青藏高原多年冻土区两种高寒草地生态系统土壤氮季节变化及其与环境因子的关系
2022-11-16谢梅珍吴晓东周华云岳广阳
谢梅珍, 赵 林, 吴晓东, 周华云, 岳广阳
(1.中国科学院西北生态环境资源研究院冰冻圈科学国家重点实验室藏北高原冰冻圈特殊环境与灾害国家野外科学观测研究站,甘肃 兰州 730000;2.中国科学院大学,北京 100049;3.南京信息工程大学地理科学学院,江苏 南京 210044)
0 引言
多年冻土为温度在0℃以下,存在两年或两年以上的岩土层[1]。我国的高纬度多年冻土主要分布在东北大、小兴安岭地区,高海拔多年冻土主要分布在西部高山及青藏高原地区,多年冻土总面积约2.15×106km2,占国土总面积的22.3%[2]。其中青藏高原地区多年冻土的实际面积为1.06×106km2[3]。在多年冻土区,活动层的冻融循环过程中,土壤会发生一系列的热量交换及水分迁移[4],这些变化会进一步影响土壤中养分的生物化学循环[5-9]。氮素在自然界中以不同形态的含氮分子或离子形式存在,通过矿化,硝化、反硝化以及固定过程在生态系统中循环,在维持生态平衡中占有非常重要的地位[10],在气候变化的大背景下,青藏高原土壤氮循环受到显著影响[11-12]。因此,了解青藏高原土壤中各形态氮素的动态变化特征以及与环境因子的关系对于揭示氮循环对气候变化的响应有重要的意义。
土壤可利用氮与微生物量氮组分比大分子有机氮更容易被转化或被微生物及植物吸收利用[13],这些氮组分的季节变化与植物生长有密不可分的关系,例如在青藏高原及北极地区,土壤可利用氮含量在植物生长旺盛期降低,在植物生长季后期或非生长季初期升高,且土壤微生物量氮含量在生长季也受到与植物之间养分竞争的限制[14-16]。除了植物吸收对土壤可利用氮与微生物量氮库的影响之外,土壤水热状况也是氮组分季节变化的重要驱动因子[17-18]。在多年冻土区,活动层的冻融循环分为夏季融化过程、秋季冻结过程、冬季降温过程及春季升温4个阶段[4],在活动层冻融循环过程中,土壤热量交换及水分迁移过程复杂,尤其在冻融交替季节[19-21]。另有研究表明,土壤可利用氮在冬春交替季/积雪融化季节有所升高[22-23],土壤微生物量亦会受到雪深的影响[24-25],这些现象对于氮淋失或减轻后续季节氮对植物生长的限制作用有非常重要的影响[26]。这些研究阐述了不同季节土壤可利用氮与微生物量氮的动态变化情况,但是,在青藏高原多年冻土区,整个冻融循环期或连续的生长季与非生长季中氮的季节变化与驱动因子的作用还不清楚。
青藏高原多年冻土区主要的草地生态系统有高寒沼泽草甸、高寒草甸、高寒草原及高寒荒漠,其中高寒草甸及高寒草原分别占有植被分布区域面积的48.59%和27.96%[27]。本研究通过对青藏高原高寒草甸及高寒草原2个不同典型草地生态系统下土壤可利用氮(铵态氮NH4+-N、硝态氮NO3--N及可溶性有机氮DON)及微生物量氮(MBN)的季节变化特征进行分析,揭示青藏高原高寒草甸及高寒草原草地生态系统下不同氮组分季节变化规律及其与环境因子之间的关系,为多年冻土区氮循环模式及其对气候变化的响应提供研究基础。
1 材料与方法
1.1 研究区概况
本研究位于青藏高原多年冻土区腹地,该地区具有典型的干旱高原气候,且分布有连续多年冻土。选择2个典型的高寒草地生态系统,包括风火山高寒草甸及特大桥高寒草原生态系统作为研究区(图1)。风火山高寒草甸及特大桥高寒草原研究区的海拔分别为4 896 m及4 563 m,年平均气温分别为-4.3℃及-4.0℃,活动层厚度分别为2.0 m及3.4 m,年均降水量分别为386 mm及350 mm,降水主要集中分布在夏季[16]。两个研究区年平均积雪天数小于15天[28]。多年冻土区活动层冻融循环分为4个阶段:夏季融化期、秋季冻结期、冬季降温期以及春季升温期。植物生长季为5月至9月,非生长季为10月至次年4月,风火山高寒草甸主要植物种类为高山嵩草(Kobresiapygmaea)、矮嵩草(Kobresiahumilis)等,总植被盖度及地上生物量分别为80%及138.5 g·m-2,特大桥高寒草原主要植物种类为青藏苔草(Carexmorrcrofti)、紫花针茅(Stipapurpurea)等,总植被盖度及地上生物量分别为23.3%及43.7 g·m-2。研究区土壤性质见表1。
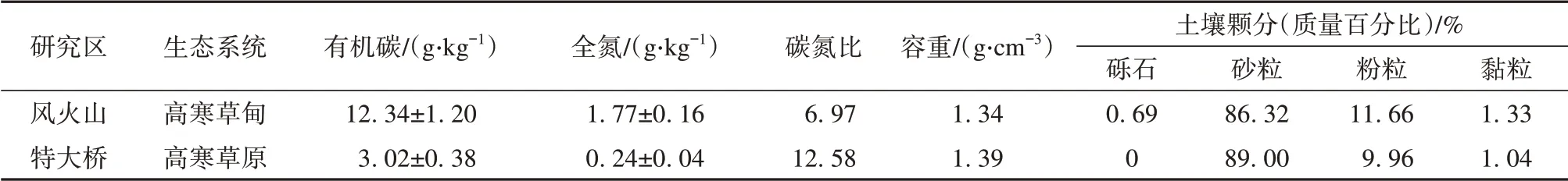
表1 研究区土壤性质Table 1 Soil properties of the study area

图1 研究区位置Fig.1 Location of the study area
1.2 样品采集与分析
从2016年4月至2017年3月,每个月初进行土壤样品采集,其中2016年12月及2017年2月未采样。植被调查在2016年8月初进行。本研究使用随机采样方法。在每个研究区中,选择平坦且植被均匀处随机设置3个5 m×5 m大小采样点,每个采样点间距至少5 m。采样时使用直径为3.5 cm的土钻进行采样,采样深度为0~10 cm。每个采样点中采集3个样品混合后形成1个混合土样,因此,每个研究区收集到9个原始土壤样品,并混合成为3个土样。将采集好的混合土样立即装入自封袋,储存在装有冰袋的保温箱中送回实验室。将样品过2 mm筛,移除砾石及植物凋落物后分成两份,一份储存到4℃冰箱中用于测定土壤NH4+-N、NO3--N、DON及MBN含量,一份在室内阴凉处风干,用于测定土壤全氮含量(TN)及pH值。具体方法如下:土壤可利用氮为NH4+-N、NO3--N、DON的总和,其中NH4+-N的测定利用2 mol·L-1KCl溶液提取后在间断分析仪(SmartchemTM200)上用苯酚钠比色法测得;NO3--N的测定利用2 mol·L-1KCl溶液提取后在间断分析仪(SmartchemTM200)上用硫酸肼比色法测得;DON采用土壤可溶性全氮与无机氮之间的差别计算得出;MBN的含量使用氯仿熏蒸后经过0.5 mol·L-1K2SO4溶液提取与高温消煮,与未熏蒸的差值得出;TN用凯氏定氮法测定(Foss8400);SOC采用重铬酸钾氧化法测定;pH值用酸度计(PHS-3C型酸度计)测定;土壤容重采用环刀法测定;土壤颗分采用具有不同直径筛孔的土筛测定。土壤10 cm处温度由Campbell Scientific Inc测定,观测精度为±0.03℃,土壤10 cm处体积含水量使用TDR-100测定,观测精度为±3%。
1.3 统计分析
采用Microsoft Excel 2003和SPSS 16.0统计分析软件进行数据处理及差异显著性检验;采用重复测量方差分析比较2种草地生态系统中土壤氮组分含量的差异,显著性水平设定为α=0.05;采用皮尔森相关系数表示土壤活性氮组分与环境因子(土壤全氮、土壤pH、土壤温度及土壤含水量)的相关性。采用Origin 8.0进行绘图。
2 结果
2.1 土壤性质
土壤温度在高寒草甸中变化幅度较高寒草原略小。在2个研究区中,土壤温度均为7月最高,1月最低。土壤水分在高寒草甸中高于高寒草原。在2个研究区中,土壤水分均在生长季初期升高,在生长季保持全年内的较高水平,随后在11月下降(图2)。

图2 土壤温度与水分状况Fig.2 Soil temperature and soil volumetric water content conditions
高寒草甸中土壤TN在生长季初期略有下降,生长季结束后逐渐升高;高寒草原中土壤TN含量在全年内保持稳定。高寒草甸及高寒草原土壤TN的年内平均含量分别为1.85 g·kg-1及0.25 g·kg-1(图3)。

图3 土壤全氮(TN)含量的年内变化Fig.3 Monthly variation of soil TN content
2.2 土壤可利用氮及微生物量氮的季节变化
在2个研究区中,土壤NH4+-N的变化趋势相似,且年内变化显著。在活动层冻融交替季,高寒草甸土壤NH4+-N含量较高,高寒草原含量较低。在生长季初期,2种草地生态系统土壤NH4+-N急剧下降,在6月至10月之间保持稳定,在11月又快速上升。总体上看,土壤NH4+-N在非生长季含量高于生长季。高寒草甸及高寒草原中土壤NH4+-N年内平均含量分别为7.57 mg·kg-1及4.13 mg·kg-1(图4)。

图4 土壤铵态氮(NH4+-N)含量的年内变化Fig.4 Monthly variation of soil NH4+-N content
土壤NO3--N在2个研究区中变化趋势相似,在生长季初期呈升高趋势,在生长季末期及非生长季初期呈降低趋势。在非生长季中期与深度冻结期,土壤NO3--N含量升高,随后在冻融交替季节下降。高寒草甸及高寒草原土壤NO3--N年内平均含量分别为8.45 mg·kg-1及6.28 mg·kg-1(图5)。
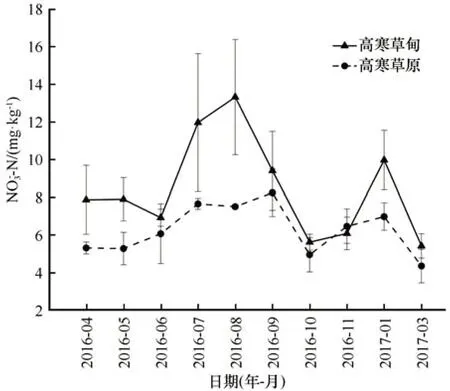
图5 土壤硝态氮(NO3--N)含量的年内变化Fig.5 Monthly variation of soil NO3--N content
土壤DON在2个研究区的变化趋势有所不同,在高寒草甸地区,土壤DON含量在5月呈上升趋势,随后下降保持稳定,直至1月再次上升;在高寒草原中,土壤DON含量在生长季逐渐下降,在非生长季逐渐上升,次年3月又有所下降。高寒草甸及高寒草原土壤DON年内平均含量分别为16.29 mg·kg-1及8.79 mg·kg-1(图6)。

图6 土壤可溶性有机氮(DON)含量的年内变化Fig.6 Monthly variation of soil DON content
土壤MBN含量在高寒草甸中变化剧烈,在高寒草原中变化趋势较平缓。在高寒草甸中,土壤MBN含量从4月至6月急剧上升,随后急剧下降,在非生长季初期和冻结初期又有所上升。在高寒草原中,土壤MBN含量在5月有所上升,随后下降,在生长季中期至非生长季初期期间再次上升,随后在非生长季缓慢下降。在高寒草甸及高寒草原中土壤MBN年内平均含量分别为41.96 mg·kg-1及5.10 mg·kg-1(图7)。
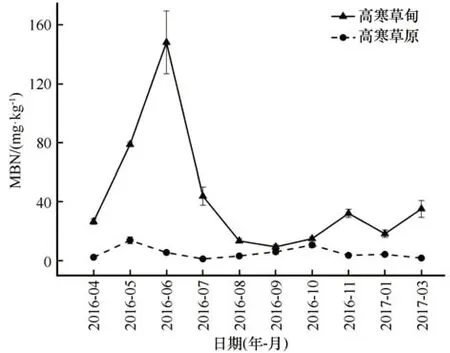
图7 土壤微生物量氮(MBN)含量的年内变化Fig.7 Monthly variation of soil MBN content
2.3 土壤可利用氮、微生物量氮与环境因子之间的关系
土壤NH4+-N与MBN在不同采样时间及草地生态系统与采样时间的交互作用下均具有显著差异,土壤NO3--N与DON在不同草地生态系统之间具有显著差异,在不同采样时间及其与草地生态系统的交互作用下差异不显著(表2)。
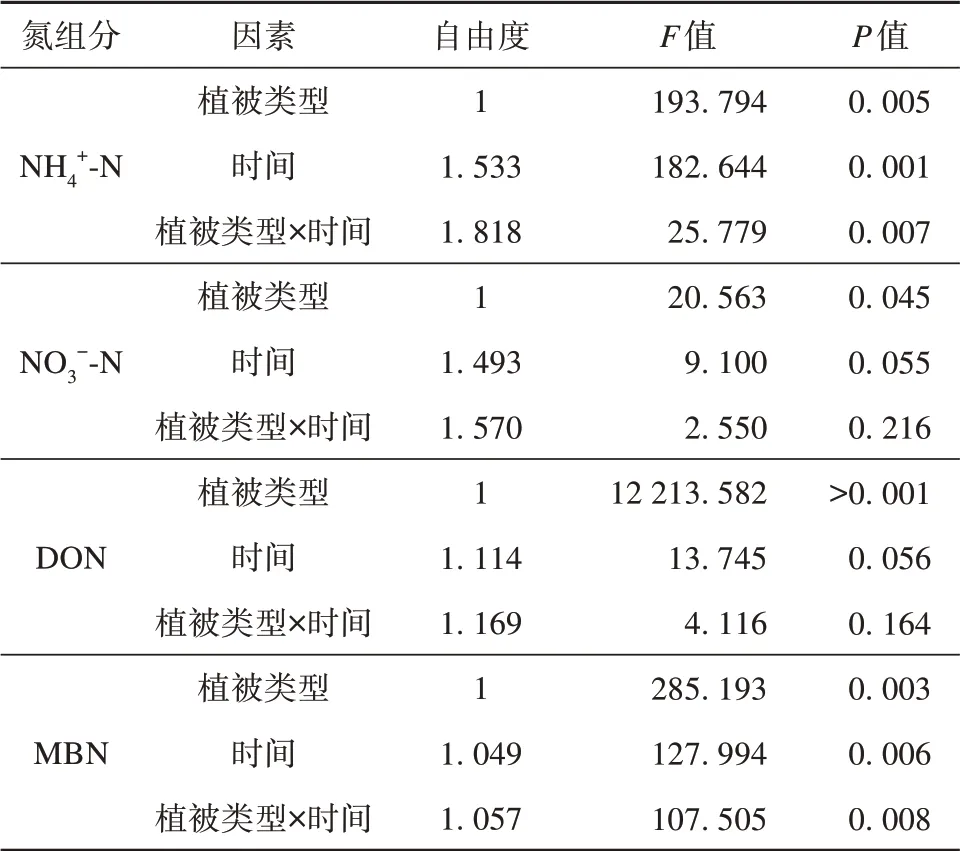
表2 土壤氮组分两因素重复测量方差分析Table 2 Results from two-way repeated-measures ANOVA analysis of soil nitrogen fractions
在高寒草甸区,土壤可利用氮与含水量之间的相关系数为-0.98,呈显著负相关,土壤MBN与TN之间的相关系数为0.69,呈显著正相关;在高寒草原区,土壤可利用氮与TN之间的相关系数为0.78,呈显著正相关(表3)。

表3 土壤可利用氮、微生物量氮与环境因子之间的皮尔森相关系数Table 3 Pearson’s correlation coefficients among soil available nitrogen,MBN and environmental factors
3 讨论
3.1 土壤可利用氮及微生物量氮的季节变化
土壤可利用氮各组分的含量在不同的草地生态系统中季节变化趋势相似,说明在研究区域内,不同草地生态系统对土壤中各组分可利用氮的季节变化趋势的影响较小。影响土壤可利用氮季节变化的主要因素有微生物分解转化,植物与微生物的吸收利用以及淋失等因素[16]。本研究中2个研究区域均位于青藏高原腹地,具有类似的气候条件与植被生长时期,可能导致了在不同草地生态系统中土壤可利用氮组分具有相似的季节变化趋势。
在整个采样期中,土壤NH4+-N在生长季较低,而NO3--N在生长季较高。硝化作用与反硝化作用在调节土壤NH4+-N与NO3--N的平衡中起到重要作用[29-30],研究表明在一定范围内土壤水分含量升高有利于增强硝化作用[31]。在青藏高原风火山高寒草甸及特大桥高寒草原地区,表层土壤含水量达不到使土壤处在厌氧环境下的含量,因此表层土壤中主要以硝化作用为主。青藏高原降雨主要发生在夏季,此时土壤含水较高,可促进硝化作用的发生,使土壤中NO3--N含量升高,且生长季中较强的氨氧化作用使土壤NH4+-N含量保持在较低的水平[30]。本研究中,高寒草甸地区土壤NH4+-N含量在冻融交替季节(4月)高于高寒草原,Freppaz等[32]、李源等[33]认为,冻融可能导致土壤中不可利用的有机胶体中NH4+-N的释放,且冻融作用可破坏土壤有机质结构使其更易分解为小分子物质[34]。在本研究中,高寒草甸土壤有机质含量显著高于高寒草原,因此在高寒草甸地区,土壤受冻融作用的影响,可能释放出更多的NH4+-N,导致在高寒草甸土壤中NH4+-N含量高于高原草原。土壤DON可被微生物矿化分解为无机氮,或被植物直接吸收利用[35-36],在氮地球化学循环及生态系统的稳定方面占有很重要的作用[37]。本研究中,土壤DON在生长季有所下降,这可能与生长季土壤DON被植物吸收以及此时期淋溶损失有较大关系。一般来说,土壤团聚体呈负电性,因此同为负电性的DON不易被吸附,导致其易在土壤含水量较高的生长季随水分迁移淋失,含量下降。
土壤微生物是土壤评价的重要指标[10],土壤MBN可表征土壤中微生物的活性[38-39]。本研究中,在生长季,土壤MBN在2种草地生态系统中均呈下降趋势。在高原上植物更倾向于吸收NO3--N,微生物更倾向于吸收NH4+-N[40]。本研究发现,土壤中NH4+-N含量在生长季较低,这可能导致了土壤MBN含量在生长季减少。冻融作用对土壤微生物的影响受环境条件的制约,一方面冻融交替可导致微生物死亡分解[41],使土壤中现有微生物活性下降[32];另一方面死亡的微生物残体中释放的养分可供幸存微生物使用,可激发残留微生物活性[42]。本研究中,土壤融化过程中(4月)微生物氮含量较低,而冻结过程中(10月)土壤微生物量氮含量略有上升。这可能是由于在融化期,冻融作用导致大量微生物死亡,而在冻结期土壤水分变化较小,为幸存微生物提供了较为稳定的生存环境,使土壤中残留微生物的活性缓慢增强[42]。在10月至3月,高寒草甸中土壤MBN含量呈波动上升趋势,而高寒草原中呈缓慢下降趋势。研究表明,土壤温度、含水量及其相互作用可导致不同草地类型土壤MBN含量变化趋势不同[15]。高寒草原地区土壤含水量小于高寒草甸地区,冬季土壤冻结可使高寒草原地区土壤未冻水含量进一步降低,进而影响土壤中微生物活性[15]。此外,土壤MBN在2种不同草地生态系统中变化趋势相似,而波动幅度差异较大。由于高寒草甸土壤有机质含量丰富,土壤中具有更多的可供分解周转的底物,而在高寒草原中,土壤有机质含量较低,微生物仅可利用有限的底物,使得高寒草甸土壤MBN变化幅度大于高寒高原[43]。
3.2 土壤可利用氮、微生物量氮与环境因子的关系
土壤可利用氮与MBN的季节变化受到环境因子的驱动[15,24]。本研究中,土壤温度与土壤水分对土壤可利用氮与MBN的影响不明显,甚至在高寒草甸中土壤水分与可利用氮呈现负相关现象,这可能是由于本研究中土壤水分主要受降水的影响[44]。降水主要集中在夏季[45],此时植被对土壤氮养分的吸收较强烈,减弱了土壤水分对土壤可利用氮与MBN的作用效果。在不同的草地生态系统中土壤可利用氮、土壤MBN与环境因子之间关系存在差异,其中土壤TN在2种草地生态系统中分别与MBN及可利用氮呈正相关。土壤TN可提供被微生物分解利用的底物,因此土壤TN含量是直接限制土壤可利用氮与MBN含量的因素之一。此外,尽管土壤pH值与可利用氮及MBN在本研究中无显著相关性,但有研究表明,土壤pH值可以影响土壤微生物群落结构以及酶活性[38],可能会进一步影响土壤MBN。
多年冻土地区具有气候条件独特,生态系统较敏感的特点,这些地区氮循环过程更容易受到气候变化的影响。目前,在青藏高原对于氮循环各个环节与环境因子之间的关系还缺乏长期规律的野外观测数据支持。因此,继续在长时间序列上对青藏高原土壤氮进行监测,对加深氮循环的认识具有重要的科学意义。
4 结论
本研究分析了青藏高原地区2种典型的草地生态系统中土壤可利用氮及MBN的季节变化,结果表明,2种草地生态系统中,土壤可利用氮组分与MBN均存在明显的季节波动。土壤可利用氮及MBN在2种不同草地生态系统中季节变化趋势相似,风火山地区高寒草甸生态系统中土壤MBN变化幅度大于特大桥地区高寒草原生态系统,此外,不同的可利用氮组分变化趋势不同。研究区中不同草地生态系统对土壤氮组分季节变化趋势影响较小,而风火山地区高寒草甸生态系统土壤中较高的土壤有机质含量使土壤MBN具有较大的变化空间。土壤TN可影响土壤氮组分含量的季节变化,但在不同的草地生态系统中其影响模式不同。
