喜马拉雅旱獭骨骼肌低氧适应的组织学特点
2022-11-17秦鸿楠张勤文
秦鸿楠,周 娟,李 琳,张勤文
(青海大学农牧学院,青海 西宁 810016)
喜马拉雅旱獭(Himalayan marmot)属啮齿动物,主要分布在我国西藏、甘肃、青海等地区。栖息于海拔2 500~5 200 m的高山草甸草原、高山草原等环境中[1-2]。旱獭为了适应这种高原低氧环境,其形态学和生理学都发生了一定的适应性变化。研究发现,处于不同海拔高度的喜马拉雅旱獭,其心肌肌纤维都出现不同程度的紊乱和肌节模糊的现象[3-4]。马嵋等[5]研究表明,新疆灰旱獭右心室肥厚指数、纤维化程度均高于SD大鼠。喜马拉雅旱獭的红细胞(Red blood cell,RBC)、红细胞比容(Hematocrit value,HCT)和血红蛋白含量随海拔增加逐渐增加,从而提高氧气的利用率[4]。贾荣莉[6]研究发现,喜马拉雅旱獭具有肺泡中毛细血管丰富、支气管上皮存在杯状细胞等特点。
骨骼肌是哺乳动物躯体运动的物质基础。对于肌肉组织学特性的研究表明,可以根据肌纤维直径和肌纤维表面积密度来判定肌肉品质,肌纤维直径越小,肉质越好;影响肌纤维粗细的因素有很多,如品种、营养状况和年龄等[7-8]。在有关高原牦牛的研究中发现,与平原黄牛相比,大通牦牛骨骼肌肌纤维直径细,而表面积密度大[9]。研究发现,随着海拔的升高,大通牦牛骨骼肌肌纤维直径随着年龄的增长逐渐增大,肌纤维表面积密度也随之增加[10]。叶于聪等[11]研究表明,急进高原大鼠的心肌细胞的超微结构发生变化,线粒体变形肿胀,基质密度下降,嵴出现不同程度的溶解或缺损等;因低氧引起急进大鼠心肌线粒体变形,使心肌细胞内腺嘌呤核苷三磷酸(Adenosine triphosphate,ATP)合成减少,能量代谢受到影响,从而引起细胞内外水肿甚至造成心肌细胞坏死,而旱獭心肌细胞超微结构正常,表明喜马拉雅旱獭对高原低氧环境有良好的适应性。目前对于旱獭骨骼肌低氧适应的组织学特点报道较少,因此本试验以喜马拉雅旱獭为研究对象,以SD大鼠为对照,采用苏木精-伊红(Hematoxylin-eosin,H.E.)染色、免疫组织化学和电镜技术,对旱獭骨骼肌的组织学特点进行观测和分析,探讨旱獭骨骼肌组织低氧适应的组织学特点,旨在阐明旱獭骨骼肌的组织学特点和对高原低氧环境适应的特点,进一步为高原医学研究提供理论依据,并为高原兽医研究积累资料。
1 材料与方法
1.1 主要试剂 兔抗鼠血管内皮生长因子(Vascular endothelial growth factor,VEGF)多克隆抗体、CD34多克隆抗体、柠檬酸盐缓冲液和DAB显色剂,均购自武汉赛维尔生物科技有限公司;胎牛血清,购自北京科瑞泰科技有限公司。
1.2 主要仪器 脱水机(Donatello),购自DIAPATH公司;包埋机(JB-P5)和冻台(JB-L5),均购自武汉俊杰电子有限公司;病理切片机(RM2016)和超薄切片机(LKB-2188),均购自上海徕卡仪器有限公司;组织摊片机(KD-P),购自浙江金华市科迪仪器设备有限公司;烤箱(DHG-9140A),购自上海慧泰仪器制造有限公司;脱色摇床(TSY-B)、涡旋混合器(MX-F)和组化笔(WG1066-1),均购自武汉赛维尔生物科技有限公司;正置荧光显微镜(Nikon Eclipse C1)、透射电镜(JEM-2000EX)和成像系统(Nikon DS-U3),均购自日本尼康公司。
1.3 动物和处理 选取青海省地方病预防控制所饲养的成年喜马拉雅旱獭5只和SD大鼠5只,临床健康,不计性别。喜马拉雅旱獭、SD大鼠均于现场屠宰后立即取其腓肠肌,一部分用4%多聚甲醛固定,用于H.E.染色和免疫组织化学染色;一部分用2.5%戊二醛固定,用于电镜指标的测定。
1.4 肌纤维直径和肌纤维表面积密度测量 常规石蜡切片,H.E.染色。用Image-Pro Plus 5.1图像分析系统直接测量肌纤维直径,并按照张勤文等[12]报道的方法测算出肌纤维表面积密度。
1.5 免疫组织化学染色 切片常规脱蜡至水;PBS冲洗;将切片放入柠檬酸盐缓冲液中修复抗原,冷却至室温;PBS冲洗,滴加3%H2O2消除内源性过氧化物酶活性并室温孵育;PBS冲洗,滴加5%的胎牛血清封闭液,不洗;分别滴加兔抗鼠VEGF和CD34多克隆抗体,4 ℃过夜;PBS冲洗,滴加生物素化山羊抗兔工作液(1∶200)稀释;PBS冲洗,滴加试剂SABC室温孵育;PBS冲洗后DAB显色剂显色,待有阳性细胞出现且背景淡时用蒸馏水终止反应;用酒精脱水,二甲苯透明,中性树胶封片,切片于荧光显微镜下观察并采集图像。VEGF阳性表达胞浆着色为棕黄色颗粒,用Image-Pro Plus 5.1图像分析系统直接测量VEGF在单位面积内的阳性表达量,按张勤文等[12]报道的方法测算出单位面积内肌纤维数量,以阳性表达量/单位面积内肌纤维数量,所得比值作为VEGF在单位面积内的阳性表达率。根据Weidner等[13]校正方法将染色的任一细胞或细胞丛作为一微血管,先在低倍镜下观察切片全部视野并确定微血管密度最高区域,后在高倍镜下计数5个视野的微血管数,取其平均值作为该样本的微血管密度(Microvascular density,MVD)。
1.6 电镜指标观测 将已处理好的样品制成70 nm超薄切片,切片用醋酸铀和枸橼酸铅进行双重染色。在JEM-2000EX透射电镜下观察,随机拍摄并在透射电镜放大10 000倍后,进行照片扫描入电脑。参照郑富盛[14]的相关计算方法,采用方格测量系统,测算出线粒体的基本参数。按立体学公式[15]计算出线粒体的平均截面积(Ax)、平均体积(V)、体积密度(Vv)、面数密度(NA)等各结构参数。
1.7 数据统计分析 通过SPSS 23.0软件对试验数据进行单因素方差分析,试验数据以平均值±标准误来表示。以P<0.05作为差异显著性的标准。
2 结果
2.1 喜马拉雅旱獭与SD大鼠骨骼肌肌纤维直径和肌纤维表面积密度比较 由表1和图1~2可知,喜马拉雅旱獭的肌纤维直径小于SD大鼠,且差异极显著(P<0.01),喜马拉雅旱獭的肌纤维表面积密度大于SD大鼠,且差异显著(P<0.05)。
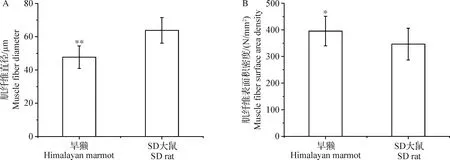
图1 喜马拉雅旱獭与SD大鼠骨骼肌肌纤维直径(A)和肌纤维表面积密度(B)的比较

表1 喜马拉雅旱獭与SD大鼠骨骼肌肌纤维直径和肌纤维表面积密度的比较
H.E.染色时细胞核呈蓝色,肌纤维呈粉红色。由图2可知,喜马拉雅旱獭骨骼肌肌纤维排列致密、规则且肌纤维较细,SD大鼠骨骼肌肌纤维排列较疏松。
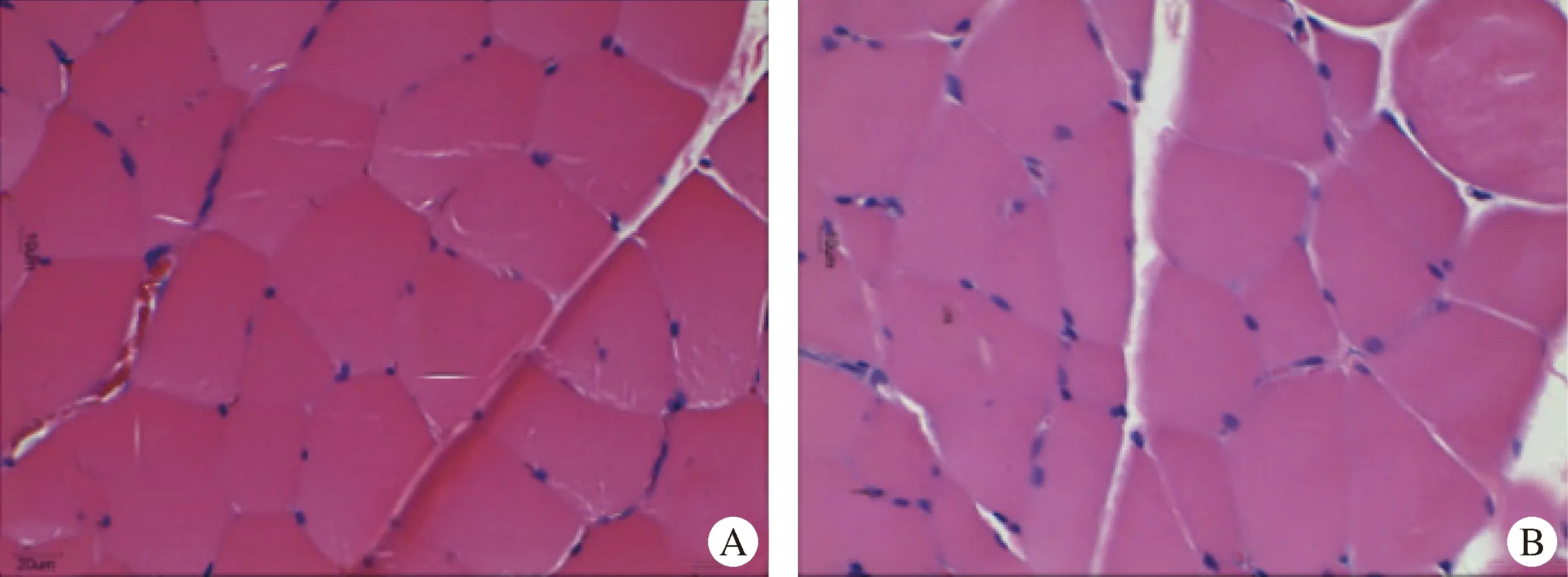
图2 喜马拉雅旱獭(A)和SD大鼠(B)骨骼肌的显微结构(H.E.染色,400×)
2.2 喜马拉雅旱獭与SD大鼠骨骼肌中VEGF和MVD比较 由表2和图3可知,喜马拉雅旱獭骨骼肌中VEGF阳性表达率和MVD均高于SD大鼠,且差异均极显著(P<0.01)。
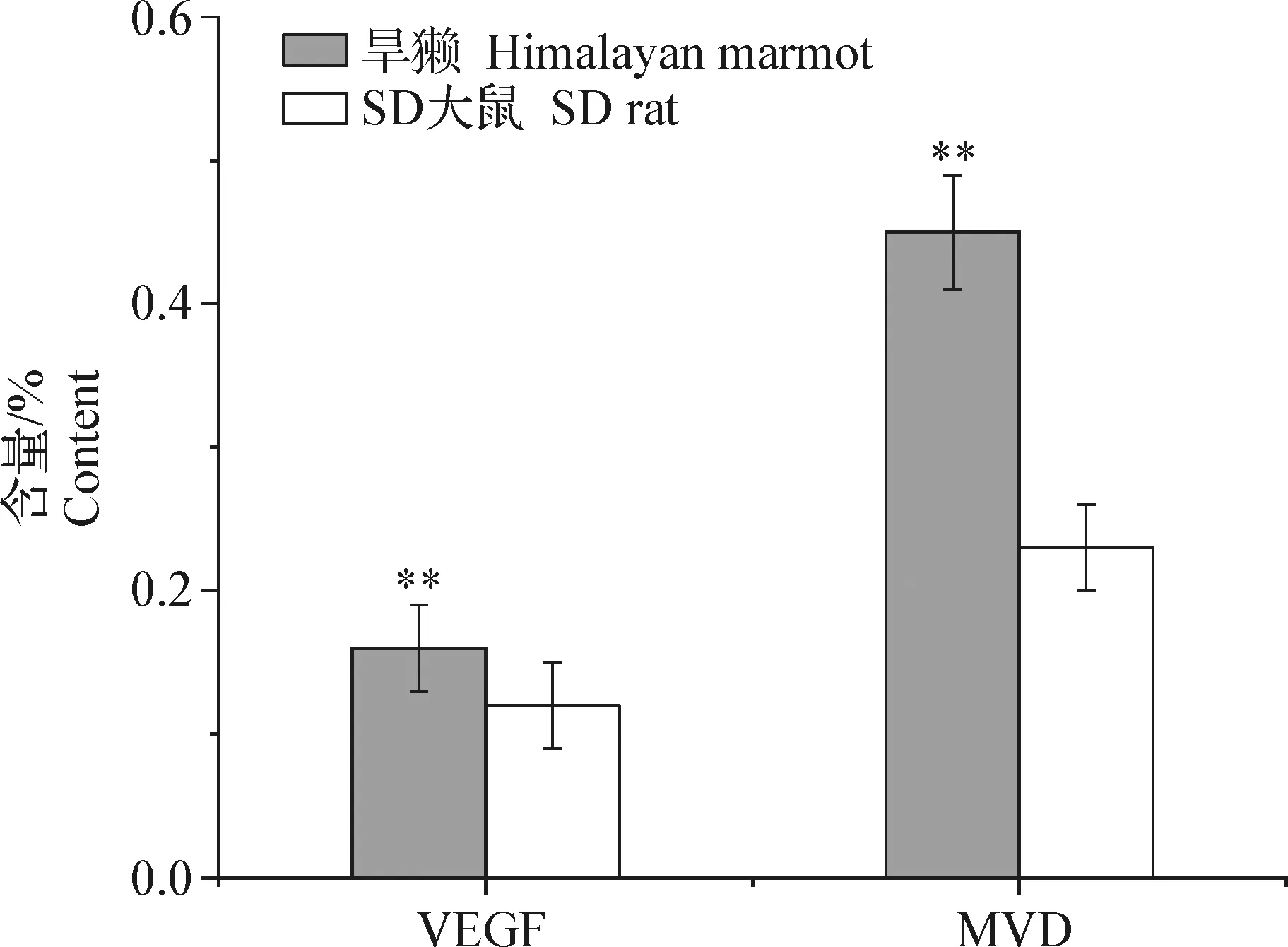
图3 喜马拉雅旱獭与SD大鼠骨骼肌中VEGF和MVD的比较

表2 喜马拉雅旱獭与SD大鼠骨骼肌中VEGF和MVD的比较
免疫组化染色中棕黄色即为阳性表达,通过棕黄色的深浅可判断VEGF的表达量,CD34标记的棕黄色为微血管内皮细胞。由图4可知,喜马拉雅旱獭VEGF、MVD阳性表达量均高于SD大鼠。

图4 喜马拉雅旱獭与SD大鼠骨骼肌的免疫组化染色(400×)
2.3 喜马拉雅旱獭与SD大鼠骨骼肌细胞线粒体的平均截面积、平均体积、体积密度和面数密度比较 由表3和图5可知,喜马拉雅旱獭骨骼肌线粒体平均截面积和平均体积均小于SD大鼠且差异极显著(P<0.01);喜马拉雅旱獭骨骼肌细胞线粒体的体积密度和面数密度均大于SD大鼠且差异极显著(P<0.01)。
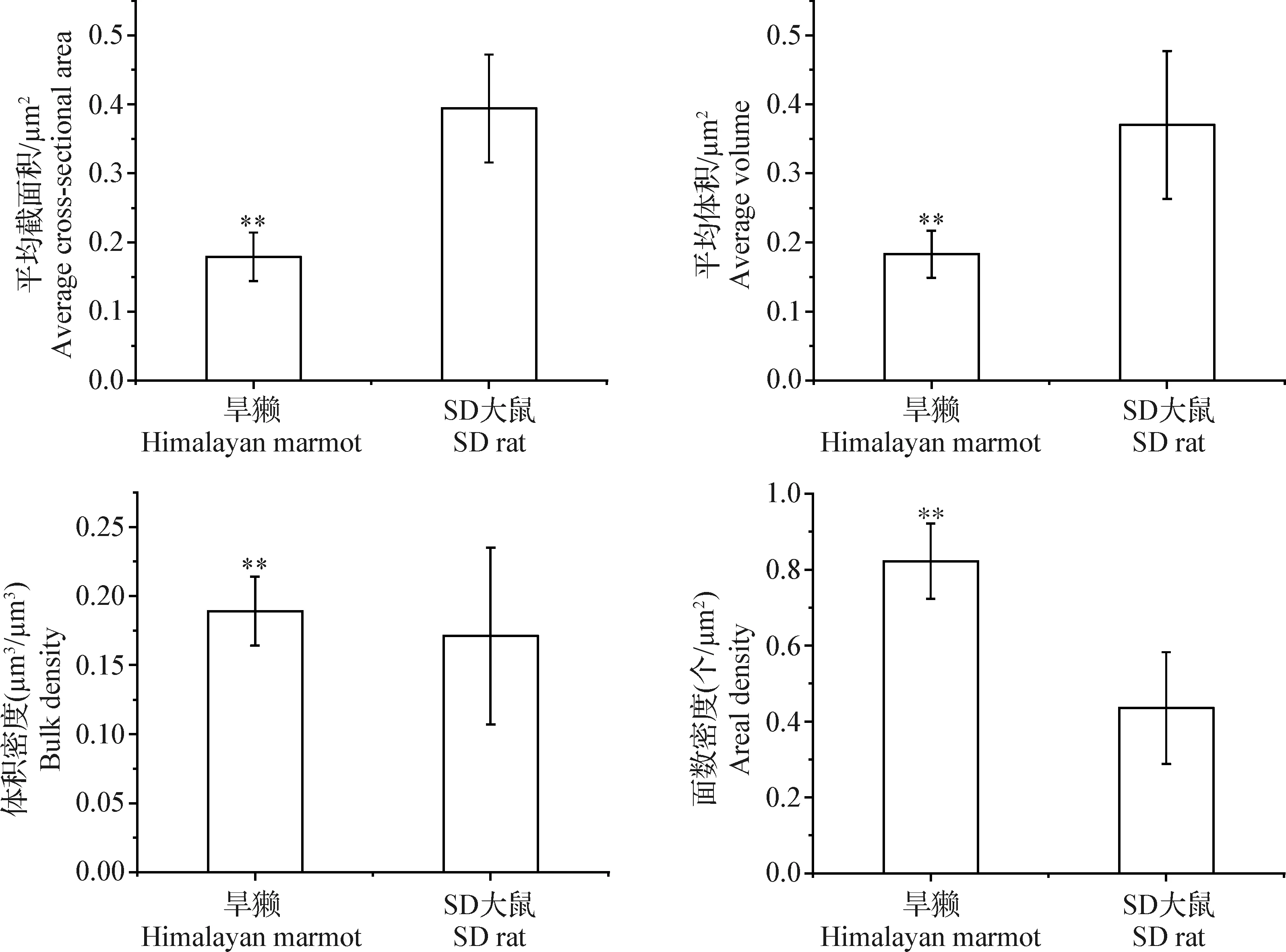
图5 喜马拉雅旱獭与SD大鼠骨骼肌细胞线粒体的平均截面积、平均体积、体积密度和面数密度的比较

表3 喜马拉雅旱獭与SD大鼠骨骼肌细胞线粒体的平均截面积、平均体积、体积密度和面数密度的比较
由图6可知,喜马拉雅旱獭与SD大鼠线粒体结构均完整规则,喜马拉雅旱獭线粒体排列紧凑,SD大鼠线粒体排列疏松;喜马拉雅旱獭骨骼肌线粒体表现出体积小、数量多的特点。

图6 喜马拉雅旱獭(A)与SD大鼠(B)骨骼肌细胞线粒体分布情况(10 000×)
3 讨论
骨骼肌在机体运动过程中有着很重要的作用,衡量骨骼肌组织学特性主要有2个重要指标:肌纤维直径和肌纤维表面积密度。有研究显示,在低氧环境下,牦牛的肌纤维直径较枣北黄牛细[9]。这与本试验中喜马拉雅旱獭肌纤维直径比SD大鼠细一致。肌纤维表面积密度是根据肌纤维数得到的,每个肌纤维就是1个肌细胞。低氧会导致动物骨骼肌受损,因此骨骼肌细胞通过不断增生和促进骨骼肌肌纤维生成来达到适应低氧的目的。有研究表明,在单纯缺氧时,骨骼肌细胞会发生增殖并在第21天达到最高峰,持续低氧28 d骨骼肌细胞增殖情况逐渐下降,这可能是骨骼肌开始适应低氧环境的表现[16]。在本试验中,喜马拉雅旱獭骨骼肌肌纤维表面积密度比SD大鼠大,这可能是因为喜马拉雅旱獭长期生活在高原低氧环境中,其骨骼肌肌纤维受到环境影响逐渐变细,从而使肌纤维表面积密度增大,以达到适应高原环境的目的。
在低氧条件下,骨骼肌毛细血管的增生可以缩短氧从毛细血管向组织弥散的距离,进一步提高了组织供氧量,是组织与细胞对低氧环境的适应性反应,引起血管增生的主要原因是VEGF的高表达,而血管增生会导致MVD发生变化[17]。VEGF的高表达与机体在缺氧时产生低氧诱导因子(Hypoxia-inducible factor,HIF)密切相关,其包括HIF-1α和HIF-1β,HIF-1α参与组织细胞缺氧的反应。在缺氧时大量的HIF-1α表达可导致VEGF反转录水平上升而大量表达[18]。朱世海等[19]研究发现,高原鼢鼠的MVD和VEGF显著高于SD大鼠,高原鼢鼠通过MVD和VEGF的高表达特征来适应高二氧化碳的洞道生活。本试验中喜马拉雅旱獭骨骼肌中VEGF阳性表达率和MVD比SD大鼠高,说明喜马拉雅旱獭对低氧环境已经具有良好的适应性。
线粒体是细胞生命活动的场所,也是细胞内氧化磷酸化和ATP形成的重要场所。细胞氧化磷酸化和ATP形成重要部位是线粒体的内膜与嵴。有研究表明,低氧时线粒体会发生变形肿胀,嵴出现不同程度的断裂损伤,从而减少氧化磷酸化和ATP形成[20]。线粒体形态的完整才能保证其正常功能,缺氧时机体通过增加线粒体的数目和面积从而加强对氧的利用。齐新章[21]研究表明,高原鼢鼠与高原鼠兔心肌组织对氧的摄取和利用能力较强,这与其心肌中线粒体面密度、面数密度大有关,心肌中线粒体通过增加线粒体数目和面数密度来提高机体对氧的摄取与利用。本试验中喜马拉雅旱獭骨骼肌线粒体的平均截面积和平均体积均小于SD大鼠;旱獭骨骼肌线粒体的体积密度和面数密度均大于SD大鼠。所以线粒体结构的特殊性可以使喜马拉雅旱獭更好地适应低氧环境。
综上所述,喜马拉雅旱獭骨骼肌显微结构的特殊性,是其适应高原低压低氧环境的组织学基础,喜马拉雅旱獭骨骼肌结构特点说明其对高原环境存在良好的适应性,主要表现为骨骼肌的肌纤维直径细,肌纤维表面积密度大,VEGF大量表达,MVD大,线粒体体积密度和面数密度高。
