氯仿熏蒸对紫色土农田土壤有机碳矿化的影响*
2022-11-16王小国
马 晗,王小国
(1.中国科学院山地表生过程与生态调控重点实验室/中国科学院水利部成都山地灾害与环境研究所 成都 610041;2.中国科学院大学 北京 100049)
土壤是陆地生态系统中最大的有机碳库,占全球陆地总碳库的2/3~3/4,超过陆地植被和全球大气碳库之和[1],其微小变化可能引起大气CO2浓度的较大波动[2]。土壤有机碳(SOC)矿化是指土壤中的有机碳在土壤微生物的作用下分解并向大气释放CO2的过程[3],受环境(温度,降水)、人为(施肥、耕作)及生物因素等的影响。普遍认为,矿化受土壤微生物的生物量大小、群落组成和活性影响[4-5]。全面理解SOC 的动态及其关键控制过程,有助于正确评估SOC 的变化方向和速率,对准确预测气候变化及制定应对气候变化的策略和措施具有重要意义[6-7]。
氯仿具有极强的杀灭微生物的特性,熏蒸后土壤理化性质基本保持不变[8],被广泛应用于土壤微生物生物量碳的测定。24 h 的熏蒸会杀死土壤中80%~90%的微生物,并破坏大多数细胞代谢过程,虽仍有部分酶介导的代谢过程在继续进行[9],但氧化酶基本上完全失活[10];熏蒸后土壤微生物多样性及种群数量会大幅下降[11],Jenkinson 等[12]表示即使进行再培养,53 d 后土壤微生物种类及数量仍不足未熏蒸土壤的20%。Kemmitt 等[13]及Van Elsas 等[14]的研究表明,通过熏蒸杀死土壤中的微生物,SOC 的矿化速率仍与未熏蒸土壤一致;1990年Joergense 等[15]的研究表明,即使没有新鲜有机质的加入,土壤仍可以在很长一段时间以非常恒定的速率矿化土壤有机质。针对以上现象,Kemmitt 等[13]提出“调节门”假说,认为:土壤有机质(SOM)的矿化与微生物生物量大小、群落结构或活性无关;矿化过程分为非生物过程(K1)与生物过程(K2),非生物有效性腐殖质通过K1转变为生物有效性有机质,在K2 中微生物对这部分生物有效性有机质进行缓慢矿化;矿化速率限制步骤受K1 控制。大量研究表明熏蒸后SOC 的矿化与未熏蒸土壤一致[12-17],但多数研究者仍对“调节门”假说提出质疑:Zhou 等[16-17]通过彻底熏蒸后接种培养及反复熏蒸后培养观测土壤中微生物情况及SOC 的矿化情况对“调节门”假说进行了验证,但该研究未提及不同群落结构对土壤有机碳矿化的影响,无法完整解释“调节门”假说中群落结构对SOC 矿化无关的结论。本研究通过改变熏蒸时长调整土壤微生物生物量,不仅改变了土壤微生物生物量,还极大改变了土壤微生物群落组成,以进一步验证“调节门”假说。
紫色土作为长江上游极为重要的耕地资源,集中分布在四川省与重庆市(三峡库区),仅四川盆地面积达16 万km2。紫色土地区气候湿润,年降雨量大多在800~1200 mm,降雨分布不均,集中在夏季。紫色土农田SOM 含量普遍偏低,平均约为1.20%[18]。根据中国科学院盐亭紫色土农业生态站多年观测数据发现,常规耕作下的旱地SOM 含量呈现下降趋势,其下降的原因主要与温暖湿润的亚热带环境条件下物质循环强烈有关。为探究土壤微生物生物量是否是紫色土SOC 矿化的主要限速因子,我们开展了SOC 矿化的室内培养试验,通过调整熏蒸时长改变土壤微生物的生物量大小来研究土壤微生物生物量对SOC 矿化速率及累积通量的影响。
1 材料与方法
1.1 研究区概况
研究样地位于四川省绵阳市盐亭县林山乡中国科学院盐亭紫色土农业生态试验站(105°27′E,31°16′N),林山乡地处四川盆地中部偏北,丘陵地带,属亚热带湿润季风气候区,雨热同期,年均降水量826 mm,年均气温17.3 ℃,无霜期294 d,平均海拔400~600 m。土壤属于碱性蓬莱镇石灰性紫色土,质地为中壤,土壤母质为泥岩。种植制度为冬小麦(Triticum aestivum)-夏玉米(Zea mays)轮作,耕层深度0~15 cm。
1.2 试验处理与样品分析
1.2.1 试验处理
2021年5月25日,小麦收获后选取长期施用猪粪肥处理的试验小区随机取15 个样点,去除地表残茬,挖取0~15 cm 土层土样带回实验室,过2 mm 筛,充分混合均匀并用镊子仔细除去动植物残体、石砾等杂质。土样分成3 份,一份新鲜土样保存在4 ℃条件下用于室内培养试验;一份新鲜土样用于测定土壤微生物生物量碳及熏蒸时长与微生物含量变化的关系;第三份土壤样品风干用于测定土壤总有机碳、全氮、pH 与颗粒组成等基础指标。土壤有机碳、全氮与全磷含量分别为(8.18±1.01)g·kg—1、(0.86±0.01)g·kg—1与(0.69±0.03)g·kg—1,土壤pH 为8.1,土壤微生物量碳含量为(184.50±28.54)mg·kg—1,土壤砂粒、粉粒、黏粒含量分别为46.9%、30.0%和23.1%。
1)试验1:熏蒸时长与土壤微生物生物量的关系
不同土壤微生物对氯仿的敏感性不同,随熏蒸时长增加,死亡微生物数量会逐渐增加[19-20]。一般利用24 h 暗培养后土壤总有机碳(TOC)增量换算的土壤微生物生物量碳来表示土壤微生物生物量[19]。目前极少有研究明确指出熏蒸时长与紫色土土壤微生物生物量变化的关系。本试验通过逐小时增加熏蒸后暗培养时长,计算熏蒸前后土壤可浸出有机碳的差值,将24 h 熏蒸后得出的微生物量视为1,通过与熏蒸24 h 土壤TOC 的比值确定土壤微生物生物量的变化情况[20],当微生物生物量变化幅度较大时,用二分法加测,确定更具体的时间(假设3 h 测得微生物生物量为90%,4 h 测得微生物生物量为20%,则增加3.5 h 熏蒸处理)。
2)试验2:培养试验
4 ℃保存的鲜土样调节含水量为田间持水量的60%[19],于25 ℃培养箱中活化土壤微生物。活化后称取相当于干土重10 g 的鲜土,进行熏蒸。选择试验1 中土壤微生物生物量变化较大的3 个熏蒸时长、24 h 熏蒸及未熏蒸5 个处理,具体熏蒸时长及编号分别为熏蒸24 h(C24)、熏蒸2.5 h(C2.5)、熏蒸1.5 h(C1.5)、熏蒸1 h(C1)与不熏蒸对照(CK),每个处理重复3 次。熏蒸后土壤转移到250 mL 培养瓶中于25 ℃培养箱培养,培养期间每周通过恒重法补充水分,保持土壤水分始终维持在田间持水量的60%。
1.2.2 气体样品采集与测定
培养试验共进行53 d,分别在培养1 d、2 d、4 d、7 d、11 d、18 d、25 d、32 d、46 d、53 d 时采集气体样品。每次采样前,利用抽气泵抽气30 min,使培养瓶内气体与大气保持一致。随后密封培养2 h,分别采集密封时的空气与密封2 h 后瓶内气体样品,用气相色谱仪(GC,Agilent 7890A/B,美国)测定CO2浓度,计算有机碳矿化释放CO2速率。
基础土样的SOC 与全氮采用元素分析仪测定,全磷采用钼锑抗比色法测定,pH 采用电位法测定,颗粒的机械组成采用吸管法测定。
1.3 数据计算与统计分析
1.3.1 微生物生物量碳计算
1)土壤微生物生物量碳
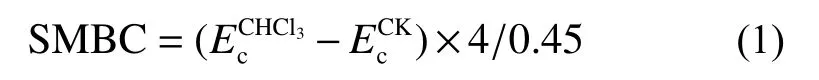
2)剩余存活微生物占比

式中:A为土壤中剩余存活微生物在土壤总微生物中所占百分比(%),SMBC24h熏蒸为熏蒸24 h 后土壤微生物生物量碳含量(mg·kg—1),SMBCih熏蒸为熏蒸ih处理后土壤微生物生物量碳含量(mg·kg—1)。
1.3.2 有机碳矿化释放CO2-C 速率、累积通量
1)有机碳矿化释放CO2-C 速率计算公式:

式中:F为CO2-C 释放速率(μg·g—1·h—1),k为转换常数1.796 g·m—3,dc/dt为在观测时间内气体浓度随时间变化的直线斜率(μg·h—1),V为培养容器的体积(m3),m为土壤干质量(g)。
2)有机碳矿化释放CO2-C 积累通量计算公式:

式中:C i为土壤CO2-C 释放积累通量(μg·g—1),F i和F i-1分别为相邻两次测定的土壤呼吸CO2-C 释放速率,T i为两次测定的时间间隔。
1.3.3 统计分析
采用Excel 2016 软件和Origin 2018 软件对数据进行处理和绘图,采用SPSS 22 软件进行数据分析,包括Pearson 相关分析、单因素协方差分析及重复测量方差分析(P<0.05)。
2 结果与分析
2.1 土壤微生物生物量随熏蒸时长的变化
土壤存活微生物量随熏蒸时长的增加呈下降趋势,熏蒸后,土壤微生物快速死亡,6 h 熏蒸时长可以杀死90%以上微生物(图1)。随熏蒸时长增加,单位质量土壤可浸提微生物生物量碳呈上升趋势,但是略有波动。根据图1,选取土壤微生物生物量变化幅度较大的3 个时间点,杀死土壤中74.53%微生物的2.5 h、52.68%的1.5 h 与杀死15.71%的1 h,再添加24 h 完全熏蒸及完全不熏蒸的空白对照进行培养试验。

图1 土壤微生物生物量随熏蒸时长的变化Fig.1 Changes of soil microbial biomass with the length of fumigation time
2.2 不同熏蒸处理对土壤有机碳CO2 排放速率的影响
通过观察培养期间CO2排放速率的变化情况(图2),将其分为3 个阶段:1~11 d,大幅波动期;11~32 d,平稳期;32~53 d,缓慢下降期。培养期间,不同熏蒸处理土壤CO2排放速率整体变化趋势相同。除C2.5 处理增长幅度最大外,其余处理表现出CO2排放速率最大值随熏蒸时长增加而增加。与CK 处理相比,C24、C2.5、C1.5、C1 处理最大排放速率分别增长182.00%、309.01%、73.85%和30.45%。

图2 不同熏蒸时长下的土壤有机碳矿化Fig.2 Mineralization of different fumigation durations on soil organic carbon
培养当天,熏蒸处理CO2初始排放速率显著高于CK(P<0.05)。培养第2 天各处理CO2排放速率间差异不显著。第2 天后,C2.5 与C24 处理土壤CO2排放速率快速增加,并在第7 天达最高点,且C2.5 处理的CO2排放速率在前7 d 培养中始终高于C24;C1.5 与C1 处理较C2.5 与C24 处理峰值出现时间更早,在第4 天达到最高点。在培养第11 天,各处理间CO2排放速率未见显著差异,且回落至与培养初期相当的水平。此后,各处理间CO2排放速率存在差异但不显著。相比于CK 处理,熏蒸处理32 d 后的CO2排放速率有所降低,但差异不显著。
通过表1 可以发现,整个培养期间内熏蒸处理CO2排放速率均值显著高于CK(P<0.05),除C2.5 处理外,其余处理CO2排放速率均值随熏蒸时长增加而增加,培养7~53 d 内各处理间CO2排放速率均值则无显著性差异。重复测量方差分析表明与CK 相比,熏蒸显著促进土壤CO2排放速率增长,事后检验发现:CO2排放速率C24 和C2.5 处理在第4 天与第7天显著高于其他时间,C1.5 处理在第4 天显著高于其他时间,所有熏蒸处理在第53 天显著低于其他时间(P<0.05)。

表1 不同熏蒸时长下土壤CO2 排放速率均值Table 1 Average CO2 emission rates of soil under different fumigation durationsμg·g—1·h—1
2.3 不同熏蒸处理对土壤有机碳矿化的影响
观察整个培养周期内的累积CO2排放通量,C24与C2.5 处理显著高于CK(P<0.05),其余处理间差异不显著(图3)。相比于CK 处理,C24、C2.5、C1.5 与C1 涨幅分别为22.37%、37.29%、6.76%和—1.45%。

图3 不同熏蒸时长下土壤CO2 累积排放量Fig.3 Soil accumulative CO2 emission under different fumigation durations
熏蒸处理在7~53 d 的累积CO2排放通量上没有表现出显著的促进或抑制作用(图3)。相比于CK,C24 处理增长—4.56%,C2.5 处理增长—5.38%,C1.5 处理增长—0.31%,C1 处理增长—7.30%。对7 d 后的土壤累积CO2释放通量进行拟合分析,累积通量变化数据符合模型:

式中:y为累积CO2排放通量,t为累积培养天数,a、b为常数。熏蒸与未熏蒸处理累积排放通量的拟合方程之间存在显著差异(P<0.05)。
从表2 与图2 可以看出不同处理系数a的大小与排放速率峰值大小相反,系数b的大小与排放速率峰值大小一致。系数a越大表明累积通量初始增量越大,系数b越接近零,表明随培养时间t的增加,累积通量的增加越平缓。所有拟合方程的b值均<1,表明随培养时间的延长,累积通量的增加越来越平缓。其中C2.5 处理增量降低的最快,CK 累积通量的增量降低速率最小,P值均小于0.01。

表2 不同熏蒸时长下土壤累积CO2 排放通量(y)与累积培养天数(t)的拟合方程Table 2 Fitting equations of accumulative CO2 release of soil(y)with incubation days(t)under different fumigation durations
3 讨论
3.1 土壤微生物生物量随熏蒸时长的变化
氯仿熏蒸可以有效破坏土壤微生物的细胞膜结构,促进其中的活性组分(C、N、P 等)释放到土壤中[8],随熏蒸时长的增加,土壤中可浸提微生物生物量碳逐渐增加。本试验结果表明,土壤中可浸提微生物生物量碳随熏蒸时长的增加呈现波动增加的变化趋势。刘雨晴等[22]测定了不同熏蒸时长处理下土壤微生物生物量碳的变化情况,结果表明18 h 熏蒸时长条件下,高有机质土壤与低有机质土壤的微生物生物量有机碳含量均略低于12 h 熏蒸时长,针对此现象,认为土壤微生物细胞膜损坏后,土壤中增加的活性碳组分部分被分解,造成可浸提微生物生物量碳含量的降低。因此,熏蒸处理一旦结束应当立即进行浸提,并测定计算土壤中的微生物生物量有机碳,以减少该部分碳的矿化损失,提高结果的准确度。
3.2 熏蒸时长对土壤有机碳矿化速率的影响
熏蒸土壤CO2排放速率与未熏蒸相比在培养前7 d 显著增加(图2)(P<0.05)。该现象不仅与传统观念:“微生物量减少会降低SOC 矿化速率”相反,与Kemmitt 等[13]提出的“SOC 的矿化与土壤微生物的生物量大小、群落组成及活性无关”也不一致。Van Elsas 等[14]的研究表明,熏蒸会暂时提高土壤中溶解细胞(含碳)资源的可用性,幸存的微生物在此基础上生长,造成土壤呼吸速率的短期增加;Dungait 等[23]与Brookes 等[24]的研究支持“调节门”假说:SOC 的矿化分为非生物阶段(K1)与生物阶段(K2),K1 控制微生物对可用基质的可及性而非影响微生物本身,熏蒸后CO2排放速率的短期提高,是由于熏蒸处理增加了溶解细胞资源,K1 产物增多;Kuzyakov 等[25]质疑了“调节门”假说,认为CO2排放速率快速增长是由于残存微生物利用微生物死亡体快速增殖的结果;Zhou 等[16]对土壤进行重复熏蒸培养,得出熏蒸后短期CO2的排放增加主要是由于微生物死体释放的碳源,该部分碳源的完全矿化需要6~8 d 的结论。上述研究均表明CO2排放速率在熏蒸后培养初期显著增加是由于熏蒸造成的微生物死亡释放的碳源。本试验中,熏蒸后前7 d 培养CO2排放速率显著提高,此后各处理间差异不显著。
随熏蒸时长增加,土壤中溶解细胞(含碳)资源增多,促进土壤CO2排放速率增加并维持较长时间,本试验中C24 与C2.5 处理CO2排放速率的提高持续到培养第7 天,而C1.5 与C1 处理则持续到第4 天。Van Elsas 等[14]表示“随熏蒸时长的增加,土壤CO2排放速率的最大值也有所增加”,本试验中,CO2排放速率随熏蒸时长的增加出现C2.5 最大值高于C24 的结果,与Van Elsas 等[14]结论不符。其试验熏蒸时长设置更少,不同设置间时长差距较大可能会导致试验结果的片面性,从而影响试验结论。为解释培养前7 d 熏蒸处理CO2排放速率显著高于CK 及C2.5 处理峰值高于C24 处理的结果,我们提出,土壤中溶解细胞(含碳)资源增加的条件下,微生物的生物量大小会对这部分有机碳的矿化产生影响。土壤CO2排放速率最大值随熏蒸时长的增加呈现先增长后下降的趋势,本试验中最大值出现在C2.5 条件下,当熏蒸时间小于2.5 h,土壤中溶解细胞资源较少,限制CO2排放速率,当熏蒸时长超过2.5 h,土壤微生物量大幅降低限制CO2排放速率。
本研究中培养7~53 d,处理间土壤CO2排放速率差异不显著,针对此现象,“调节门”假说认为“SOM的矿化主要受到非生物过程的调控,与微生物生物量大小、群落结构或活性无关”,质疑“调节门”假说的观点则认为:“生物过程是影响SOC 矿化的主要因素。熏蒸后土壤中幸存的微生物增长以R 策略为主,在微生物残体资源上快速繁殖,微生物数量恢复造成排放速率差异不显著;氯仿熏蒸杀死土壤中大部分微生物,但仍有部分存活,在生态位冗余原则下,残存的微生物替代死亡种群的功能,导致土壤微生物生物量不影响SOM 矿化;熏蒸仅影响土壤微生物活性,土壤中不受熏蒸影响的胞外酶是维持基础呼吸速率的关键”[25-26]。Brookes 等[24]指出“熏蒸后微生物种群数量大幅降低,培养10 d 后即使有所恢复,微生物群落组成也会发生巨大改变,微生物生物量约为未熏蒸土壤中种群数量的10%~20%,这小部分微生物无法满足与未熏蒸土壤中微生物相同的作用”;Zhou 等[17]认为“土壤中的胞外酶来源于微生物,且寿命短、易失活,而Kemmitt 等[13]培养不同熏蒸处理的土壤超过两百天后不同处理间CO2排放速率差异不显著,表明胞外酶并非维持基础呼吸速率的原因”;此外,熏蒸后土壤中溶解细胞(含碳)资源增加导致CO2排放速率增加,这部分碳源消耗后,熏蒸与未熏蒸处理CO2排放速率一致,是否由于土壤本身有机质含量不足限制了土壤CO2排放速率?Brookes 等[24]研究表明高有机质含量的土壤熏蒸与未熏蒸处理CO2排放速率仍具有一致性。
针对上述分析推测,自然土壤条件下,SOC 的矿化速率与土壤中微生物生物量的大小无关。本试验结果表示“调节门”假说适用于长期施用猪粪肥处理的石灰性紫色土农田,但我们做出补充,在K1 阶段产物充足的条件下,微生物的生物量大小会对这部分有机质的矿化产生影响。
3.3 熏蒸时长对土壤有机碳CO2 排放累积通量的影响
培养7~53 d 的CO2排放累积量(图3)表明处理间无显著差异,支持“调节门”假说,但未熏蒸处理略高于熏蒸处理。我们认为是由于培养前7 d,溶解细胞(含碳)的增加引起的SOC 矿化的激发效应[27],易分解新鲜碳源为微生物提供能量。Fontaine 等[28]提出当这些能量充足时,微生物会对土壤中存在的难以分解或者分解消耗能量高于得到能量的有机质进行分解,此时外源有机碳作为微生物的能量泵促进土壤微生物对稳定有机碳进行分解,导致熏蒸处理SOC 含量减少,最终导致熏蒸处理7~53 d 内的累积CO2排放通量低于CK。不同熏蒸时长处理CO2累积排放通量符合相同的模型,其中C2.5 处理系数a最大、b最小,熏蒸2.5 h 最大地促进了土壤的激发效应,降低了SOC 含量,导致C2.5 处理CO2累积排放通量的增量随培养时间的增加减小最快,该结果与Fontaine 等[28]的结论一致。
本试验结果支持了“调节门”假说,长期施用猪粪肥处理的石灰性紫色土农田中土壤微生物的生物量大小不影响SOC 的矿化速率,但土壤微生物的研究仍十分重要,土壤中微生物碳库虽然仅占土壤总有机碳库的0.3%~7%,但却占土壤易分解碳库的70%~80%[29],土壤微生物是其他形式有机碳积累、矿化分解的主要影响因素[30]。近些年来有研究表明,真菌和细菌坏死体是土壤稳定有机碳库的主要碳成分[31-32]。Liang 等[33]的研究表明,50%的土壤生物量碳以死生物质的形式储存在土壤中,土壤有机碳中约9%~35%来源于土壤细菌死体,约23%~87%来源于土壤真菌死体。土壤中约有10%的土壤微生物利用微生物死体存活[34]。减少易分解碳库大小,增加稳定有机碳库(细菌、真菌坏死体)的大小,有助于从根本上减少SOC 矿化。
4 结论
土壤微生物量随熏蒸时间增加而减少,微生物细胞膜损坏后,土壤中增加的活性碳组分部分被分解,造成最终可浸提微生物生物量碳含量的降低。因此,熏蒸处理一旦结束应当立即进行浸提,并测定计算土壤中的微生物生物量有机碳,以减少该部分碳的矿化损失,提高结果的准确度。非生物过程产物正常情况下,长期施用猪粪肥处理的石灰性紫色土农田SOC 的矿化与土壤中微生物量的大小无关,支持了“调节门”假说,认为SOC 的矿化与微生物生物量大小、群落组成及活性无关。
本研究针对长期猪粪肥处理的石灰性紫色土农田开展,该结论是否适用于其他土壤条件仍需进一步研究,在本研究结论下,未来针对长期猪粪肥处理的石灰性紫色土农田微生物对SOC 矿化影响的研究应当将更多的精力集中在研究及改变土壤微生物的组成(增加细菌真菌生物量)、探究微生物死体对土壤有机碳库的作用上,而非微生物量大小对土壤有机碳库的作用。
