遮阴对粗肋草生长、光合特性和养分含量的影响
2022-11-18许珊珊钟明慧李玲燕伍丽华林开敏曹光球叶义全
许珊珊,唐 银,钟明慧,李玲燕,伍丽华,林开敏,曹光球,叶义全
(1. 福建农林大学林学院, 福建 福州 350002;2. 国家林草局杉木工程技术研究中心, 福建 福州 350002;3. 林木逆境生理生态及分子生物学福建省高校重点实验室, 福建 福州 350002)
光是影响植物生长的重要环境因子,它不仅与植物光合碳同化过程密切相关,而且还可作为重要的信号参与植物的生长发育与形态建成[1]。在长期适应不同光环境过程中,植物进化出适应不同光环境的能力以及在不同光环境下植物所特有的光能利用特性[2]。例如,杉木(Cunninghamia lanceolata)等喜光树种,它们对光强的要求相对较高,在过度遮阴条件下,由于无法获得足够的光能,导致光合作用受抑,影响植株生长[3-4]。人参(Panax ginseng)和三七(Panax notoginseng)等喜阴植物在高光强条件下,则可能由于高光强诱导活性氧的积累,破坏光反应机构,抑制植物光合作用,最终抑制植物生长[5-6]。在不同光环境中,植物主要通过叶片形态特征、叶色和光合生理特性等的改变,调控体内光合产物的合成、运输和分配等过程,增强自身对变化光环境的适应[7-8]。因此,筛选适合不同类型植物生长的光照条件是苗木培育的关键环节。
粗肋草(Aglaonema commutatun)“彩蝶”属于天南星科(Araceae)粗肋草多年生观叶草本植物,其叶片呈花色,叶面有斑点,富有光泽,叶片多呈卵形或卵状披针形,是一种重要的园林观赏植物,具有较高的观赏价值[9]。近年来,随着人们生活水平的不断提高,市场对观赏植物的需求量日益增长,对色彩鲜艳的不同粗肋草品种需求也随之增长。植物叶色的形成除了受遗传控制外,研究表明,环境因子对植物叶色的形成具有非常重要的调控作用[10]。光强作为重要的环境因子,不但与叶色转变过程紧密相关,而且对叶色的鲜艳程度具有重要影响[11]。因此,如何通过光环境的调控更好地培育粗肋草,满足市场对大量优质粗肋草苗木的需求已成为亟待解决的技术需求。目前关于粗肋草的研究主要集中在组培快繁[12-14]、叶片解剖结构特征[15]以及抗寒性[16]等方面,而有关粗肋草“彩蝶”品种的研究鲜见报道,特别有关“彩蝶”的生长、光合特性以及养分元素积累对不同遮阴处理响应的研究更是未见报道。鉴于此,本研究以3 月龄粗肋草“彩蝶”为试验材料,比较不同光照强度条件下“彩蝶”生长、生物量、光合特性和养分元素积累的差异,揭示“彩蝶”幼苗对不同光强的响应规律,以期为“彩蝶”合理光强的选择及其园林观赏植物的栽培提供理论依据。
1 材料和方法
1.1 试验材料与试验设计
本试验于2019 年在福建农林大学田间试验室大棚内进行(119°27′ E, 26°9′ N),海拔10 m。“彩蝶”幼苗为福建省鑫闽种业有限公司提供的优良无性系植株,于2019 年3 月中旬将组培苗经1 个月时间的驯化成活的,且生长健壮、长势一致的植株移栽 入 装 有 泥 炭 土 ∶ 珍 珠 岩 ∶ 粗 沙 = 3 ∶ 2 ∶ 1 (体 积比)基质的花盆(下底直径8 cm,盆口直径11.8 cm,高9.7 cm)中培养2 个月,进行常规的田间栽培管理。培养基质中的主要化学元素含量为:全氮13.41 g·kg-1,水解氮0.38 g·kg-1,全磷1.48 g·kg-1,有效磷0.65 g·kg-1,全钾20.12 g·kg-1,速效钾8.76 g·kg-1,镁6.37 g·kg-1,钙11.48 g·kg-1,锰0.47 g·kg-1,铁22.13 g·kg-1,铜0.12 g·kg-1。2019 年5 月中旬选取长势一致的“彩蝶”植株,选用不同规格遮阳网进行遮阴处理(试验处理排列如图1 所示),遮阳网设置好后用手持海博特光谱仪测定不同光照强度处理下光强,确保每个处理达到试验的遮阴程度要求。试验共设置3 个处理:不遮阴(CK)、遮阴率25% (T1)和遮阴率50% (T2),每个处理3 个重复,每个重复10 株,共计90 株幼苗。不同处理水肥管理一致,即每隔半个月叶片喷施1 次0.1%水溶性复合肥,水分管理主要通过喷灌设施,每隔1 d 喷10 min,培养两个月后调查不同处理下“彩蝶”的生长情况,根据调查结果从每个重复中选取3 株作为标准株,并将3 株混合取样当成1 个重复进行测定,每个处理共3 个重复。

图1 试验处理布设情况Figure 1 Layout of shading treatments
1.2 测定项目与方法
1.2.1 叶绿素荧光参数测定
叶绿素荧光方法参照叶义全等[17]的方法进行。即在遮阴处理结束后,选取不同处理相同位置成熟叶片利用PAM-2 500 便携式叶绿素荧光仪,分析不同光强处理下“彩蝶”植株成熟叶片的叶绿素荧光参数。测定前首先将叶片放入暗盒,进行30 min 的暗适应,接着用弱光测得初始荧光(Fo),随后给叶片饱和脉冲光测得最大荧光(Fm),在光适应10 min 后,打开饱和脉冲光测得光下最大荧光(Fm′),其余叶绿素荧光指标通过相关公式计算得到。
1.2.2 叶片光合色素含量测定
处理结束后从不同遮阴处理“彩蝶”的标准植株中选取成熟的叶片,在测定完光合特性后,将叶片取下进行光合色素含量分析。将叶片擦拭干净后,用剪刀除掉叶片主脉,随后将余下部分用剪刀剪碎混匀。使用电子天平称取0.3 g 的碎片,加入95%乙醇进行研磨提取,随后进行离心过滤,去除沉淀取上清定容至25 mL。利用分光光度计分别测定664、647 和466 mm 处吸光值。随后根据Arnon[18]公式计算出叶绿素a、叶绿素b 和类胡萝卜素含量。
1.2.3 光响应曲线测定
光响应曲线测定法参照蔡建国等[19]方法。选择晴天利用LI-6400 便携式光合仪(Li-Cor, Lincolin,USA)进行光响应曲线测定。进行光合特性测定时选取每株标准株上相同位置的成熟叶片进行分析。CO2浓度设为400 μmol·mol-1,叶室温度和相对湿度分别控制在(25 ± 0.5) ℃和(50% ± 5%),利用内置程序将光合有效辐射强度设置为1 200、1 000、800、600、400、200、100、50、20、0 μmol·(m2·s)-1。选取相同位置成熟叶片作为测定叶,首先将叶片置于1 000 μmol·(m2·s)-1的光强下进行光诱导15 min,随后利用光响应自动测量程序进行测量,每个光强梯度下测定3 min。光响应曲线拟合采用直角双曲线修正模型[20]:Pn(I) = α(1 -βI)/(1 +γI)I-Rd。式中:Pn为净光合速率,I为光合有效辐射,α 为初始量子效率,β和γ为系数,Rd为暗呼吸速率。根据模型计算表观量子效率(apparent quantum yield, AQY)、最大净光合速率(maximum net photosynthetic rate,Pnmax)、光饱和点(light saturation point, LSP)和光补偿点(light compensation point, LCP)等指标。
1.2.4 根系形态测定
根系形态扫描参考洪凯等[21]方法。试验结束后取出不同处理下标准株,将根系取出,用蒸馏水清洗根部后用吸水纸吸干,接着利用加拿大数字化根系扫描仪(STD1600 Epson)对不同处理下“彩蝶”根系进行扫描,最后采用WinRhizo 根系分析软件对总根长、根系总表面积、侧根数量和根系总体积进行定量分析。
1.2.5 营养元素含量测定
试验处理结束后,取出不同遮阴处理的“彩蝶”标准株幼苗,将根和地上部分分别收获后用纯净水将叶和根洗净,再用吸水纸吸干水分,分别称重,随后将根和叶分开装入信封,在105 ℃下烘箱杀青30 min 后,再在75 ℃下烘干至恒重。粉碎后用硝酸-高氯酸的方法消煮。采用电感耦合等离子体发射光谱仪(Perkin ICP-MS)测定植株根和叶中养分元素含量。
1.2.6 数据处理
试验数据采用DPS 8.5 软件进行方差分析(ANVOA),平均值间差异显著性采用LSD 法进行检验(P< 0.05),并用Origin 8.5 软件作图,图中数据以平均值 ± 标准差表示。
2 结果与分析
2.1 遮阴对粗肋草“彩蝶”生长表型和生物量的影响
不同遮阴程度下 “彩蝶”的生长表型存在较大差异(图2)。其中,T1处理下的“彩蝶”植株生长最好,叶色鲜艳,根系发达;进一步提高遮阴程度到50%(T2处理) 则明显抑制“彩蝶”地上部分和根系的生长,而且叶色不如T1鲜艳,有些泛黄;CK 处理下“彩蝶”根系和地上部植株的生长均明显受抑,叶片卷曲,叶片边缘和中部部分区域出现日灼现象导致坏死斑点。与表型结果类似,生物量结果(图2)表明,“彩蝶”的地上部分和根系生物量均表现为T1> T2>CK,尽管不同处理间地上部分生物量不存在显著差异(P> 0.05),但是T1处理下“彩蝶”根系生物量显著高于T2和CK 处理(P< 0.05) 。不同处理根冠比同样也表现为T1> T2> CK,其中,T1处理下根冠比显著高于T2和CK 处理(P< 0.05) (图2)。上述结果表明过高或者过低的光照强度均不适合“彩蝶”的生长,而适宜光照强度对“彩蝶”生长至关重要。
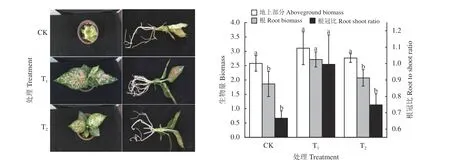
图2 遮阴对粗肋草“彩蝶”表型、生物量和根冠比的影响Figure 2 Phenotype, growth and root to shoot ratio of Aglaonema commutatum variety “Butterfly”seedlings grown under different shading treatments
2.2 遮阴对粗肋草“彩蝶”根系形态的影响
不同遮阴处理对“彩蝶”根系形态指标同样存在显著影响(表1)。 T1处理下“彩蝶”的根长显著高于T2和CK 处理(P< 0.05),分别较两者提高了96.31%和115.59%。不同遮阴处理下“彩蝶”根表面积大小表现为T1> T2> CK,其中T1处理下根表面积分别较T2和CK 处理增加18.64%和106.90%,且不同处理间根表面积大小均存在显著差异(P< 0.05)。根系总体积和平均根直径对不同遮阴处理的响应类似,两者均表现为T1> T2> CK,其中T1和T2处理下“彩蝶”根系总体积和根平均直径均显著大于CK(P< 0.05),两者分别较CK 提高了101.48%、77.78%和111.30%、78.53%。可见适度遮阴有利于促进“彩蝶”根系的生长。

表1 遮阴对粗肋草“彩蝶”根系形态的影响Table 1 Changes in root morphology of Aglaonema commutatum variety “Butterfly” seedlings grown under different shading treatments
2.3 遮阴对粗肋草“彩蝶”叶片光合色素含量的影响
与CK 相比,不同遮阴处理对光合色素的积累同样存在显著影响(表2)。不同遮阴处理下“彩蝶”叶片叶绿素a、叶绿素b 和类胡萝卜素的含量的变化均表现出类似规律,即随着遮阴程度的增加,叶绿素a、叶绿素b 和类胡萝卜素的含量均呈逐渐增加的趋势,T1和T2处理叶片叶绿素a、叶绿素b、类胡萝卜素含量均显著高于CK 处理(P< 0.05),分别较CK 提高了113.54%和187.51%、80.56%和149.31%、128.43%和241.07%。尽管随着遮阴程度的增加,叶绿素a/b 比值则呈“先升后降”的趋势,但不同处理之间叶绿素a/b 比值不存在显著差异(P> 0.05) 。

表2 遮阴对粗肋草“彩蝶”叶片光合色素含量的影响Table 2 Photosynthetic pigment contents in leaves of Aglaonema commutatum variety “Butterfly”seedlings grown under different shading treatments
2.4 遮阴对粗肋草“彩蝶”光合-光响应曲线的影响
3 种光照强度处理下“彩蝶”的光响应曲线变化趋势类似(图3),当光合有效辐射(photosynthetically active radiation, PAR)小于200 μmol·(m2·s)-1时,Pn随PAR 增加迅速增大,且T1处理的Pn增幅最大,随后Pn随PAR 增加的趋势放缓,当增加到一定程度时随着PAR 的增加Pn值表现出逐渐降低的趋势。在PAR 大 于200 μmol·(m2·s)-1时,相 同 光 强 条 件下Pn大小均表现为T1> T2> CK,说明T1处理下其净光合速率最高。此外,与CK 相比,遮阴处理提高了“彩蝶”叶片Pnmax和光饱和点值,降低了暗呼吸速率,其中T1处理Pnmax和光饱和点值分别较CK 提高了321.88% 和35.70%,而暗呼吸速率较CK 下降了55.04% (表3)。不同处理“彩蝶”表观量子效率和光补偿点的大小顺序分别为T1> CK > T2和T2>CK > T1。

表3 遮阴对粗肋草“彩蝶”叶片光合-光响应参数的影响Table 3 Photosynthesis-light response parameters in leaves of Aglaonema commutatum variety “Butterfly”seedlings grown under different shading treatments
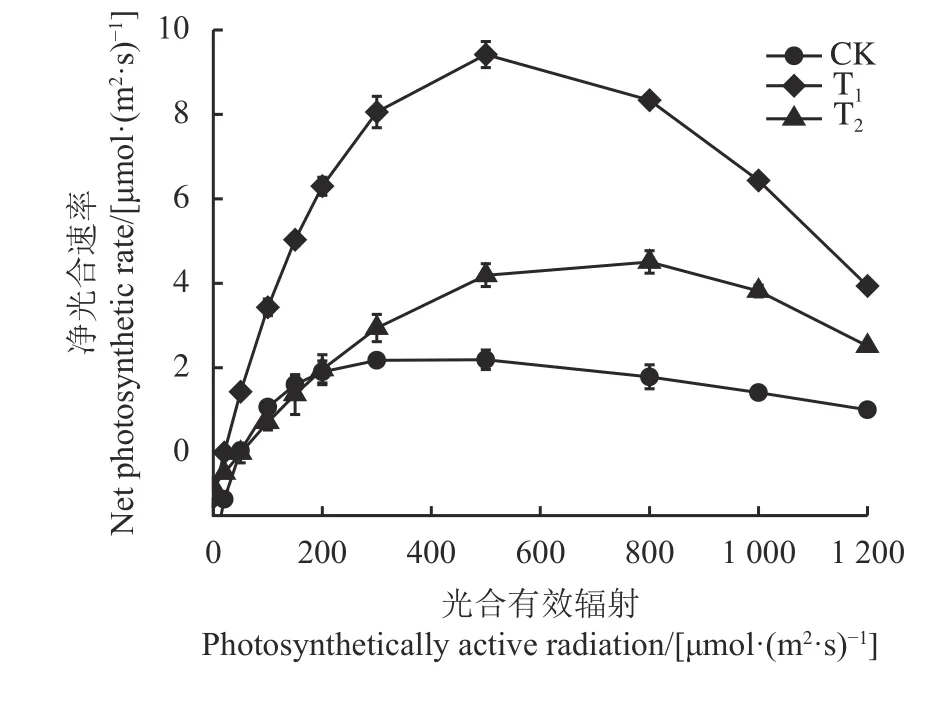
图3 遮阴对粗肋草“彩蝶”叶片光合-光响应曲线的影响Figure 3 Photosynthesis-light response curve in leaves of Aglaonema commutatum variety “Butterfly” seedlings grown under different shading treatments
2.5 遮阴对粗肋草“彩蝶”叶片叶绿素荧光参数的影响
随着遮阴程度的增加,Fo值逐渐下降,其中T1和T2处理下该值分别较CK 下降了22.76%与25.89%(表4)。与该变化趋势相反,随着遮阴程度的增加PSⅡ最大光化学效率(Fv/Fm)和非光化学猝灭系数(non-photochemical quenching coefficient, NPQ)值呈逐渐增大的趋势,T2处理下Fv/Fm、NPQ 值分别较比CK与T1处理提高了7.10%、2.75% 和42.06%、10.83%。此外,随着遮阴程度的增加,Fm、可变荧光(Fv)、PSⅡ潜在光化学活性(Fv/Fo)、光化学猝灭系数(qP)和PSⅡ实际光化学效率(QY) 值均呈“先升后降”的趋势,并T1处理下达到最大值,且T1处理下上述指标均显著大于CK (P< 0.05),分别较CK 提高了43.07%、49.45%、93.54%、23.15%和37.20%。

表4 遮阴对粗肋草“彩蝶”叶片叶绿素荧光参数的影响Table 4 The changes of chlorophyll fluorescence parameters in leaves of A. commutatum variety “Butterfly” under different shading treatments
2.6 遮阴对粗肋草“彩蝶”植株养分元素含量的影响
与CK 相比,遮阴抑制了“彩蝶”对磷的吸收,显著降低植株地上部分和根中的磷含量(P< 0.05) (表5)。其中,T1、T2处理下地上部分和根中磷含量分别较CK 下降了22.65%、27.93% 和23.26%、18.64%。不同遮阴处理降低“彩蝶”根中镁和钾元素含量,其中T1处理下这两种元素含量显著低于CK (P< 0.05),但显著促进这两种元素在地上部分的积累,T1处理下这两种元素含量显著高于CK,暗示T1处理可能通过促进这两种元素向地上部转运,改变其在植株体内的分布情况。与钾和镁变化趋势相反的是T1处理下“彩蝶”根中钙含量显著高于CK 和T2处理(P< 0.05),而其地上部分钙含量则显著低于CK 和T2处理(P< 0.05),暗示适度遮阴可能通过抑制钙离子向地上部转运,促进其在根中积累。

表5 遮阴对粗肋草“彩蝶”根和地上部分养分元素含量的影响Table 5 Nutrient element contents in roots and aboveground of A. commutatum variety“Butterfly” seedlings under different shading treatments
3 讨论
3.1 遮阴对“彩蝶”生长的影响
光与植物生长和形态建成密切相关,不同的光照强度可通过对植物体内碳同化产物合成、运输、分配和积累等一系列过程的影响,进而对植物的生长产生调控作用[22-25]。因此,在苗木培育过程中选择适宜的光照强度对幼苗的生长至关重要。本研究结果表明不同遮阴处理下“彩蝶”生长表型存在较大差异,25%遮阴(T1)处理下“彩蝶”生长最好,叶片色泽鲜艳,生物量明显高于50%遮阴(T2)和不遮阴(CK)处理,说明适宜的光强有利于“彩蝶”光合碳同化产物积累,促进植物生长,而过高或者过低的光强均不利于其生长。过高的光强可能由于叶片无法及时耗散过剩光能,引起叶片内活性氧大量积累,导致光合系统受损,进而抑制植物生长[26],而过低的光强由于植物无法获得足够的光能进行光合作用,无法为植株正常生长提供足够的物质和能力,从而导致其生长受抑。此外,本研究结果还表明,适度遮阴同样有利于促进“彩蝶”根系生长,提高根冠比,这可能是它适应特定光环境的一种生长策略,当植物所处光环境改变后,影响其光合碳同化产物分配,通过向根部转运更多的碳同化产物促进根系生长,来维持植物自身在适宜光环境下快速生长过程中对养分和水分的大量需求。综上所述,在本研究条件下25%的遮阴是比较适合“彩蝶”生长的光强条件。此外,由于本研究光强梯度设置不够详细,要想得到更为准确的适合“彩蝶”生长的光强条件,需要在今后试验中进一步细化光强梯度,进行研究。
3.2 遮阴对“彩蝶”光合生理的影响
叶绿素是植物进行光能捕获和传递等过程的重要成分,其含量的高低与植物光合能力密切相关[27]。研究表明,遮阴能不同程度提高叶绿素a 和b 以及总叶绿素含量,增强植物对光能的吸收能力,从而使植物在低光强下能捕获更多的光能进行光合作用,提高光合效率。本研究发现遮阴处理叶片叶绿素含量显著高于CK,说明在遮阴条件下,“彩蝶”可通过提高叶片叶绿素含量来增强植株光合能力。这与百山祖冷杉(Abies beshanzuensis)[28]和多花黄精(Polygonatum cyrtonema)[29]等的结果类似,这些结果共同表明增加叶片叶绿素含量可能是不同植物种类适应遮阴环境的共性机制,通过提高遮阴条件下叶片叶绿素含量来增强植株光合能力。除叶绿素含量外,叶绿素a/b 值也是表征植物对光环境适应的一个重要指标,它常用于衡量植物叶片光能利用能力的高低[30]。一般而言,在低光强条件下叶绿素a/b 值下降有利于促进叶片对蓝紫光的吸收,从而弥补因光强减弱导致的光能捕获量的下降[31]。本研究中,尽管不同处理间叶绿素a/b 值差异不显著,但遮阴处理下该值却高于对照,这与前人的一些研究结果不一致。尽管导致这种现象的确切机制尚不清楚,但部分原因可能由于“彩蝶”对强光较为敏感,强光处理导致叶绿素降解,对叶色产生“漂白”作用,从而在一定程度上对叶绿素含量及其a/b 比值产生干扰。类胡萝卜素在植物体内具有双重功能,它既能参与对光能的捕获,同时还可作为一种重要的内源抗氧化剂,在清除植株体内活性氧中扮演着重要角色[32-33]。本研究表明遮阴处理显著增加叶片类胡萝卜素的含量,这更可能是“彩蝶”适应遮阴环境下增强光能吸收能力的一种策略,从而使叶片在低光照环境条件能捕获更多的光能用于光合作用。
植物光响应曲线是了解植物光合能力的一种有效方法[34]。与对照相比,遮阴处理“彩蝶”叶片最大净光合速率显著增大,而暗呼吸速率则显著下降,表明弱光条件下“彩蝶”具有较强的弱光利用能力,它一方面通过增强光合作用能力,另一方面通过降低暗呼吸速率来共同促进光合碳同化产物积累,维持其在低光强条件下的碳平衡,满足自身正常生长对能量的需求[35-36]。表观量子效率主要用于表征植物叶片在弱光环境下的光合能力,其值大小与叶片中负责吸收与转换光能的色素蛋白复合体呈显著正相关,该值越大表明色素蛋白复合体越多,对弱光的利用能力就越强[37]。本研究中,与对照相比,T1处理下表观量子效率显著升高,而T2处理下该值显著降低,说明“彩蝶”对弱光具有一定的适应性,但过度遮阴则会显著降低其对光能的利用能力。光补偿点和光饱和点分别反映了植物对弱光和强光的利用能力以及植物的需光范围[34]。通常而言,光饱和点低且光补偿点低的植物具有较强的耐阴性[38]。与CK 相比,尽管在T1处理下“彩蝶”光补偿点显著下降,但在T2处理中其光补偿点高于CK 处理,而且遮阴处理下光饱和点均显著高于对照,表明“彩蝶”不是耐阴性较强的植物,但具有一定的耐阴性。一般而言,耐阴植物的光补偿点小于20 μmol·(m2·s)-1[39],而“彩蝶”的光补偿点介于21.68~59.36 μmol·(m2·s)-1,因 此 从 这 一 点 上 也 印 证 了“彩蝶”不是耐阴性强的植物。该结果与生长和表型结果共同说明对于耐阴能力不强的“彩蝶”而言,由于其在过度遮阴的环境下光能利用效率较低,因此过度遮阴会抑制光合作用,减少碳同化产物的积累,进而在一定程度上抑制其生长。而适当的遮阴,则会通过提高表观量子效率、光饱和点,降低光补偿点和暗呼吸速率来适应弱光环境,促进碳同化产物的积累,表现出良好的生长态势。
叶绿素荧光技术主要用于衡量叶片光系统对光能的吸收、传递、耗散和分配的特征,与光合作用关系密切[40]。Fo是反映PSⅡ反应中心基本状态的关键指标,该中心发生不可逆破坏以及可逆失活都会导致该值的升高[41]。与遮阴处理相比,CK 处理Fo值显著提高,暗示全光照处理可能导致叶片中的PSⅡ反应中出现不可逆破坏或可逆失活。Fv/Fo主要用于表示活性的PSⅡ反应中心的数量,而Fv与Fm则用于分别表征该反应中心的活性大小和电子传递情况[42]。本研究中,遮阴处理上述指标均高于CK,说明适度遮阴可以增加反应中心数量,提高电子传递效率,进而增强叶片对光能的利用效率。Fv/Fm衡量PSⅡ反应中心原初光能转换效率的指标,正常情况下该值通常较稳定,维持在0.80~0.85,而在胁迫条件下会引起该值出现不同程度的下降[43]。与CK 相比,遮阴处理不同程度提高Fv/Fm值,且CK处理下该值低于0.80,说明在全光照条件下植株受到一定程度的光胁迫,发生光抑制现象。NPQ 是一种光合机构的自我保护机制,植物通过将吸收的多余光能以热能形式耗散,防止过剩光能对光合机构造成破坏[41]。遮阴处理下NPQ 值均高于CK,暗示在全光照条件下,由于叶片无法及时耗散过剩光能,导致其自身光合结构造成破坏,这可能也是导致其叶片光合能力受抑的一个原因。可见,适度的遮阴有利于减轻强光对“彩蝶”PSⅡ反应中心的损害,提高叶片PSⅡ反应中心的光化学效率和电子传递速率,进而增强植物碳同化的能力,促进植株生长。
3.3 遮阴对“彩蝶”养分元素含量的影响
本研究发现,CK 处理下“彩蝶”植株茎与根中的磷含量均显著高于遮阴处理,引起这种现象的部分原因可能由于强光照条件显著抑制“彩蝶”的生长,造成该处理下植株生物量较小,从而产生一定的“浓缩”效应,但确切的原因有待进一步研究。值得注意的是,与CK 和T2处理相比,在本研究中最适宜“彩蝶”生长的T1处理中,尽管植株根中镁和钾含量明显下降,但地上部这两种元素含量显著增加,暗示遮阴处理可能促进了这两种元素向地上部转运。遮阴促进这两种元素向地上部转移的确切机制尚不清楚,但这两种元素在遮阴条件下向地上部转运增强具有非常重要的生理学功能,因为它们与植物的光合作用密切相关。钾元素可参与调控叶片气孔开度,进而影响胞间二氧化碳浓度,同时它也参与光合电子传递链,因此缺钾会导致叶片发黄,显著降低叶片光合能力[44-45]。类似地,镁元素作为叶绿素核心成分,在植物光合作用中扮演关键角色。缺镁能显著降低叶绿素含量,降低植物光合速率,同时缺镁还会抑制光合碳同化产物向根部转运,导致其在叶片中积累,从而进一步抑制叶片的光合能力[46-47]。因此,适度遮阴下促进钾和镁离子向地上部转运,对于促进叶绿素合成提高“彩蝶”叶片光合能力具有重要意义,该结果与上述光合结果相一致。此外,与钾和镁元素结果相反的是,适度遮阴(T1)处理显著抑制钙离子向地上部转运,促进其在根中显著积累。作为细胞壁的重要组成部分之一,钙离子在调控根细胞壁延展以及根系发育中扮演关键角色,它通过与果胶羧基上的残基交联形成钙桥凝胶网络,赋予细胞果胶一定的结构刚性,进而调控根系生长[48],而缺钙会导致细胞壁刚性增强,无法正常伸长,进而影响根系的发育[49]。因此适度遮阴(T2)处理下植株根系生长显著优于其他处理,部分原因可能是由于该处理促进钙离子向根部转运,为根系生长发育提供充足的钙离子。
综上所述,本研究结果显示25%遮阴是粗肋草“彩蝶”最适宜生长的光照条件,其潜在的调控机制是:1)适度的遮阴通过增加叶片光合色素含量,增强叶片对光能的传递和利用效率,提高光合效率,进而增强叶片碳同化能力,增加光合产物积累。2)适度的遮阴可以通过促进根系的发育,增强根系对钙、镁和钾元素的吸收和合理分配,从而最终共同促进“彩蝶”的生长。
