重金属铜和铅胁迫对东方山羊豆幼苗生长及生理特性的影响
2022-11-23穆海婷王英哲苗一凡郁伟杰徐博
穆海婷,王英哲,苗一凡,郁伟杰,徐博
(吉林农业大学林学与草学学院,吉林 长春 130118)
随着当今各行各业重工业技术与化工产业的大力发展,重金属污染问题日趋严重,并逐渐成了国际上最为严峻的环境污染问题之一[1]。重金属污染的土壤对植物的影响具有长期性、不可逆性以及影响范围广等特点,通过破坏细胞正常活动,进而影响植物生长发育,最终导致产量和质量下降[2-3]。铜(Cu)是重金属污染中常出现的元素,其参与植物生长发育中多种生理生化代谢过程,也是植物生长发育所需的微量元素,与植物的生长繁殖密切相关[4],但土壤中过量的铜会影响植物光合作用、抑制细胞代谢,造成植物毒害[5]。铅(Pb)作为常见的重金属元素,在土壤中被植物根系吸收,进入系统内部循环并在体内积累[6]。相关研究表明,过量的铅会增加植物吸水量及耗氧量,进而对植物造成毒害作用,破坏正常的生理功能[7-8]。近年来我国由重金属超标所引起的食品安全事故频发,人们对重金属污染的重视达到了空前的程度[9]。
东方山羊豆(Galega orientalis)又称饲用山羊豆,我国自1999年从哈萨克斯坦共和国引种以来,已在新疆、甘肃、宁夏等地种植,生长发育良好。该草为多年生豆科草本植物,具有鲜草产量高、易调制干草、抗逆性强等优点,是一种具有较好饲用价值的豆科牧草[10-11]。由于牧草具有生长发育快、产量高、生态保护等优点,近年来,牧草既可作为饲料,同时也可作为重金属污染土壤的修复植物[12]。但目前对东方山羊豆重金属胁迫的生理响应机制及修复重金属能力尚不清晰。通过研究东方山羊豆在重金属铜、铅胁迫下的生长、生理特性的变化,评价其抗逆性的强弱,不仅在理论上有指导意义,更重要的是能为植物修复土壤重金属污染和城市绿地的治理提供科学依据。
1 材料与方法
1.1 供试材料
供试材料为东方山羊豆,种子来源于中国农业科学院畜牧兽医研究所。
供试金属离子:CuSO₄·5H₂O、Pb(NO₃)₂均为分析纯试剂。
1.2 试验设计
试验于2021年10月在吉林农业大学开展,试验采用盆栽方式,供试基质由腐殖土、蛭石与珍珠岩按照4∶1∶1(体积比)的比例配成。分别设置5个Cu和Pb处理浓度,分别为:0、240、360、480、600 mg·kg-1,对照(CK)为去离子水。挑选处理好的种子种于塑料花盆中,于25℃培养箱中培养,试验期间用不同浓度的Cu和Pb溶液对东方山羊豆进行浇灌处理,每盆浇灌量为200 mL,间隔2~3 d进行一次,每处理重复5次。经过3周的浇灌处理,东方山羊豆生长情况基本正常。
1.3 测定指标及方法
1.3.1 生长指标测定 植株的芽长和根长的测定:在第7和15天,采集东方山羊豆完整植株,测量幼苗芽长、根长等指标。
植株鲜重的测定:在第7和15天,将东方山羊豆分为地上(芽)与地下(根)两个部分,分别称取鲜重。
1.3.2 生理指标测定 在胁迫第21天时,随机选取东方山羊豆幼苗新鲜叶片,测定生理生化指标。采用南京建成生物有限公司生产的试剂盒测定。
1)丙二醛含量的测定:使用试剂盒,采用硫代巴比妥酸(TBA)比色法,测定532 nm下的吸光度[13]。
2)过氧化物酶活性的测定:利用过氧化物酶催化过氧化氢反应原理,通过测定420 nm下吸光度的变化得出其酶活性[14]。
3)脯氨酸含量的测定:按照试剂盒抽取上清液,加入试剂后沸水浴30 min,流水冷却,测定520 nm下的吸光度。
4)可溶性糖的测定:采用蒽酮显色法测定可溶性糖的含量。将植物上清液、底物液和浓硫酸按一定比例充分混匀,置沸水浴中反应10 min,取出流水冷却后,于波长620 nm下测定各管吸光度值[15]。
5)可溶性蛋白测定:可溶性蛋白含量测定参考高俊凤[15]的考马斯亮蓝G-250染色法。首先,用牛血清蛋白制作标准曲线,将新鲜的待测叶片清洗干净,研磨成匀浆,吸取稀释后样品提取液1 mL加入试管中,加入5 mL考马斯亮蓝G-250试剂,充分混合放置5 min后,用分光光度计(上海菁华721可见分光光度计)测定其在波长595 nm时的吸光度,通过制作的标准曲线查出蛋白质含量。
以上各指标均分别称取新鲜叶片0.1 g,重复3次。
1.4 数据处理
用SPSS 20.0进行方差分析,用Excel 2016、GraphPad Prism进行绘图。
2 结果与分析
2.1 重金属胁迫对东方山羊豆生长的影响
2.1.1 不同浓度Cu、Pb胁迫下处理7 d对东方山羊豆生长的影响 随着Cu浓度的升高,东方山羊豆幼苗的根长在240 mg·kg-1最高、而根鲜重则逐渐升高,芽长、叶鲜重则呈先升后降的趋势(表1)。当Cu浓度达600 mg·kg-1时,与CK相比,东方山羊豆的根长、根鲜重上升幅度为43.22%、150.00%。随着Pb浓度的升高,东方山羊豆的芽长、根长、叶鲜重均呈现先增加后下降的趋势。其中根长的值在Pb处理浓度为480 mg·kg-1时,与CK相比增加16.47%,而芽长在其他处理中的值均低于CK。
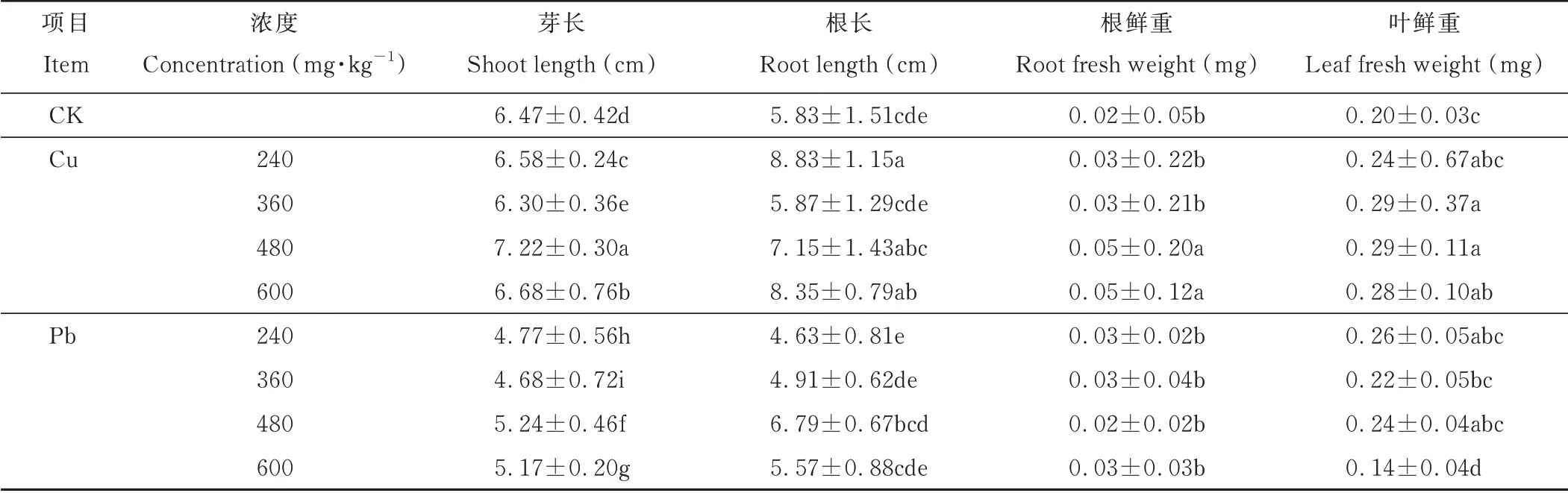
表1 不同Cu、Pb浓度胁迫7 d对东方山羊豆生长的影响Table 1 Effects of different concentrations of Cu and Pb stress for 7 days on the growth of G.orientalis
2.1.2 不同浓度Cu、Pb胁迫下处理15 d对东方山羊豆生长的影响 重金属胁迫15 d后,如表2所示,在Cu浓度达600 mg·kg-1时,东方山羊豆幼苗的芽长、叶鲜重与CK相比,分别增加了13.40%、95.45%。根长、根鲜重在Cu胁迫处理中则与CK差异不显著。随着Pb浓度的增加,东方山羊豆各生长指标呈先上升后下降趋势,在处理浓度为480 mg·kg-1时,芽长、根鲜重、叶鲜重与CK相比均呈现上升趋势,而根长在处理浓度为600 mg·kg-1时,与CK相比降低23.60%。
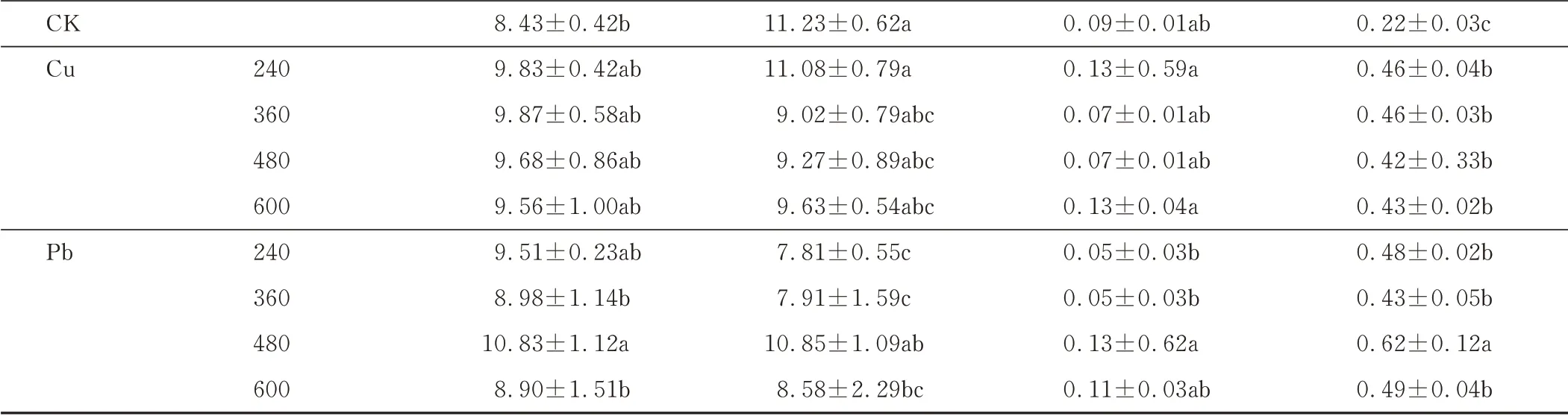
表2 不同Cu、Pb浓度胁迫15 d对东方山羊豆生长的影响Table 2 Effects of different concentrations of Cu and Pb stress for 15 days on the growth of G.orientalis
2.2 重金属胁迫对东方山羊豆生理生化特征的影响
2.2.1 不同浓度Cu、Pb胁迫对东方山羊豆丙二醛(malondialdehyde,MDA)含量的影响 MDA是植物在逆境胁迫下膜脂过氧化的产物,测定重金属胁迫下植物体内MDA的累积量,可在一定程度上反映植物质膜受破坏的程度[16]。由图1可以看出,重金属Cu、Pb胁迫下,随着处理浓度的增加,东方山羊豆幼苗体内的MDA含量总体呈递增趋势。Cu胁迫下,随着胁迫浓度的增加,MDA含量与CK相比呈显著差异(P<0.05),240、360、480 mg·kg-1处理下的MDA与CK相比,分别增加了16.34%、73.02%、77.95%,处理浓度为600 mg·kg-1时的MDA含量最高,比CK增加1倍 多。当Pb浓 度 小 于480 mg·kg-1,植 物 体 内MDA含量呈上升趋势,当处理浓度为600 mg·kg-1时,东方山羊豆幼苗的MDA含量出现明显下降。
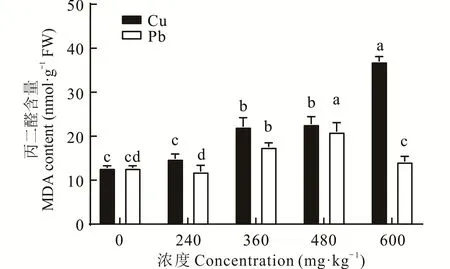
图1 不同浓度Cu、Pb胁迫下丙二醛含量的变化Fig.1 Changes in malondialdehyde(MDA)content under different concentrations of Cu and Pb stress
2.2.2 不同浓度Cu、Pb胁迫对东方山羊豆脯氨酸(proline,PRO)含量的影响 PRO以游离状态广泛存在于植物中。当植物受到胁迫时,植物体内的PRO含量一般来说会显著增多,PRO增加量一定程度上反映了植物的抗逆性[17]。图2可以看出在未受到胁迫时,东方山羊豆所含的PRO含量并不多,但受到重金属胁迫时,随着胁迫浓度的增加,植物体内的PRO含量逐渐增加,且差异显著(P<0.05)。在Cu胁迫下,随着浓度的升高,在240、360、480 mg·kg-1时,东方山羊豆体内的PRO相对于CK增长率分别为49.92%、76.98%、125.43%,当处理浓度为600 mg·kg-1时,植物体内的PRO含量为CK的3.23倍。在Pb胁迫下,随着浓度的升高,东方山羊豆PRO含量的增长率分别为39.50%、63.99%、104.12%、157.83%。总体来看,随着胁迫浓度的增加,东方山羊豆体内游离PRO含量呈持续累积的状态。
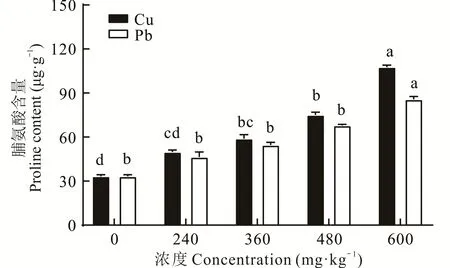
图2 不同浓度Cu、Pb胁迫下脯氨酸含量的变化Fig.2 Changes in proline content under different concentrations of Cu and Pb stress
2.2.3 不同浓度Cu、Pb胁迫对东方山羊豆过氧化物酶(peroxidase,POD)活性的影响 东方山羊豆幼苗的POD活性随着不同重金属胁迫浓度的增加呈现出不同的变化趋势(图3),随着Cu胁迫浓度的增加,东方山羊豆叶片中的POD活性呈上升趋势,显著高于对照(P<0.05)。在Cu处理浓度为600 mg·kg-1时,POD活性与CK相比,增加383.26%。随着Pb胁迫浓度的增加,POD活性增长趋势平缓,Pb胁迫下幼苗的POD活性仅在浓度为360、600 mg·kg-1时显著高于对照组。
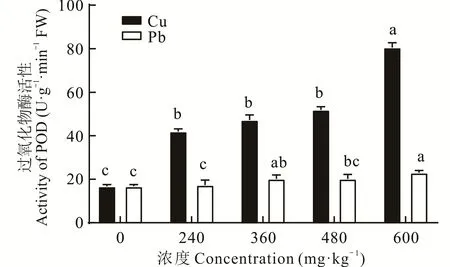
图3 不同浓度Cu、Pb胁迫下过氧化物酶活性的变化Fig.3 Changes in peroxidase activity under different concentrations of Cu and Pb stress
2.2.4 不同浓度Cu、Pb胁迫对东方山羊豆可溶性蛋白含量的影响 由图4可以看出,重金属Cu胁迫下,随着处理浓度的升高,东方山羊豆叶片中的可溶性蛋白含量出现先升后降的趋势,Cu胁迫浓度在240 mg·kg-1时,植物的可溶性蛋白含量显著高于600 mg·kg-(1P<0.05),并在此时达到最大值,而随着胁迫浓度增加,可溶性蛋白含量逐渐降低,360、480、600 mg·kg-1时,可溶性蛋白含量与CK相比分别降低6.49%、11.69%、42.86%。Pb胁迫下,随着处理浓度的升高,可溶性蛋白含量呈先下降后上升趋势,与CK相比,在处理浓度为600 mg·kg-1时,增加78.35%。结果表明,较低浓度的Cu处理有助于东方山羊豆的可溶性蛋白含量积累,而重金属浓度过高会导致其含量下降,这可能是过高的重金属胁迫会对可溶性蛋白的合成造成干扰。
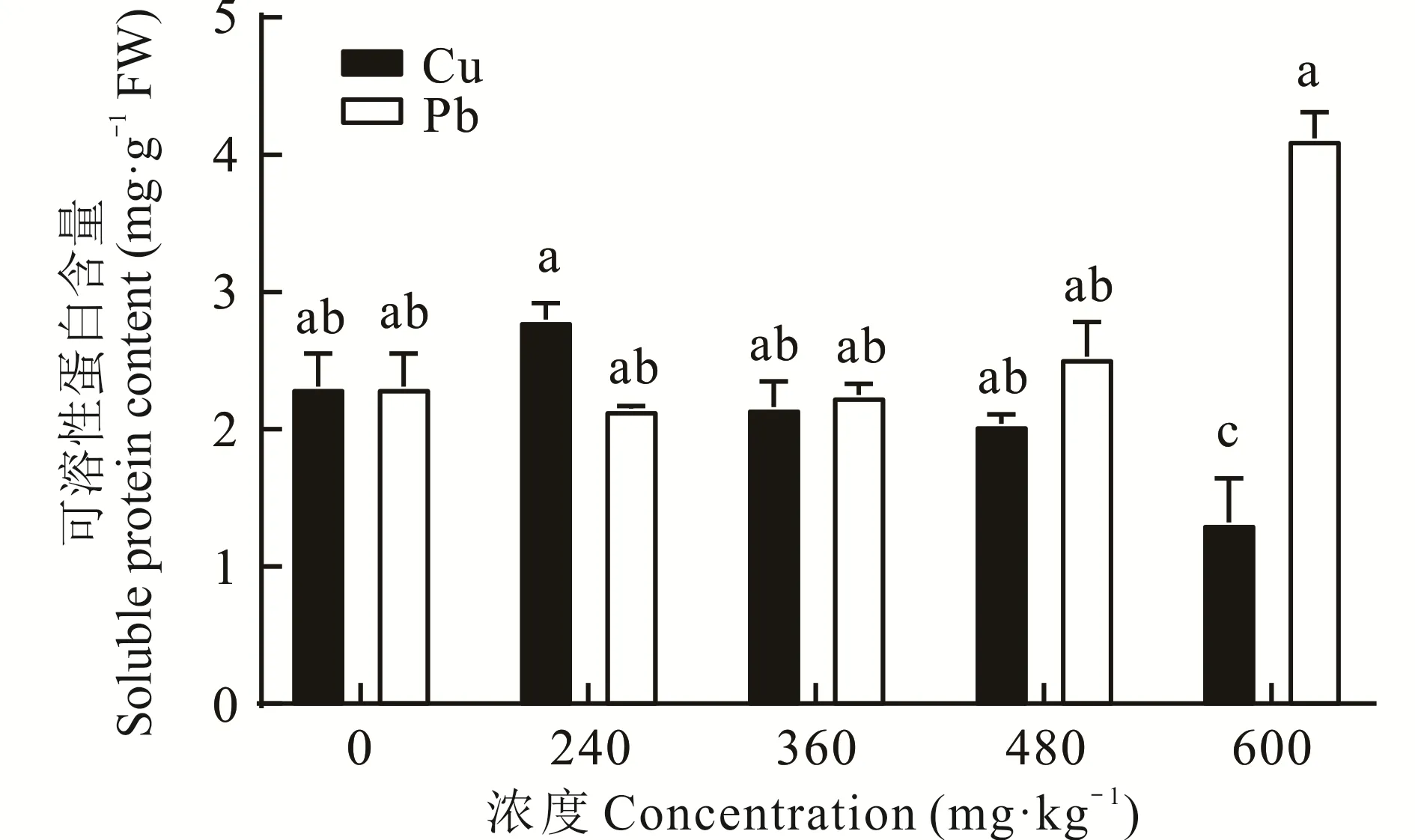
图4 不同浓度Cu、Pb胁迫下可溶性蛋白含量的变化Fig.4 Changes in soluble protein content under different concentrations of Cu and Pb stress
2.2.5 不同浓度Cu、Pb胁迫对东方山羊豆可溶性糖含量的影响 由图5可以看出,可溶性糖含量随着Cu、Pb胁迫浓度的增加,呈现出不同的变化趋势。随着Cu胁迫浓度的增加,植物的可溶性糖含量总体呈先下降后上升趋势,在Cu浓度低于480 mg·kg-1时,东方山羊豆的可溶性糖含量均低于CK。与CK相比,在Cu胁迫浓度为600 mg·kg-1时,植物的可溶性糖含量仅上升14.22%。在Pb胁迫下,东方山羊豆体内的可溶性糖含量呈先上升后下降的趋势,但均高于CK,分别增加了4.30%、37.64%、40.95%、13.86%。说明在Pb胁迫下,东方山羊豆具有更好的渗透调节能力。
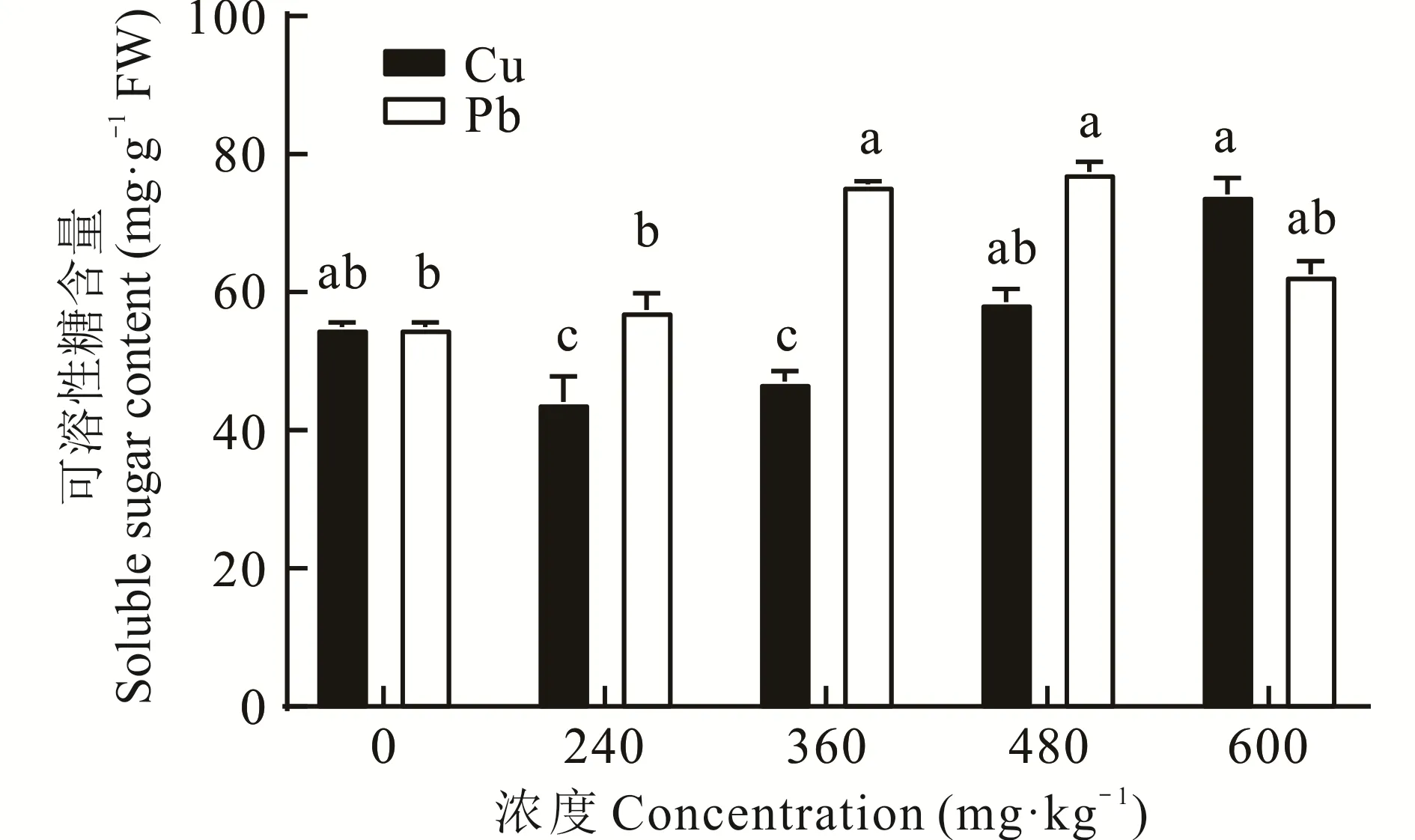
图5 不同浓度Cu、Pb胁迫下可溶性糖含量的变化Fig.5 Changes in soluble sugar content under different concentrations of Cu and Pb stress
3 讨论与结论
重金属污染对环境与生物正常生命活动具有较强的破坏作用,具有长期性、潜伏性和不可逆性等特点[18]。近年来环境中铜污染日益严重,主要来源于工业“三废”的排放,土壤中铜含量普遍提高[19]。土壤中的铅主要来源于工业废物的大量排放、大规模的采矿作业、农药和化肥广泛使用等,使土壤中的铅过量积累,通过植物根系进入植物体内,最终产生严重的毒害作用[20]。在Cu不同浓度胁迫7 d时,东方山羊豆幼苗未受到明显影响,根长、根鲜重、叶鲜重整体呈上升趋势,在Cu胁迫第15天时,东方山羊豆的幼苗芽长、叶鲜重相较于CK略有增加。总体来说东方山羊豆在不同浓度Cu、Pb胁迫下,幼苗生长受抑制规律不明显,在重金属污染土壤上,大多数作物的生长都会受到一定的抑制作用,而随着重金属浓度的增加会导致植物生理代谢的改变,甚至阻碍生长。这与其他学者的研究结果一致[21-22]。土壤重金属污染植物修复的基本原理是通过植物吸收作用将重金属从土壤中携出,不同植物对重金属的累积量不同,相同植物不同品种之间因存在基因型差异,对重金属的累积量也有显著的差异[23]。
植物机体内MDA的积累在一定程度上反映了植物受伤害程度[24]。本研究中,两种重金属胁迫下,东方山羊豆的MDA含量总体呈上升趋势,而在Pb胁迫为600 mg·kg-1时,MDA含量开始下降,这可能是保护酶对植物应对重金属胁迫的作用[16]。植物的抗逆性与POD的表达和活性密切相关,植物通过增强抗氧化酶活性及增强活性氧代谢来增强植物的耐受性[25]。在本研究中,Cu胁迫下,POD活性随着胁迫浓度的增加而上升,Pb胁迫下幼苗的POD活性仅在浓度为360、600 mg·kg-1时显著高于对照组。陆奇杰等[26]在研究铜胁迫对茅苍术(Atractylodes lancea)光合特性及生理指标的影响时发现随着铜浓度的增加,其POD活性随着胁迫浓度的增加而上升,本试验结果与之相似。
PRO、可溶性蛋白、可溶性糖等作为渗透调节物质,在植物面对胁迫时起到重要作用,当植物本身面临胁迫时,会增加自身的含量,以提高抗逆性[27]。脯氨酸是调节植物水分胁迫条件下细胞水势的物质之一,在逆境条件下变化较大,受到胁迫后PRO含量的积累与CK相比会大幅上升,在一定程度上也反映了植物对逆境条件的忍受能力[28]。随着两种胁迫浓度的增加,东方山羊豆的PRO含量呈上升趋势,变化趋势与MDA相似。本研究结果表明,在Cu胁迫下的可溶性糖呈先下降后上升的趋势、可溶性蛋白含量总体呈下降趋势,东方山羊豆叶片中的可溶性糖含量在Pb胁迫下,随着胁迫浓度的增加,总体呈先升后降趋势,而可溶性蛋白则呈先下降后上升的趋势。在Cu胁迫浓度较低时,可溶性糖含量下降,可能是光合作用产生的淀粉在运输过程中受到了阻碍的结果,在Pb胁迫浓度较高时,可溶性糖含量大幅下降,可能是由于抑制植物呼吸作用和光合作用,有关糖和氮代谢的酶活性降低,导致植物细胞生化和生理活动降低,所以渗透调节物质的量不能长期保持在高位,在升高后会慢慢降低[29]。随着重金属Cu胁迫的增加,东方山羊豆叶片中可溶性蛋白含量呈下降趋势,可能是叶片的细胞结构受到破坏,阻碍了蛋白质的合成,使原有蛋白质的分解速度加快,导致细胞代谢酶和可溶性蛋白质受到破坏。
综上所述,在短时间内随着胁迫浓度的增加,Cu和Pb两种胁迫对东方山羊豆的生长未表现出明显抑制作用。东方山羊豆对重金属的反应不同,但总趋势是一致的。随着Cu、Pb胁迫浓度的增加,东方山羊豆通过调节自身的生长状况、渗透调节及酶活性的变化来应对一定范围内的重金属胁迫,重度胁迫会对其产生不良的影响,但达不到致死程度,说明东方山羊豆对Cu、Pb胁迫耐受力较强,作为多年生豆科植物,其共生的根瘤菌可以固定并改变土壤中重金属的形态,降低土壤重金属污染;另一方面根瘤菌具有固氮作用,可恢复土壤养分[30]。东方山羊豆在土壤铜、铅污染修复中的应用具有一定的潜力,可以作为候选修复植物在实际铜、铅污染土壤中进一步深入研究。
