柳枝稷Hsp70 家族基因鉴定与生物信息学分析
2022-11-18方志刚王玉龙蔡庆生
宋 刚,方志刚,王玉龙,蔡庆生,徐 彬
(1. 江苏农林职业技术学院茶与食品科技学院, 江苏 句容 212400;2. 南京农业大学生命科学学院, 江苏 南京 210095;3. 喀什大学生命与地理科学学院, 新疆 喀什 844099;4. 南京农业大学草业学院, 江苏 南京 210095)
热激蛋白(heat shock proteins, Hsps)是一类受环境胁迫诱导表达的蛋白[1],其中Hsp70 家族(分子量在68~78 kDa)是在生物体内分布最广和研究最多的一类热激蛋白,具有典型的Hsps 的结构特征[2],也是Hsps 中结构和功能最保守的蛋白家族[3]。Hsp70s 结构上主要由核苷酸结合结构域(nucleotide binding domain, NBD)、底物结合结构域(substrate binding domain, SBD)与可变C 端区组成,其可变C端区柔韧、有利于底物与SBD 结合[4]。
当植物遭遇非生物胁迫时,能够快速激活特定的Hsp70s。作为分子伴侣,Hsp70s 能帮助蛋白质在不利环境中正确折叠,阻止蛋白质聚集或失活,在细胞蛋白质量控制和降解系统中起着中心作用,以促进细胞耐受胁迫环境[5-6]。一些Hsp70s 在条件适宜时也发挥着“管家”蛋白的关键作用[7]。例如,定位于内质网(endoplasmic reticulum, ER)上部分Hsp70s基因编码的BiPs (binding proteins),具有执行重要蛋白合成、折叠和装配的功能[8]。研究表明,Hsp70基因表达水平与植物耐热性之间存在显著的正相关关系[9],这已经在辣椒(Capsicum annuum)[1,3]、拟南芥(Arabidopsis thaliana)[10]、水稻(Oryza sativa)[11]中得到证实。植物基因组中Hsp70基因通常具有多拷贝,模式植物拟南芥[12]基因组有19 个Hsp70基因,其中3 个编码BiP,且3 个AtBiPs 表达均受干旱、冷、病虫害等环境胁迫的调控[13-14]。水稻含有32 个Hsp70基因,5 个是BiP[15]。其中,OsBiP1主要在种子成熟过程中表达,影响种子的表型和胚乳细胞的胞内结构,参与种子发育调控[16];OsBiP3参与调 控Xa21 介导的植物抗病免疫力[17];OsBiP4和OsBiP5与内质网胁迫响应高度关联[18]。大豆(Glycinemax)[19]中鉴定出61 个Hsp70基因,玉米(Zea mays)[20]中是35 个,芝麻(Sesamum indicum)[21]为21 个,马铃薯(Solanum tuberosum)[22]中则有20 个。
柳枝稷(Panicum virgatum)是一种多年生禾本科C4草本植物,原产于北美洲20°~60° N,适应性强。柳枝稷根系发达,可在边际土地和污染土地生长,其生物量所含纤维素能用于生产乙醇和丁醇等生物燃料[23-25],早在1992 年柳枝稷就被美国能源部确立为生物能源模式植物[26]。此外,由于柳枝稷生物产量大,根系发达,且能够耐受中低浓度镉污染[27-29],因此柳枝稷也可作为镉污染土地修复的重要植物物种[30-31]。目前,柳枝稷全基因组测序已经完成[32],但关于柳枝稷Hsp70基因家族(PvHsp70s)研究尚未有报道。本文通过生物信息学方法鉴定出PvHsp70家族基因成员,分析了基因结构、染色体分布、进化和表达模式,以期挖掘柳枝稷逆境响应基因,为研究PvHsp70基因功能与柳枝稷分子改良提供理论依据。
1 材料与方法
1.1 柳枝稷PvHsp70 基因鉴定
柳枝稷基因组与蛋白数据下载自Phytozome(Panicum virgatumv4.1, http://phytozome.jgi.doe.gov/)数据库网站。从Pfam (http://pfam.xfam.org)下载Hsp70 蛋白结构域的隐马尔可夫模型(hidden markov modle,HMM) 种子文件PF00012,用于本地基因库的PvHsp70基因鉴定。通过HMMER 软件(http://hmmer.org)在柳枝稷全基因组蛋白序列中搜索含有该保守结构域的序列,设置E-value < 0.001。候选序列提交SMART (http://smart.embl.de)、Pfam 蛋白数据库进行Hsp70 结构域验证,并从柳枝稷基因组数据中提取验证确认的蛋白和基因编码序列(coding sequence, CDS)。使用在线软件WOLFPSORT (https://www.genscript.com/wolf-psort.html)预测分析氨基酸序列大小、分子量、等电点等理化特征。
1.2 序列比对、系统进化树构建、基因结构及保守基序分析
从拟南芥TAIR 数据库(http://www.arabidopsis.org)和RGAP 数据库(http://rice.plantbiology.msu.edu)下载拟南芥和水稻Hsp70 蛋白序列,通过MEGA7.0中clustalw (http://www.megasoftware.net)进行多序列比对,比对结果采用邻接法[13]构建系统发育进化树,Bootstrap 值设置1 000,其余参数使用默认值。通过在线工具GSDS2.0 (http://gsds.cbi.pku.edu.cn/),分析PvHsp70基因的外显子-内含子结构。利用MEME在线软件(http://meme.nbcr.net/meme/) 鉴定保守基序(motif),motif 的最大数目设定为10,长度6 到100个氨基酸,其余参数默认。在CDD 数据库(https://www.ncbi.nlm.nih.gov/cdd)分析motif 结构特征。
1.3 基因启动子分析与染色体定位、基因复制
使用PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html)数据库分析PvHsp70基因上游2 kb 启动子序列中顺式作用元件(cis-acting element)。使用Mapchart 软件(https://www.wur.nl/en/show/Mapchart.html)绘制基因染色体定位图。参照Gu 等[33]比对条件(a.比对的短序列占较长的序列的70%以上;b.两序列比对相似性大于70%),使用makeblastdb 和blastall 命令筛选复制基因。通过MCsacnx 软件(http://chibba.pgml.uga.edu/mcscan2/MC ScanX.zip)分析PvHsp70基因共线性关系,并绘制circos 圈图。
1.4 基因表达分析
分别在柳枝稷microarray 数据库网站(https://switchgrassgenomics.noble.org)中下载PvHsp70s基因在柳枝稷的根、茎、叶、花等不同组织的14 个不同发育阶段以及授粉后不同发育天数(days after pollination, DAP) 7 个阶段的表达量数据(FPKM 值),并从镉胁迫柳枝稷根系RNA-seq 结果[34]中提取PvHsp70s的表达量数据(FPKM 值)。利用TBtools 软件(https://github.com/CJ-Chen/TBtools/releases)分析上述基因表达并绘制表达量热图。
2 结果与分析
2.1 柳枝稷PvHsp70 基因鉴定
经HMMER 搜索和SMART、PFAM 数据库结构域确认,除去冗余和缺失序列,存在可变剪切的仅选取1 个最长的转录本,并剔除分子量预测小于60 kDa 的转录本,共在柳枝稷基因组中鉴定出41 个PvHsp70基因。由于柳枝稷为四倍体(2n= 4x= 36),有两个亚基因组K 和N,基本染色体数为9[35],因此,按转录本信息中染色体序号从小到大、亚基因组先K 后N 及基因组位置从小到大的规则将33 个分子量在60~80 kDa 之间的命名为PvHsp70-1~PvHsp70-33。参照拟南芥[13]和水稻[16]中分类,将其余8 个分子量大于80 kDa 的Hsp70s,归为Hsp110/SSE 亚家族成员,命名为PvHsp110-1~PvHsp110-8。PvHsp基因的CDS 序列长度在1 626 (PvHsp70-24)~2 694 bp (PvHsp110-1、PvHsp110-2) 之 间,氨 基 酸数量范围在548 (PvHsp70-23)~897 aa (PvHsp110-1、PvHsp110-2),分子量介于60.55 (PvHsp70-23)~98.95(PvHsp110-1)。PvHsp70-4、PvHsp70-21、PvHsp70-24和PvHsp70-33 这4 个蛋白预测等电点大于7.0,剩余蛋白等电点都小于7.0。亚细胞定位预测显示,PvHsp70s 中有22 个定位在细胞质(cyto)、7 个定位于内质网(ER)、6 个定位于质体(plasto)、6 个定位在线粒体(mito)上(表1)。
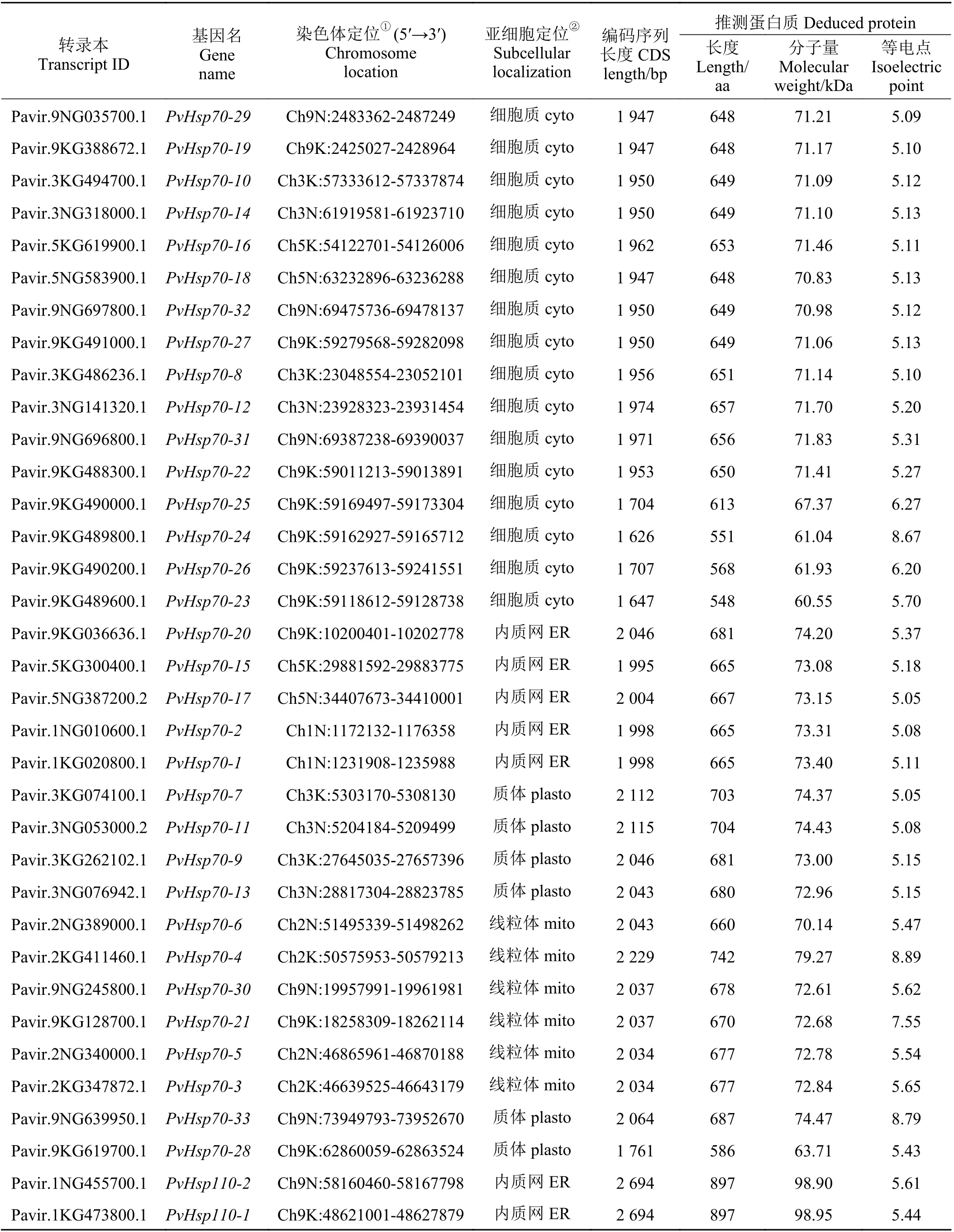
表1 柳枝稷PvHsp70 基因鉴定、序列特征和亚细胞定位Table 1 List of PvHsp70 genes identified in switchgrass, their sequence characteristics and subcellular localization
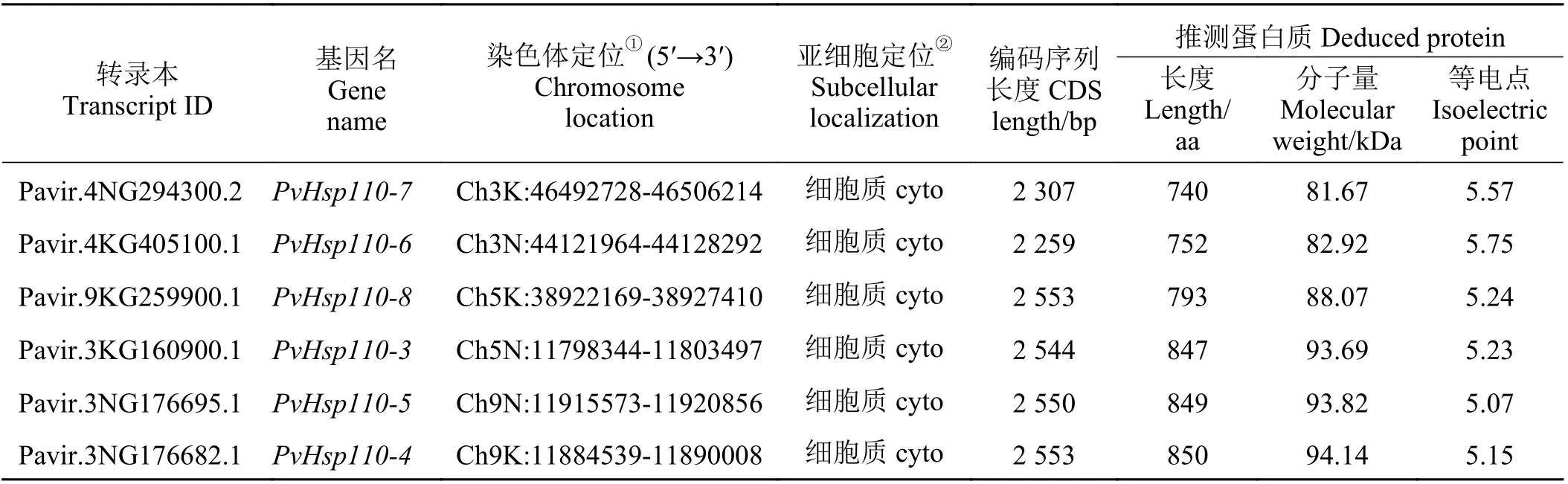
续表1Table 1 (Continued)
2.2 PvHsp70 中的BiP 基因分析
BiP 蛋白序列包含一个C 端ER 驻留信号,植物中这个信号序列通常是“HDEL”[36]。 41 个PvHsp70中有6 个符合此特征,且亚细胞定位预测均在ER上。经CDD 在线工具数据库(KOGv1.0) 进行序列分析,其中2 个含有KOG0104 (Molecular chaperones GRP170/SIL1, HSP70 superfamily)结构域,另4 个含有BiP基因结构域特征KOG0100 (Molecular chaperones GRP78/BiP/KAR2, HSP70 superfamily) (表2)。

表2 柳枝稷PvHsp70 基因家族中BiP 基因分析Table 2 Analysis of BiP genes in the PvHsp70 family of switchgrass (Database: KOGv1.0)
2.3 系统进化、基因结构和保守基序分析
PvHsp70 系统进化树(图1A)显示,41 个成员一共聚类成6 个组。组1 含有16 个(占39%)成员,数量最多;组5 仅含2 个成员(4.9%),数量最少。相同组内PvHsp70 成员基因结构和保守基序(motif)的类型、数量与顺序分布基本相似,不同组之间则有明显区别(图1B、C)。组1 中,PvHsp70-12和PvHsp70-16具有2 个内含子,其他14 个基因都含有1 个内含子。组2 成员基因内含子数为0、1 和7 三种类型,组3 中是7 和8,组4 中为4 和5,组5 中则是0 和2,组6 中为8、9、13 和14 四种结构形式。同组内成员基因内含子数量差异较大,预示着同组基因在功能上可能具有多样性特征。MEME 网站分析出的10个motif 长度在21~80 aa (表3),motif 1、2、4、5、6、7 和10 属于Hsp70 核苷酸结构域(NBD of the sugar inase/HSP70/actin superfamily),PvHsp70 成员都具有上述motif 中的全部或大部。motif 9 为分子伴侣Dank (molecular chaperone DnaK)结构域,只存在1~5 组成员(PvHsp70-1~PvHsp70-33) 中,因此可把这5 组成员划为分子伴侣Dank 亚家族。组6 中成员(PvHsp110-1~PvHsp110-8) 都不含motif 9,也没有motif 3 或8 [ 均为Hsp70 蛋白结构域(heat shock 70 kDa protein)]存在,并非分子伴侣Dank 亚家族成员,这与2.1 中依据分子量将其划分为Hsp110/SSE 亚家族的结果是吻合的。综上可知,PvHsp70 家族成员分属于分子伴侣Dank 亚家族(PvHsp70-1~PvHsp70-33)和Hsp110/SSE 亚家族(PvHsp110-1~PvHsp110-8)。

表3 柳枝稷PvHsp70 蛋白的基序序列组成Table 3 Basic sequence composition of motifs of PvHsp70 proteins in switchgrass
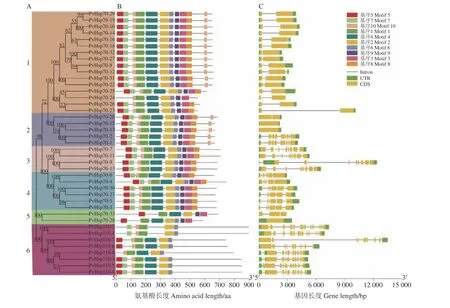
图1 柳枝稷PvHsp70 成员系统进化、基因结构与保守基序组成Figure 1 Phylogenetic relationships, motif composition, and gene structure of PvHsp70 proteins of switchgrass
2.4 柳枝稷、水稻和拟南芥Hsp70 基因的进化关系分析
将柳枝稷(41 个)、拟南芥(18 个)、水稻(32个)的Hsp70s 构建了系统发育进化树(图2),结果显示:91 个Hsp70 蛋白可聚成4 类,根据2.1 亚细胞定位预测结果,分别是细胞质Hsp70 蛋白(CⅠ和CⅡ区)、内质网Hsp70 蛋白(ERⅠ和ERⅡ区)、线粒体Hsp70 蛋白(M 区)和质体Hsp70 蛋白(PⅠ和PⅡ区)。细胞质Hsp70 蛋白数量达到47 个(柳枝稷22 个,水稻16 个,拟南芥9 个),占总数的51.6%,表明细胞质是Hsp70 的主要功能区域。此外,54 个Hsp70基因有同源关系,3 对为直系同源基因(AT3G12580与Os03g60620,PvHsp110-2与Os02g48110,PvHsp70-20与Os03g50250),24 对属于旁系同源基因(柳枝稷内17 对,拟南芥内4 对,水稻内3 对)。
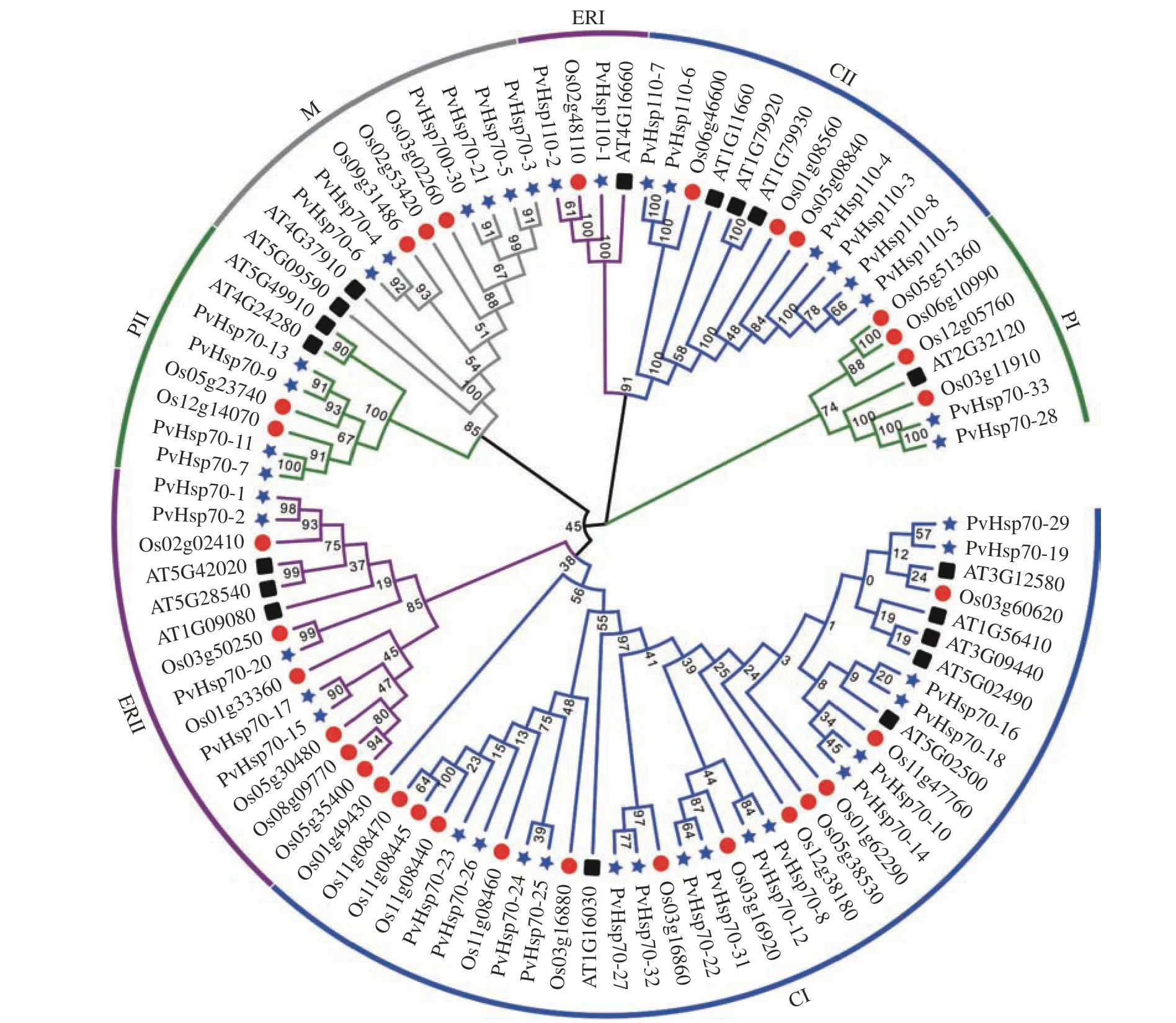
图2 柳枝稷、水稻及拟南芥Hsp70 蛋白系统进化树Figure 2 Phylogenetic tree of Hsp70 proteins from switchgrass, rice, and Arabidopsis
2.5 柳枝稷PvHsp70 基因启动子序列分析
柳枝稷PvHsp70基因上游2 kb 启动子区主要存在两类顺式作用元件,即响应非生物胁迫相关作用元件和激素应答顺式作用元件(表4)。响应非生物胁迫相关作用元件主要有:响应热胁迫的HSE 和CCAAT-box 元件、响应低温胁迫的LTR 元件、响应干旱胁迫的相关作用元件MBS、与机械损伤有关的WUN-motif 元件、植物响应胁迫的TC-rich repeats元件。激素应答顺式作用元件主要有:生长素响应元件(AuxRR-core)、乙烯反应作用元件(ERE)、脱落酸作用反应元件(ABRE,motif IIb)、水杨酸作用反应元件(TCA-element)、 赤霉素作用响应元件(GARE-motif, TATC-box)、茉莉酸甲酯作用元件(CGTCA-motif)和植物激素反应作用元件(TGA-box)。上述结果表明PvHsp70基因与多种胁迫密切相关。

续表4Table 4 (Continued)
2.6 柳枝稷PvHsp70 基因染色体定位和基因复制
PvHsp70基因在染色体上分布显示,柳枝稷18 条染色体(K 和N 亚基因组各9 条)中12 条上有PvHsp70基因,且分布不均衡(图3)。其中,09K 染色体上PvHsp70数量最多(11 个,占基因总数的26.8%),其次是03N 染色体(6 个),03K、09N 染色体各有5 个,01K、01N、02K、02N、05K、05N 染色体各有2 个,04K、04N 染色体只含1 个。MCsacnx 查找和circos 图呈现出柳枝稷染色体上PvHsp70基因的共线性关系(图4),有32 个PvHsp70基因具有共线性关系,形成21 对具有复制关系基因(PvHsp70-1和PvHsp70-2、PvHsp110-6和PvHsp110-7等)。多条红色线连接基因可能经历多次复制事件,如PvHsp-16与PvHsp70-8、PvHsp70-12、PvHs70-18、PvHs70-25四个基因。此外,MCsacnx 查找发现03N 染色体上PvHsp110-4和PvHsp110-5为一对串联复制基因。

图3 柳枝稷PvHsp70 基因在染色体上的分布Figure 3 Chromosomal distribution of PvHsp70 genes of switchgrass
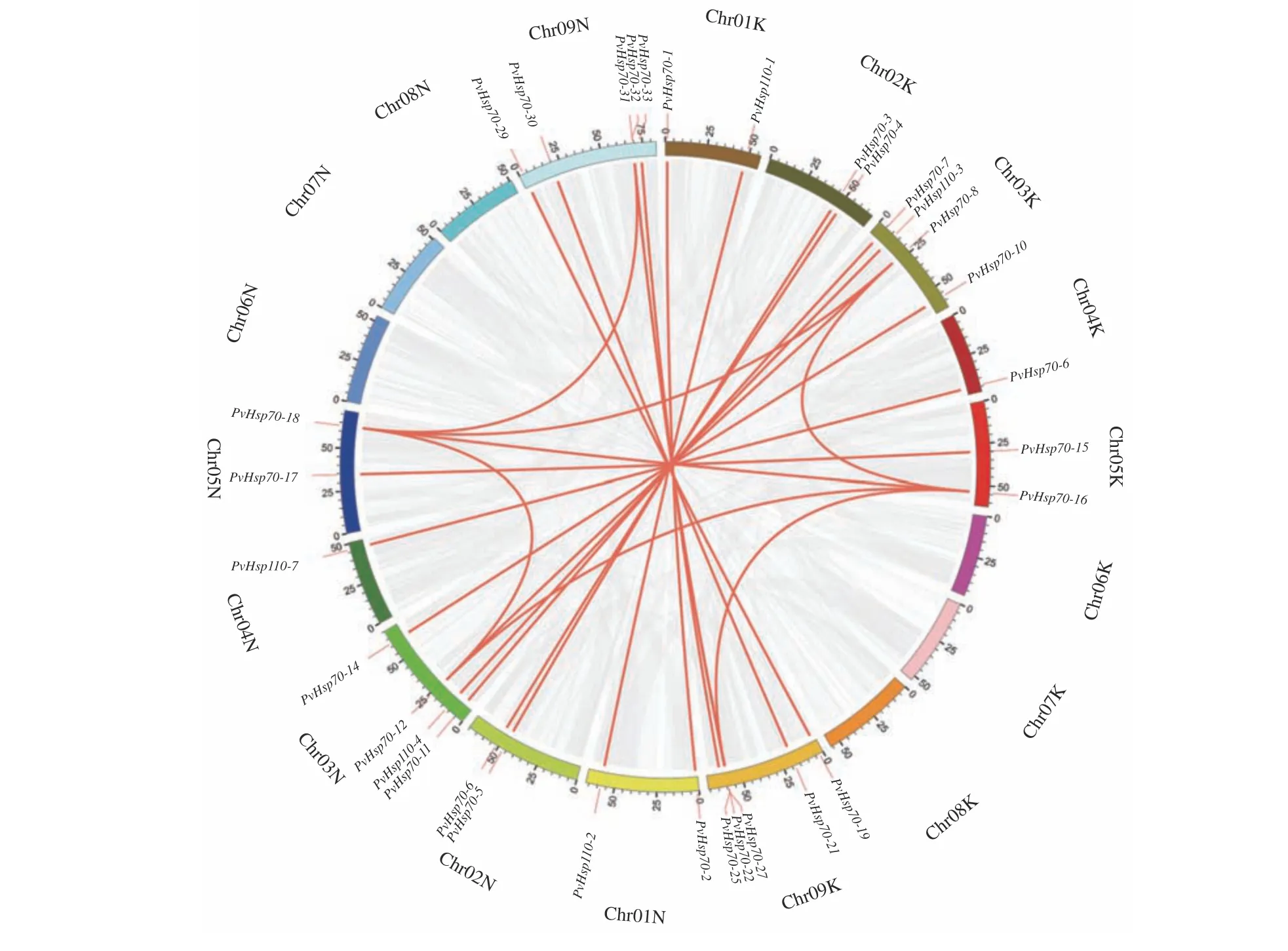
图4 柳枝稷PvHsp70 基因共线性Circos 图Figure 4 Circos diagram of collinearity relationships of PvHsp70 genes in switchgrass
2.7 柳枝稷PvHsp70 基因表达模式分析
从PvHsp70基因在生长和发育21 个阶段的表达量热图(图5) 可看出,PvHsp70家族基因的不同成员之间表达量具有差异。其中,PvHsp70-2和PvHsp70-10在各个阶段表达量均很高,PvHsp70-22和PvHsp70-31明显在种子发育阶段中具有更高的表达。PvHsp70-5和PvHsp70-21在花器官中表达量明显高于其他阶段。PvHsp70-18在根中表达量较高,叶中表达量较高的有PvHsp70-4、PvHsp70-7、PvHsp70-9和PvHsp70-13等。总体上,在种子发育形成过程中,多个PvHsp70的表达量要高于其他生长阶段,说明其可能参与种子发育调控。
此外,根据CdCl2处理(0、6、24 h)后的柳枝稷根系RNA-seq 数据,共发现41 个PvHsp70基因中有29 个在镉胁迫后发生上调表达,其中PvHsp70-16、PvHsp70-18、PvHsp70-32和PvHsp70-27四个基因上调表达倍数较高,而PvHsp70-15是唯一在经6 h、24 h 镉胁迫下表达量持续上升的基因。另外,有4 个基因在镉处理下持续下调表达(PvHsp70-3、PvHsp70-5、PvHsp110-6和Hsp110-7),还有7 个基因(PvHsp70-9、PvHsp70-26、PvHsp70-24、PvHsp70-20、PvHsp70-17、PvHsp70-25和PvHsp70-23) 表 达 量 较低,基本不受镉诱导表达(图6)。

图6 柳枝稷根系PvHsp70 基因在镉胁迫下的表达量热图Figure 6 Heatmap of expression of PvHsp70 genes in switchgrass roots under cadmium stress
3 讨论
Hsp70基因是在生物体内分布最广和研究最多的一类基因家族。本研究在柳枝稷基因组中共鉴定命名了41 个PvHsp70基因,通过分子量大小以及motif 结构域特征分析,33 个成员(PvHsp70-1~PvHsp70-33)被归于分子伴侣Dank 亚家族,其余8 个成员(PvHsp110-1~PvHsp110-8)被划为HSP110/SSE 亚家族,在拟南芥[12]和水稻[15]中也存在类似划分。无或仅有1 个内含子的基因通过可变剪切实现单个基因多个蛋白质翻译的可能性较小,在进化中其功能会较保守和单一[37]。PvHsp70基因中无内含子或含1 个内含子的成员共19 个,占基因总数的46.3%。在玉米[20]、大豆[19]、芝麻[21]及辣椒[38]中此比例分别是37.1%、34.4%、42.8%和33.3%,与本研究中基本接近,说明柳枝稷与其他物种中具有相似比例且高度保守的Hsp70基因。不同物种间进化分析发现,同一进化枝的柳枝稷、水稻直系同源基因(PvHsp110-2、Os02g48110) 又与拟南芥AT4G16660聚为一枝(图2),既表明单子叶植物柳枝稷与水稻间亲缘关系比双子叶的拟南芥更近,更能揭示出起源于共同祖先的直系同源基因在单/双子叶植物分离前就已形成的进化特征[20]。此外,17 对旁系同源PvHsp70基因是两两分别位于柳枝稷K 和N 亚基因组中同源染色体上(图2、图3),在柳枝稷PvHsp20基因研究中也有相同结果[39],加之32 个PvHsp70基因存在共线性关系(图4),揭示出柳枝稷进化过程中的基因组复制与加倍,有助于对柳枝稷异源四倍体形成机制的深入研究。基因表达分析表明(图5),PvHsp70基因间在不同组织、发育阶段和镉处理下表达呈现差异。其中,PvHsp70-2和PvHsp70-10在各组织发育阶段的表达量都较高,可能在柳枝稷生长发育过程中具有重要调节作用。PvHsp70-4、PvHsp70-5、PvHsp70-7、PvHsp70-9、PvHsp70-13 、PvHsp70-18、PvHsp70-21、PvHsp70-22、PvHsp70-31和PvHsp70-32在特定生长发育阶段中表达量较高,暗示这些基因可能功能上有差异。

图5 柳枝稷PvHsp70 基因在21 个组织的表达量热图Figure 5 Heatmap of expression of Hsp70 genes of switchgrass in 21 tissues
研究表明,Hsp70在适应各类植物胁迫中发挥着重要的作用[40],可提高植物对逆境的抵御能力,如高温和干旱[41-42]、高盐[43]和重金属[44]。41 个PvHsp70基因中有29 个在镉胁迫下上调表达(图6),马铃薯在4 种非生物胁迫(盐、干旱、冷和热) 时,20 个StHsp70基因共有14 个基因全部处理或部分处理时上调,都反映出Hsp70基因能够响应非生物胁迫,调控植物的耐受能力[22]。植物遭受非生物胁迫(如高温、干旱、盐害等)时,植物细胞内会积累大量未折叠蛋白或错误折叠蛋白,造成内质网胁迫,此时细胞会启动蛋白监控信号通路-未折叠蛋白应答响应(UPR)[45-46]。分子伴侣BiP 是响应UPR 的重要成员,能帮助蛋白质正确折叠或迁移,从而起到稳定蛋白质的功能[5]。PvHsp70-15是镉胁迫下唯一持续上调表达的PvHsp70基因,其编码BiP 蛋白已被证实具有提升拟南芥耐镉的能力[34]。油菜(Brassica campestris)BiP同源基因BnA7Hsp70超表达后使油菜在缺水、渗透胁迫以及衣霉素处理后表现较强的耐受性[47]。由此可见,PvHsp70-15 在响应胁迫时可能提升柳枝稷逆境耐受能力,柳枝稷Hsp70家族中有4 个BiP基因(PvHsp70-1、PvHsp70-2、PvHsp70-15、PvHsp70-17) (表2),深入开展其逆境功能研究将对挖掘柳枝稷耐逆基因资源具有重要意义。
4 结论
本研究在柳枝稷基因组中共鉴定了41 个PvHsp70基因,分属分子伴侣Dank 亚家族(33 个)和HSP110/SSE 亚家族(8 个)。41 个基因不均衡分布在柳枝稷12 条染色体上,位于09K 染色体上分布数量最多(11 个)。基因共线性研究发现,32 个基因有共线性关系且1 对是串联复制基因。所有PvHsp70基因均为组成型表达,大部分具有组织特异性,参与柳枝稷生长与发育的调控。PvHsp70-15为分子伴侣BiP 编码基因,镉胁迫下持续上调表达,可作为耐逆基因深入开展功能研究。
