亚热带红壤区土壤氮添加对杂交构树光合作用的影响*
2022-11-15刘铭王景升
刘铭,王景升
(1 中国科学院地理科学与资源研究所 中国科学院生态系统网络观测与模拟重点实验室, 北京 100101;2 中国科学院江西产业技术创新与育成中心, 南昌 330029;3 中国科学院大学, 北京 100049)
光合作用是植物一项重要的生理过程,植物的产量形成 90%以上来自光合作用[1]。光合作用及有机物积累除受环境因素(气温、土壤水分和土壤肥力等)影响外,还由植物的种类决定[2-3]。大量研究表明,氮肥施用量对作物光合作用有显著影响。氮素是组成蛋白质、核酸和叶绿素等的基本元素,通过参与复杂的生理过程来调节植物的光合作用[4]。鞠正春和于振文[5]研究表明,追施氮肥可以改善小麦旗叶PSⅡ反应中心活性,提高光化学猝灭参数、最大光化学效率和实际光化学效率,促进光合作用。蒋万[6]通过研究证明施氮可以显著提升木薯叶片的叶绿素含量,提高其光合速率。魏猛等[7]的研究表明,增加氮素不仅显著提高叶菜型甘薯的叶绿素含量,还提高了气孔导度及净光合速率。目前已进行的田间试验表明施肥可以明显提升杂交构树的产量,但杂交构树光合生理对氮肥的响应研究尚存空白。
杂交构树是中国科学院植物研究所科技人员在收集野生构树资源基础上,通过杂种选育结合现代生物技术培育的木本饲用新品种,具有高蛋白、产量高、抗逆性强、一年种植多年收割等特点,已被广泛应用于畜禽养殖和产业扶贫等国家战略中。中国地域广泛,气候类型多样,大多数地区均适合杂交构树生长,但不同气候、土壤条件下其产量和品质有很大的差别。在赣江流域的红壤丘陵区,突出的问题是土壤氮素营养普遍不足,影响作物的产量和品质。本研究在施用氮肥情况下,测定杂交构树光合生理学参数,探讨高产优质形成机制,为杂交构树丰产栽培提供理论依据。
1 材料与方法
1.1 试验材料与试验设计
试验于2020年5月至8月末在江西省吉安市泰和县灌溪镇中国科学院千烟洲试验站(115°04′13″E,26°44′48″N)“杂交构树种养殖一体化试验示范基地”进行。试验材料为一年生“科构101”杂交构树树苗,选取高度、分蘖数、地径基本一致的健康的苗木移栽至长3 m、宽2 m的试验样方中,每样方栽种10株,样方中按2列栽种,每列5株,每株间隔0.5 m。试验所施用的氮肥为尿素(含氮46%),于树苗栽种20 d后按下述试验梯度追肥一次。
试验共设置6个梯度:对照CG(施加氮肥0 kg/ha)、低氮N1(7.5 kg/ha)、中低氮N2(15.0 kg/ha)、中氮N3(22.5 kg/ha)、中高氮N4(30.0 kg/ha)、高氮N5(37.5 kg/ha)。各梯度设置3个试验样方,作为3次重复。
1.2 测定方法
于施肥后45~50 d,杂交构树植株生长至1.5 m左右时,择晴好天气,于9:00—11:00测定下述光合参数。测量时在各处理样方中随机选取3株杂交构树,选择植株冠层顶部向阳面、长势和叶龄相近成熟健康叶3片,重复测量3次,取平均值。
采用Li-6400便携式光合系统测定仪(美国LI-COR公司生产)测量杂交构树光合作用一系列参数,主要包括:净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)。
采用Li-6400便携式光合系统测定仪测定净光合速率的光响应曲线,用与Li-6400相匹配的红蓝光源模拟光辐射强度1 800、1 500、1 200、1 000、800、600、400、200、100、50、20、0 μmol·m-2·s-1共12个光照强度,使用CO2注入系统,CO2浓度设定为400 μmol·mol-1,流速设定为 500 μmol·s-1。
采用SPAD-502便携式叶绿素含量测定仪(日本美能达公司生产)测定叶绿素相对含量(SPAD值)。
叶绿素荧光参数采用mini-PAM-Ⅱ超便携调制叶绿素荧光仪(德国WALZ公司生产)测定,包括:最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)、表观光合电子传递速率(electron transport rate,ETR)、光化学猝灭参数(qP)和非光化学猝灭参数(qN)。
1.3 数据处理
采用 Microsoft Excel 2013进行数据的整理汇总。采用SPSS 21.0统计分析软件进行方差分析和相关分析,处理间差异用Duncan多重比较实现。采用Sigmaplot 10.0进行图表制作。
1.4 净光合速率光响应曲线的模型构建
1)净光合速率的光响应曲线模型
直角双曲线模型(rectangular hyperbola model,RHM)[8]:
(1)
非直角双曲线模型(non-rectangular hyperbola model,NRHM)[9]:
(2)
指数模型(exponential function model,EFM)[10]:
Pn=Pn max(1-e-αI/Pn max)-Rd.
(3)
修正直角双曲线模型(modified rectangular hyperbola model,MRHM)[11]:
(4)
式中:Pn为净光合速率;I为光照强度;α为初始量子效率;θ为反映光响应曲线弯曲程度的参数,范围为0<θ<1;Pn max为最大净光合速率;Rd为暗呼吸速率;β与γ为系数,其中β为修正系数。
2)模型精度评价
采用决定系数R2、均方误差(mean square error,MSE)、平均绝对误差(mean absolute error,MAE)检验4个模型拟合的优良程度,计算公式如下:
(5)
(6)
2 结果与分析
2.1 杂交构树净光合速率的光响应过程
在亚热带红壤丘陵区,日均气温为23.78 ℃,日均大气湿度为87.6%,土壤10~20 cm处温度为28 ℃,10~20 cm土壤体积含水量为7.8%的条件下,无氮添加杂交构树的光响应实测值及模型拟合曲线见图1。在光照强度超过50 μmol·m-2·s-1后,净光合速率开始迅速上升,当光照强度达到600 μmol·m-2·s-1后,净光合速率增速明显趋缓。光照强度1 200 μmol·m-2·s-1其净光合速率达到最大值13.09 μmol·m-2·s-1,之后随着光照强度的继续增加,净光合速率处于波动状态甚至出现下降的趋势。

图1 杂交构树光响应实测及模拟曲线
采用直角双曲线模型、非直角双曲线模型、指数模型、修正直角双曲线模型对光响应曲线进行拟合,并通过比较R2、MSE、MAE的大小对各模型的拟合效果进行分析[12],选取光合参数模拟的最优模型。拟合优良度见表1,MRHM、EFM、NRHM等3种模型的R2均为0.999,MSE和MAE越小,表明拟合值越接近观测值[13]。MRHM模型的MAE和MSE分别为0.114和0.019,都是4个模型中最小值,故本文选择MRHM模型为杂交构树最佳光响应曲线拟合模型。根据模型计算其光补偿点为24.2 μmol·m-2·s-1,光饱和点为1 147.6 μmol·m-2·s-1。

表1 杂交构树光响应曲线模型选择
与其他饲草[14-16]相比,杂交构树光补偿点低于柳枝稷、山羊豆、紫花苜蓿、草木樨,光饱和点与紫花苜蓿相当,略高于草木樨,远高于白三叶、山羊豆和柳枝稷,见表2。LCP较低和LSP较高的特点,为杂交构树具有较高的有机物积累能力奠定了基础。

表2 不同饲草(牧草)光补偿点、光饱和点比较
添加适量氮素对杂交构树光响应曲线的影响见图2,光补偿点及光饱和点没有明显改变,但当光照强度高于50 μmol·m-2·s-1时,土壤氮素添加能显著提高净光合速率,且在光照强度为1 000 μmol·m-2·s-1时达到Pn最大值21.46 μmol·m-2·s-1,较无氮素添加组提升63.94%,为杂交构树的追肥增产提供了数据支持。

图2 土壤中添加氮素对杂交构树光响应曲线的影响
2.2 土壤氮添加对杂交构树光合作用的影响
氮添加水平对各个测定项目的方差分析的差异性结果见表3。由表3可知,不同氮添加处理对于净光合速率、叶片水分利用效率(water use rate,WUE)、叶绿素相对含量SPAD值均有显著性差异(P<0.05),而对于气孔导度Gs、胞间CO2浓度Ci的差异性不显著。不同氮添加处理对于最大光化学效率Fv/Fm、实际光化学效率ΦPSⅡ和表观光合电子传递速率ETR有显著性差异,而对于光化学猝灭参数qP和非光化学猝灭参数qN差异不显著。

表3 氮添加水平对各测定项目的方差分析结果
2.2.1 土壤氮添加对净光合速率的影响
随着土壤氮素添加量的增加,杂交构树净光合速率逐渐增加,当氮添加量达到22.5 kg/ha(N3)时,杂交构树净光合速率达到最高值20.90±0.87 μmol·m-2·s-1(见图3);之后,随着土壤氮添加量的增高,净光合速率表现为缓慢下降趋势。总体而言,土壤氮素添加,能明显提高杂交构树的净光合速率,与对照组相比,分别提升26.88%、37.73%、66.77%、53.14%、58.55%(图3)。

不同小写字母表示P<0.05显著差异(下同)
2.2.2 土壤氮添加对叶绿素合成的影响
在众多的光合色素中,叶绿素含量对净光合速率的影响最为显著[17-19]。在亚热带红壤区,土壤中添加氮素能明显增加杂交构树叶中叶绿素相对含量(见图4),随着土壤中添氮量的增加,杂交构树叶片中SPAD值增加,当土壤氮素添加量达22.5 kg/ha时,SPAD值最高达48.77±1.45,之后,土壤氮素继续增加则SPAD值减少,净光合速率随之降低(图4,图3)。综合分析表明,在一定范围内,土壤中添加氮素,能促进杂交构树叶片中叶绿素含量增加,前述5个氮添加水平中SPAD值分别比对照提高4.47%、8.79%、12.80%、7.17%、10.10%(图4)。
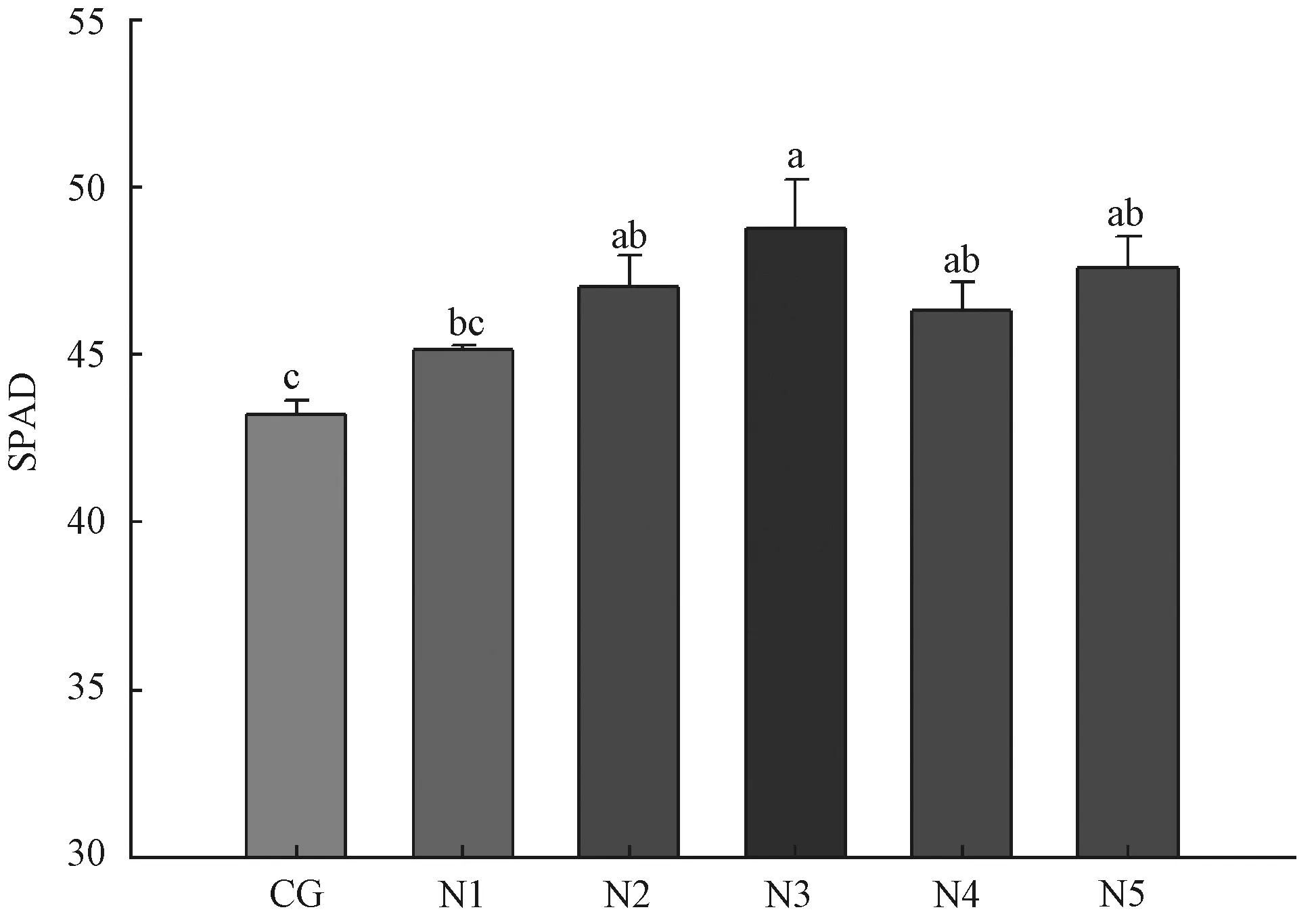
图4 土壤氮素添加与叶片叶绿素含量关系
2.2.3 氮添加对气体交换参数的影响
一般情况下,植物的净光合速率随着气孔导度的增大而增大,增大速率逐渐减小[19-21]。在土壤少量添加氮素(N1、N2)的条件下,杂交构树的气孔导度却略低于对照,但组间差异不显著,随着添加氮量的增加,气孔导度缓慢增加,当氮素添加量超过22.5 kg/ha时,与对照组相当;胞间CO2浓度随氮添加水平的提高呈先下降后上升的趋势,但组间差异不显著;叶片水分利用效率随氮添加水平的增加总体呈现显著先上升后下降的趋势,N3水平叶片水分利用效率最高,为2.16±0.11 μmol·mol-1,是对照组的近2倍(图5)。

图5 土壤氮素添加与叶片气体交换参数关系
2.2.4 氮添加对叶绿素荧光参数的影响
如图6所示,随氮添加水平的增高,最大光化学效率总体增加,N3水平达到峰值,为0.837±0.002,N3~N5组间差异不显著,均较对照组明显升高,分别升高1.58%、1.46%、1.21%。随氮添加水平的提升,实际光化学效率增加,于N3处理水平达到峰值,为0.24±0.005,较对照组提高22.45%,后缓慢下降。表观电子传递效率随氮添加水平的增高而增加,于N3处理水平达到最大值,为83.37±1.82 μmol e-·m-2·s-1,较对照组提高22.60%,后缓慢下降。氮添加水平的变化对光化学猝灭参数和非光化学猝灭参数无显著影响。综合分析表明,在土壤中添加氮素,能提高杂交构树对环境因子的抗性,提升PSⅡ反应中心开放的比例和表观电子传递效率,促进叶片的光合作用。

图6 土壤氮素添加与叶片叶绿素荧光参数关系
2.2.5 光合参数相关性分析
氮添加水平与杂交构树各项测定参数的相关性分析结果见表4:氮水平与Pn、Gs、Ci、WUE、SPAD、Fv/Fm、ΦPSⅡ、ETR、qP及qN的相关系数(γ)分别为0.809、0.238、0.089、0.935、0.622、0.576、0.618、0.622、0.365、0.071(|γ|>0.8时,两变量间高度相关;0.5≤|γ|≤0.8时,两变量间中度相关;0.3≤|γ|<0.5时,两变量间低度相关),说明氮添加水平与Pn、WUE呈高度正相关关系,与SPAD、Fv/Fm、ΦPSⅡ和qP呈中度正相关关系。对应的显著性P值为0、0.394、0.754、0、0.013、0.025、0.014、0.013、0.181、0.8,即氮添加水平与SPAD、Fv/Fm、ΦPSⅡ、ETR的相关性显著,与Pn、WUE的相关性极显著。
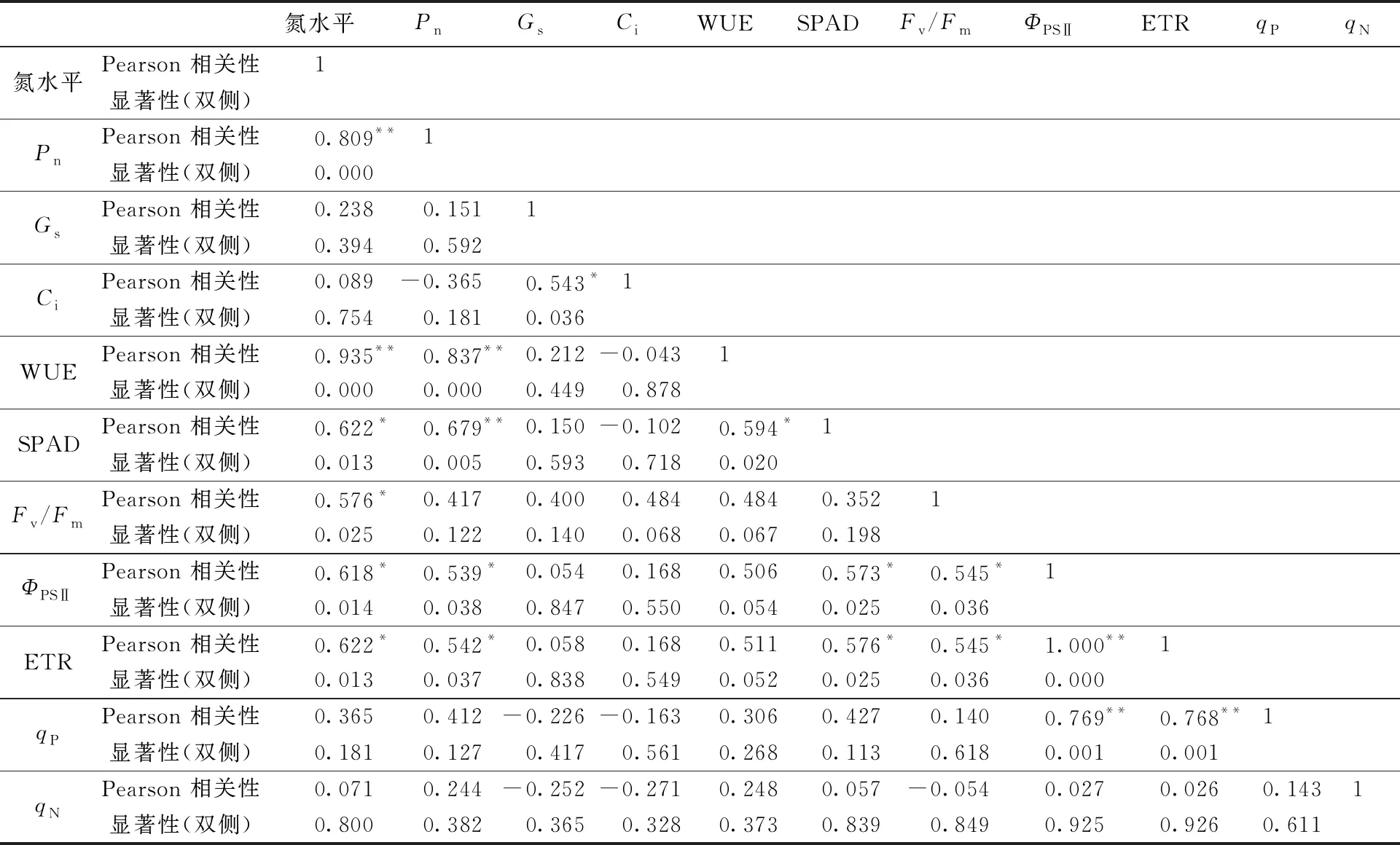
表4 氮添加水平与各测定参数的相关性分析
Pn与Gs、Ci、SPAD、Fv/Fm、ΦPSⅡ、ETR、qP及qN的相关系数(γ)分别为0.151、-0.365、0.679、0.417、0.539、0.542、0.412、0.233,对应的显著性P值为0.592、0.181、0.005、0.122、0.038、0.037、0.127、0.382,即Pn与SPAD、ΦPSⅡ、ETR呈现显著中度正相关关系,而与Gs、Ci、Fv/Fm、qP及qN的相关性不显著。
3 讨论与结论
3.1 讨论
光合作用是为植物生长提供物质和能量的重要生理过程,是植物产量形成的基础[22],植物叶片的光合能力是影响植物生产力和产量的重要因素[19],光照、土壤肥力等多种环境因素均会影响植物的光合能力。
植物叶片的光饱和点和光补偿点反映了植物对光照环境的适应能力,光补偿点较低、光饱和点较高的植物有较强的光环境适应性[23]。与柳枝稷、山羊豆、紫花苜蓿、草木樨、白三叶等常见牧草相比,杂交构树的光补偿点仅高于白三叶,低于其余牧草品种,光饱和点高于柳枝稷、山羊豆、白三叶,与紫花苜蓿和草木樨基本持平,说明杂交构树有较强的光环境适应能力,有利于其进行光合作用和光合有机物积累,是一种高产的木本饲用作物。
氮是影响植物生长的重要因子,叶绿体是植物叶片进行光合作用的主要场所,植物叶片中的氮绝大部分都存在于叶绿体中。一些研究[24-25]表明,施加氮肥可以提高植物的光合能力,但也有研究[26-27]认为植物的光合作用与施氮量之间呈非线性关系,当施氮量超过一定水平后,施氮不会提高植物的光合速率,过量施氮甚至会使得植物体内的光合有机产物水解[28],抑制植物的光合作用,可见氮素对植物光合作用的影响存在一定阈值。本研究中,杂交构树净光合速率与氮添加呈高度正相关关系,在N3及以下氮添加处理水平,净光合速率随氮添加水平增高而增强,而在N4及以上氮添加水平时杂交构树净光合速率降低,与上述结论一致。
一般认为,土壤氮素对植物光合作用的影响可分为气孔因素和非气孔因素两类[29-30]。气孔是植物进行气体交换的门户[31],当植物受到气孔因素限制时,植物叶片气孔导度下降,阻碍CO2进入叶片,抑制叶片的光合作用;当植物受到非气孔因素限制时,植物光合器官的光合活性下降,光合作用减弱[32]。本研究中,杂交构树的气孔导度与净光合速率不存在显著相关性,在N3及以下水平,随着施氮量的增加气孔导度无显著变化,而净光合速率显著增高,说明本研究中施氮未能对气孔导度产生明显影响,此时杂交构树的光合作用不受气孔限制。此结论与闫慧等[33]对亚热带树木幼苗秃瓣杜英、枫香、木荷、青冈的研究结论一致。
氮添加影响植物光合作用的非气孔因素可分为对叶绿素荧光参数的影响和对叶绿素含量的影响两类。叶绿素荧光动力学参数表达着光合作用过程的重要信息,是反映光合作用系统的“内在性”特点的参数[34]。前人研究认为,氮添加能促进 PSⅡ 反应中心的开放程度,提高植物叶片实际光化学效率和电子传递速率,进而提高光合速率[27,35]。而氮素过量会造成土壤环境产生逆境胁迫的条件,可能导致植物对水分和氮肥的吸收受到抑制,降低植物叶片的潜在活性[36]。本研究中,在N3及以下水平,杂交构树叶片的实际光化学效率随着氮添加量的增高而显著增加,表明氮添加对杂交构树净光合速率的提升作用可能是由于随着施氮量增加叶绿素荧光参数得到显著改善;在N4~N5水平,净光合速率、实际光化学效率和表观光合电子传递速率均较N3水平有所下降,说明此时氮添加量过多,对杂交构树的光合作用造成一定程度的抑制。
叶绿素是植物叶片进行光合作用的必要条件和主要物质,叶绿素含量直接影响植物的光合能力[37]。叶绿素含量受氮添加水平的影响,低氮通常导致叶绿素含量较低[38],有研究认为增加氮素可以提高植物叶片的叶绿素含量[6]。另一方面,高氮累积到一定程度会产生毒副作用,可能抑制植物叶片中的色素[39-40]。在前人研究中,采用叶绿素相对含量(SPAD值)来代表植物叶绿素的含量已被广泛认可和使用,有研究表明SPAD值与叶绿素绝对含量显著相关[41-42]。本研究中,SPAD随氮添加水平的增高呈先升高后趋于平缓的趋势,与上述研究结论相一致。同时,SPAD与净光合速率呈中度正相关,随氮添加变化的趋势基本一致,也证明了本氮添加控制试验中,叶绿素含量是影响杂交构树光合速率的因素之一。
3.2 结论
1)修正直角双曲线模型更适合于模拟杂交构树的光响应曲线,杂交构树的光补偿点为24.2 μmol ·m-2·s-1,光饱和点为1 147.6 μmol ·m-2·s-1;较低的光补偿点和较高的光饱和点为杂交构树高产奠定了理论基础。
2)土壤氮素添加能显著提高杂交构树净光合速率,每公顷追肥22.5 kg尿素,叶绿素相对含量增加至48.8,增加了12.8%,促使杂交构树净光合速率提升66.8%,此时水分利用效率较对照也有显著提高,为构树饲料的施肥增产提供了参考数据。
3)每公顷添加22.5 kg尿素可使最大光化学效率值达到0.837±0.002,实际光化学效率增加22.45%,表观光合电子传递速率提高22.60%,氮添加对光化学猝灭参数和非光化学猝灭参数无显著影响。
