诱导分化的第1、3和7天用CCK-8检测细胞增殖活性。加入10 μL CCK-8试剂,37 ℃避光孵育细胞2 h。使用多功能酶标仪在490 nm波长处测量各孔的光密度(A)值,以表示细胞增殖活性。
1.2.4 RT-qPCR检测细胞中lncRNA GAS5的表达量变化 取诱导分化14 d的hDPSCs,使用Trizol试剂分离总RNA。NanaDrop分光光度计检测总RNA的浓度和纯度;RNA反转录为cDNA,使用SYBR®Premix Ex TaqTM试剂盒进行RT-qPCR。反应体系(25 μL):SYBR®Premix Ex TaqTM(2×)12 μL,上下游引物各0.5 μL,cDNA(200 ng/μL) 1 μL,ddH2O 11 μL。反应条件:94 ℃预变性10 min;95 ℃变性15 s,60 ℃退火20 s,75 ℃延伸20 s,进行40 个循环。用GAPDH作为内参对照,采用2-ΔΔCt方法计算目的基因的相对表达量。lncRNA GAS5正向引物:5′-GTGTGGCTCTGGATAGCAC-3′和反向引物:5′-ACCCAAGCAAGTCATCCATG-3′;GAPDH正向引物:5′-ACAGGGGAGGTGATAGCATT-3′和反向引物:5′-GACCAAAAGCCTTCATACATCTC-3′。
1.2.5 ELISA检测细胞上清液中炎症因子TNF-α、IL-1β水平 收集诱导分化14 d的hDPSCs细胞培养上清液,按照ELISA试剂盒的说明书操作检测上清液中TNF-α、IL-1β的水平。
1.2.6 比色法检测hDPSCs中ALP活性 在诱导分化第7天,弃去培养液,使用0.1% TritonX-100裂解细胞,12 000 g离心10 min,取上清液,使用ALP活性检测试剂盒测定上清液中ALP活性。
1.2.7 茜素红染色检测hDPSCs中钙离子沉积情况
hDPSCs培养14 d,用4%多聚甲醛固定后,加入200 μL茜素红S染液。显微镜下观察钙化结节情况,拍照后使用Image-Pro Plus 6.0软件统计钙化结节染色阳性区域的面积及钙化结节的数量。
1.2.8 Western blot检测hDPSCs中Notch通路和成骨相关蛋白的表达 BCA蛋白测定法测定蛋白质浓度。在室温下用5%脱脂奶粉(溶于TBST,TBS加0.1%Tween-20)封闭1 h后,在4 ℃下用一抗(NICD、Hes1、Hey1、RUNX2、OCN,1∶1 000;GAPDH,1∶5 000)孵育过夜。随后,将膜与HRP标记的山羊抗兔IgG(H+L,1∶10 000)在室温下孵育1 h。然后使用ECL显色,使用Image-Pro Plus 6.0软件量化条带灰度值并以GAPDH为内参对照进行标准化。
1.3 统计学分析
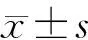
2 结 果
2.1 hDPSCs细胞形态
体外培养hDPSCs结果显示,原代细胞培养5~7 d,hDPSCs表现出典型的间充质细胞特点,可见细胞贴壁生长,细胞呈长梭形,成纤维细胞样生长(图 1A);原代细胞传代培养(正常对照组),细胞贴壁生长呈旋涡状(图 1B);用成骨诱导培养基诱导成骨分化后,hDPSCs仍具有成纤维细胞样的梭形形态,但是较正常对照组培养基中的细胞更大,在钙化沉淀区域有细胞簇形成(图 1C);经LPS处理后,细胞密度降低,稀疏,细胞间隙增大,细胞未见明显伸展(图 1D);NC+LPS组细胞形态和LPS组相似(图 1E);GAS5过表达组细胞排列紧密,多为长梭形,呈漩涡样生长,有细胞簇形成(图 1F);Jagged-1组细胞体积较小,细胞密集度不高,细胞多呈圆形,长梭形细胞较少(图 1G);GAS5过表达+Jagged-1组细胞体积有所增大,排列较为密集,长梭形细胞较Jagged-1组增多(图 1H)。
2.2 各组细胞增殖能力比较
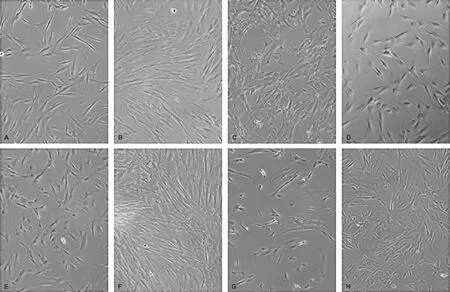
图 1 hDPSCs细胞形态(相差显微镜, ×400)
在诱导分化的第3和7天,与正常对照组相比,成骨诱导组细胞的增殖活性升高(P<0.05),而LPS组细胞的增殖活性降低(P<0.05);与LPS组相比,GAS5过表达组细胞的增殖活性升高(P<0.05),Jagged-1组细胞的增殖活性降低(P<0.05);与GAS5过表达组相比,GAS5过表达+Jagged-1组细胞增殖活性降低(P<0.05);与Jagged-1组相比,GAS5过表达+Jagged-1组细胞增殖活性升高(P<0.05)(表 1)。
表 1 各组细胞增殖能力比较
Tab 1 Comparison of cell proliferation among the groups

表 1 各组细胞增殖能力比较
组 别1 d3 d7 d正常对照组0.24±0.030.50±0.060.71±0.08成骨诱导组0.25±0.050.63±0.07①0.82±0.11①LPS组0.22±0.040.38±0.05①0.41±0.06①NC+LPS组0.21±0.030.36±0.040.43±0.07GAS5过表达组0.27±0.050.72±0.08②0.95±0.10②Jagged-1组0.20±0.040.25±0.04②0.30±0.05②GAS5过表达+Jagged-1组0.23±0.040.49±0.06③④0.64±0.08③④
注: 与正常对照组相比, ①P<0.05; 与LPS组相比, ②P<0.05; 与GAS5过表达组相比, ③P<0.05; 与Jagged-1组相比, ④P<0.05
2.3 各组细胞中lncRNA GAS5的表达量变化
与正常对照组相比,成骨诱导组细胞中lncRNA GAS5水平升高(P<0.05),而LPS组细胞中lncRNA GAS5水平降低(P<0.05);与LPS组相比,GAS5过表达组细胞中lncRNA GAS5水平升高(P<0.05);与Jagged-1组相比,GAS5过表达+Jagged-1组细胞中lncRNA GAS5水平升高(P<0.05)(表 2)。
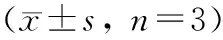
表 2 各组细胞中lncRNA GAS5的表达量变化
2.4 各组细胞上清液中TNF-α、IL-1β水平变化
与正常对照组相比,LPS组TNF-α、IL-1β水平升高(P<0.05);与LPS组相比,GAS5过表达组TNF-α、IL-1β水平降低(P<0.05),Jagged-1组TNF-α、IL-1β水平升高(P<0.05);与GAS5过表达组相比,GAS5过表达+Jagged-1组TNF-α、IL-1β水平升高(P<0.05);与Jagged-1组相比,GAS5过表达+Jagged-1组TNF-α、IL-1β水平降低(P<0.05)(表 3)。
2.5 各组细胞中ALP活性比较
与正常对照组相比,成骨诱导组ALP活性升高(P<0.05),而LPS组ALP活性降低(P<0.05);与LPS组相比,GAS5过表达组ALP活性升高(P<0.05),Jagged-1组ALP活性降低(P<0.05);与GAS5过表达组相比,GAS5过表达+Jagged-1组ALP活性降低(P<0.05);与Jagged-1组相比,GAS5过表达+Jagged-1组ALP活性升高(P<0.05)(表 4)。
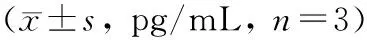
表 3 各组细胞上清液中TNF-α、IL-1β水平
表 4 各组细胞中ALP活性
2.6 各组细胞的矿化能力比较
与正常对照组相比,成骨诱导组钙化结节的数量和面积增加(P<0.05),而LPS组钙化结节的数量和面积降低(P<0.05);与LPS组相比,GAS5过表达组钙化结节的数量和面积升高(P<0.05),Jagged-1组钙化结节的数量和面积降低(P<0.05);与GAS5过表达组相比,GAS5过表达+Jagged-1组钙化结节的数量和面积降低(P<0.05);与Jagged-1组相比,GAS5过表达+Jagged-1组钙化结节的数量和面积升高(P<0.05)(图 2、表 5)。

图 2 各组hDPSCs茜素红染色(×200)
表 5 各组细胞中钙化结节的数量和面积
2.7 各组细胞中Notch通路和成骨相关蛋白的表达
与正常对照组相比,成骨诱导组细胞中NICD、Hes1、Hey1蛋白表达降低,RUNX2、OCN蛋白表达升高(P<0.05),LPS组细胞中NICD、Hes1、Hey1蛋白表达升高,RUNX2、OCN蛋白表达降低(P<0.05);与LPS组相比,GAS5过表达组细胞中NICD、Hes1、Hey1蛋白表达降低,RUNX2、OCN蛋白表达升高(P<0.05),Jagged-1组NICD、Hes1、Hey1蛋白表达升高,RUNX2、OCN蛋白表达降低(P<0.05);与GAS5过表达组相比,GAS5过表达+Jagged-1组NICD、Hes1、Hey1蛋白表达升高,RUNX2、OCN蛋白表达降低(P<0.05);与Jagged-1组相比,GAS5过表达+Jagged-1组NICD、Hes1、Hey1蛋白表达降低,RUNX2、OCN蛋白表达升高(P<0.05)(表 6,图 3)。
表 6 各组细胞中Notch通路和成骨相关蛋白的表达
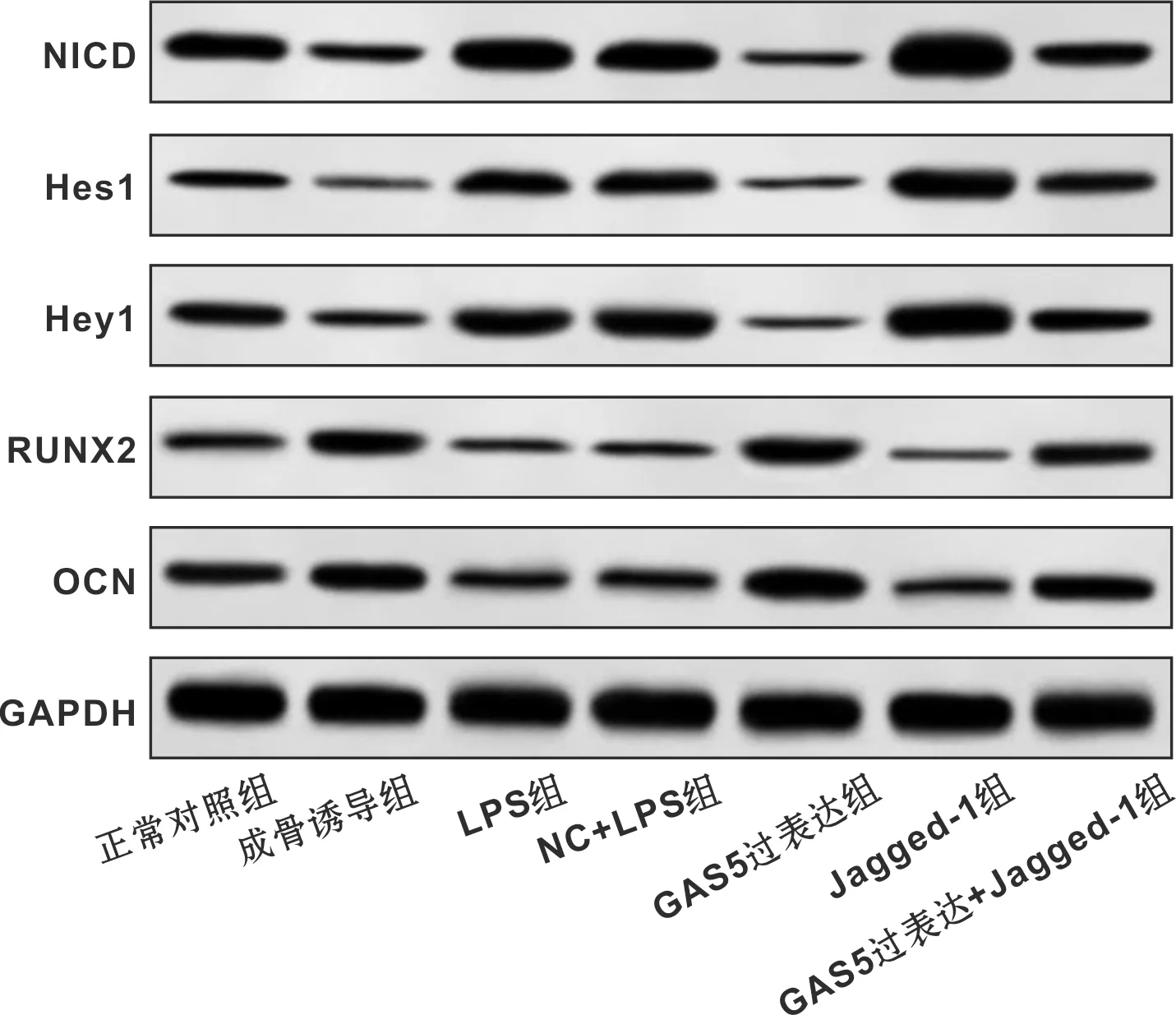
图 3 各组细胞中Notch通路和成骨相关蛋白的表达
3 讨 论
炎症环境不仅影响牙髓细胞的活力,还影响DPSCs的分化功能[9]。细菌主要成分LPS会诱导牙髓组织炎症导致牙髓损伤,同时抑制hPDLSCs的成骨分化[10]。lncRNA GAS5在hDPSCs中的特异性激活促进了成骨分化。这一发现提供了一种促进hDPSCs成骨能力的有效方法。
本研究结果显示,lncRNA GAS5在hDPSCs的成骨分化诱导过程中的表达增加;而在LPS处理的炎症条件下,hDPSCs的成骨分化被抑制时,其表达降低;同时研究发现过表达lncRNA GAS5后,hDPSCs的增殖活性和成骨分化能力均较LPS组增加,提示:过表达lncRNA GAS5在炎症环境中可促进hDPSCs成骨分化。在本研究中,LPS可抑制hDPSCs的增殖和成骨分化,而另有文献显示LPS能够促进hDPSCs向成牙本质细胞分化,并可通过增强hDPSCs进入牙髓时的粘附和迁移来影响牙髓愈合[11]。这与本文研究结果不一致,经查阅资料发现,LPS对hDPSCs的影响具有时间依赖性和浓度依赖性[12],结果不一致的原因可能与LPS的浓度有关。Notch信号通路调节牙髓组织中的多种生物过程,并可调节成骨分化[13]。研究发现,过表达NICD可抑制RUNX2、OCN的表达从而抑制成骨分化[14],表明Notch信号通路的激活与成骨分化的抑制有关。此外,Notch信号被视为针对感染和炎症驱动疾病的潜在治疗靶点[15],据报道,在LPS刺激或机械损伤后的牙髓细胞中发现了Notch信号的体外和体内激活[16]。本研究结果显示,经LPS处理的hDPSCs中,TNF-α、IL-1β水平和NICD、Hes1、Hey1表达均升高,且使用Jagged-1后上述指标进一步升高,说明在炎症环境下hDPSCs成骨分化受到抑制的过程中Notch信号通路被激活。Manokawinchoke等[17]的研究显示Jagged-1可激活人牙周膜基质细胞中的Notch信号通路,以剂量依赖性方式增强Hes1和Hey1 mRNA的表达,与本研究结果一致。然而,另有研究显示,Notch信号通路的激活对人牙髓和牙周膜细胞成骨分化和骨矿化具有促进作用[18],可能为Notch信号参与成骨分化的作用因细胞类型和分化阶段而异。
同时本研究发现,在lncRNA GAS5过表达组中NICD、Hes1和Hey1的表达较LPS组降低,而RUNX2和OCN的表达较LPS组升高,说明过表达lncRNA GAS5可抑制Notch信号通路的激活,促进hDPSCs成骨分化;此外,Jagged-1可明显减弱lncRNA GAS5过表达对Notch信号通路的抑制作用和hDPSCs成骨分化的促进作用,进一步提示过表达lncRNA GAS5可能通过抑制Notch信号通路激活,增强炎症环境下hDPSCs成骨分化。这与Li等[8]的研究一致,lncRNA GAS5可能通过抑制Notch信号通路改善LPS诱导的软骨细胞炎症损伤。
综上所述,lncRNA GAS5在hDPSCs成骨分化中上调,过表达lncRNA GAS5可增强炎症环境下hDPSCs成骨分化能力,其作用机制可能与抑制Notch信号通路的激活有关。但由于体内外环境存在较大差异,本研究结果尚需体内实验进一步验证。
