妊娠后期添加丁酸钠对湖羊母羊生长性能、养分表观消化率、血清抗氧化和免疫指标及羔羊生长性能的影响
2022-11-15张昕妍段春辉杨若晨张赛伟张英杰
张昕妍 段春辉 杨若晨 张赛伟 李 菲 张英杰*
(1.河北农业大学动物科技学院,保定 071000;2.河北省畜牧总站,石家庄 050000)
湖羊为我国著名高繁品种,因其具有早熟、四季发情、每胎多羔、抗逆性强等优点[1],在国内肉羊养殖行业中备受关注。母羊在妊娠期间不仅需要满足自身营养需要,还要满足胎儿的生长发育。研究表明,胎儿重量的80%~90%是在妊娠后期增长的[2],因此母羊妊娠后期的营养摄入对胎儿发育及初生重至关重要。母羊妊娠后期最大的代谢特点是由于胎儿快速生长发育压迫胃肠道,干物质采食量(DMI)降低,易使母羊出现能量负平衡,导致脂肪动员加剧,产生大量非酯化脂肪酸,加重肝脏负担,损害母畜机体健康,降低生产性能[3-5]。在能量负平衡状态下,机体动员脂肪、蛋白质供能增加,极易导致机体代谢、免疫状态的改变,使母畜免疫力下降,死淘率上升。
丁酸钠的有效成分是丁酸,属于短链脂肪酸[6]。对反刍动物而言,短链脂肪酸的吸收主要在瘤胃进行,可转化为能量物质供能[7]。研究发现,将丁酸钠添加到犊牛代乳粉中可提高犊牛生长速度和饲料转化率,可能与改善犊牛胃肠道消化酶活性和消化能力有关[8]。Kai等[9]研究表明,丁酸钠可以减轻由高精料造成的山羊瘤胃上皮局部炎症,并保护瘤胃上皮的完整性[10]。Honma等[11]发现,在饲粮中补充丁酸钠可以降低脂肪合成速率,刺激肝脏中脂肪酸氧化,从而抑制脂肪积累。妊娠后期母畜极易出现采食量降低、免疫能力下降等问题,丁酸钠对提高幼龄动物的生长性能、改善消化能力和增强免疫机能有一定的作用[8]。但关于丁酸钠在哺乳动物妊娠后期是否可以促进采食量、改善机体免疫水平等问题鲜有研究。因此,本试验在母羊妊娠后期添加不同剂量丁酸钠,研究其对母羊生长性能、养分表观消化率、血清抗氧化和免疫指标及羔羊体重的影响,旨在为母羊妊娠后期添加丁酸钠改善健康状况、提高生长性能奠定理论基础。
1 材料与方法
1.1 试验时间与地点
本试验在沧州孟村县河北保森畜牧有限公司进行,试验时间为2021年6月—2021年11月。
1.2 基础饲粮
1.3 试验设计
选择在配种期同期发情且妊娠90 d的体况良好、体重相近的湖羊母羊60只,随机分为4组:对照组(基础饲粮)、A组[基础饲粮+2.5 g/(d·只)丁酸钠]、B组[基础饲粮+5.0 g/(d·只)丁酸钠]、C组[基础饲粮+7.5 g/(d·只)丁酸钠],每组3个重复,每个重复5只羊。母羊妊娠90 d开始饲喂丁酸钠,预试期7 d,直至产羔,产羔后所有试验羊饲喂哺乳期TMR。在母羊妊娠第132天进行消化代谢试验。试验期为妊娠母羊产前60 d至产后30 d。
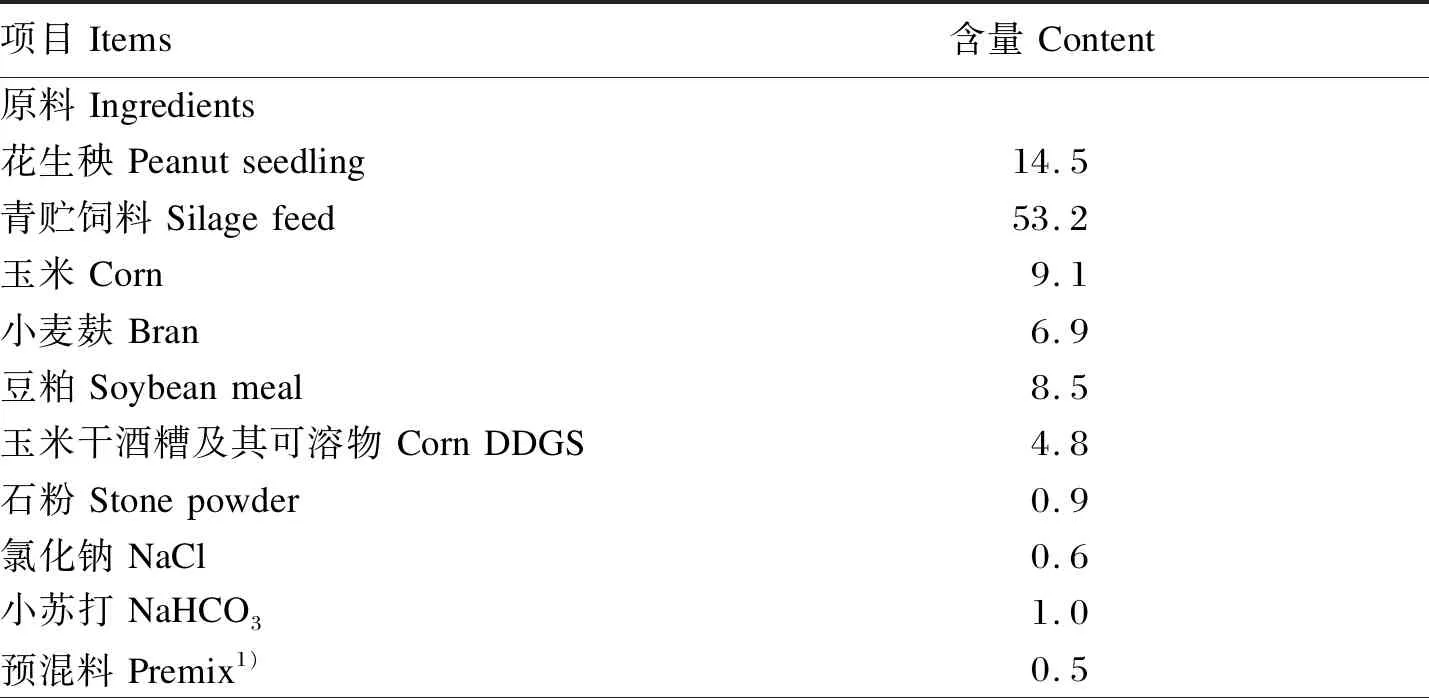
表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet (air-dry basis) %
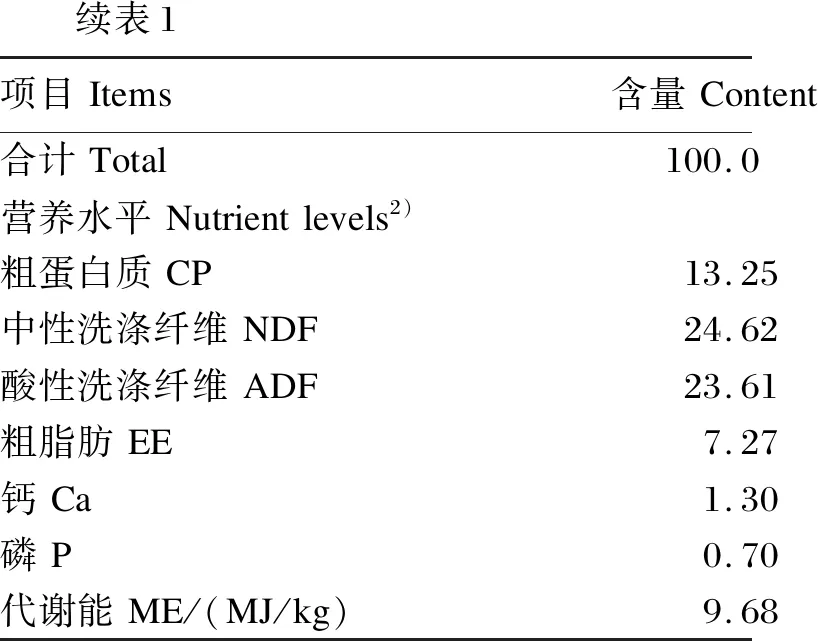
续表1项目Items含量Content合计Total100.0营养水平Nutrientlevels2)粗蛋白质CP13.25中性洗涤纤维NDF24.62酸性洗涤纤维ADF23.61粗脂肪EE7.27钙Ca1.30磷P0.70代谢能ME/(MJ/kg)9.68
试验母羊每日清晨正常饲喂时,将丁酸钠均匀拌入少量TMR中,确保采食后进行正常饲喂TMR。产前60 d至分娩,每天称量给料量与剩料量,记录平均日采食量,计算DMI。在产前60 d、产前15 d和产后14 d于清晨空腹称量母羊体重并记录,随时观察母羊精神状况和健康情况。产前60 d、15 d及产后14 d,从每组中随机选10只母羊,并记录耳号,于晨饲前进行颈静脉无菌采血,置于5 mL促凝管中,3 000 r/min离心10 min。吸取上层血清,分装到1.5 mL无菌离心管中,置于-20 ℃冷冻保存,用于测定血清中抗氧化、免疫指标。母羊分娩当天记为第0天,在母羊产后1、7、14 d,测定母羊泌乳量并记录。在产后1和30 d,早晨空腹称重各组试验羔羊的体重。
1.4 消化代谢试验
消化代谢试验在正式试验饲喂第42天后进行,每组选择体重相近的3只试验羊进行消化代谢试验,用收粪袋收集全部粪样,预试期4 d,正试期3 d。正试期开始后每天采集每只羊的饲粮和剩料样品,记录每只羊的日采食量,计算DMI,并采用全收粪法收集粪样。每天称取并记录每只羊的排粪量,按10%取样,将每只羊3 d的粪样混合后冷冻保存,每100 g粪便加入10 mL 10%的硫酸溶液固氮,再置于-20 ℃冰箱保存。然后将收集到的粪样解冻、混匀,最后和饲粮样品一起放入烘箱中65 ℃烘干,粉碎后备用。
1.5 测定指标与方法
1.5.1 生长性能
羔羊平均日增重(g/d)=[羔羊1月龄重(g)-羔羊初生重(g)]/试验天数(d);母羊DMI(kg/d)=[试验期内每组羊总给料量(kg)-试验期内每组羊总剩料量(kg)]/[试验天数(d)×组内试验羊只数]。
1.5.2 繁殖性能
平均产羔数(只)=产羔总数/分娩母羊数;分娩率(%)=100×每组分娩母羊数/妊娠母羊数;死胎率(%)=100×产死羔母羊数/分娩母羊数;单羔率(%)=100×产单羔母羊数/分娩母羊数;双羔率(%)=100×产双羔母羊数/分娩母羊数;多羔率(%)=100×产3羔以上母羊数/分娩母羊数。
采用羔羊体重之差法测定母羊泌乳量[13]。将羔羊与母羊隔离4 h后放回到母羊身边,30 min后对羔羊进行体重称量,每日早、中、晚各测定1次,泌乳量为3次测定的总和[14]。
1.5.3 养分表观消化率
粪便和饲粮样品中干物质含量采用105 ℃烘干法测定,粗蛋白质含量采用凯氏定氮法测定,粗脂肪含量采用索氏浸提法测定,中性洗涤纤维和酸性洗涤纤维含量采用范式纤维分析法测定,能量采用燃烧热法测定,钙含量采用高锰酸钾滴定法测定,磷含量采用钼黄比色法测定。测定方法依据《饲料分析及饲料质量检测技术》(第2版)[15]。
在企业发展进程中,天业集团始终把培养、造就一流人才与企业生产经营并重,积极探索和打造吸引人才的新途径,通过创新机制,与石河子大学联合开设“屯垦戍边班”、举办“知识竞赛”等方式,扎实推进企业文化进校园,以此吸引、培养和引进人才,这些宝贵的人才资源,成为支撑企业不断科学发展、持续创新的支柱。
1.5.4 血清抗氧化及免疫指标
血清中超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)活性,总抗氧化能力(T-AOC),免疫球蛋白G(IgG)、白细胞介素-2(IL-2)、白细胞介素-4(IL-4)、白细胞介素-6(IL-6)、丙二醛(MDA)含量采用试剂盒测试。所用试剂盒购自南京建成生物工程研究所,按说明书要求严格测定。
1.6 数据统计与分析
试验数据采用SPSS 25.0软件进行单因素方差分析(one-way ANOVA),差异显著后进行Duncan氏多重比较,结果均以“平均值±标准误”表示,以P<0.05为差异显著。
2 结 果
2.1 丁酸钠对妊娠后期湖羊母羊生长性能的影响
由表2可知,B组母羊产前60 d至生产时DMI有高于对照组和A组的趋势,但差异不显著(P>0.05)。在产前60 d和产后14 d,各组间母羊体重无显著差异(P>0.05);在产前15 d时,随着丁酸钠添加量的增加,母羊体重显著线性变化(P<0.05),B、C组母羊体重显著高于对照组(P<0.05),B组显著高于A组(P<0.05)。
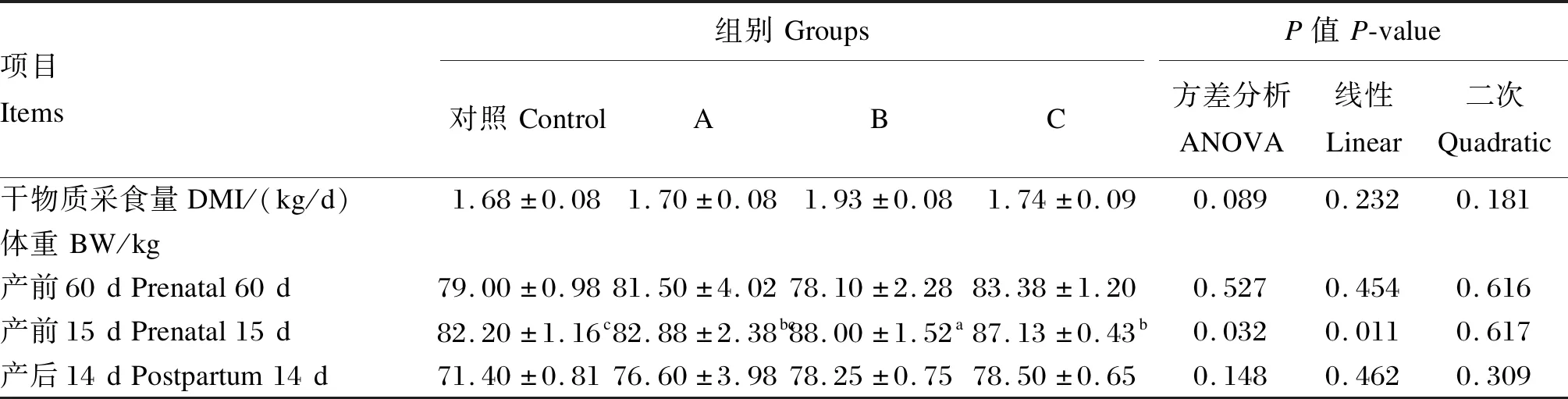
表2 丁酸钠对妊娠后期母羊生长性能的影响Table 2 Effects of sodium butyrate on growth performance of ewes at late gestation
2.2 丁酸钠对妊娠后期湖羊母羊产羔数和泌乳量的影响
由表3可知,对照组和A组母羊的死胎率为7%,B、C组的死胎率为0。B组平均产羔数为2.07只,对照组平均产羔数为2只,A、C组平均产羔数为1.93只。由表4可知,产后1、14 d各组母羊泌乳量没有显著差异(P>0.05);产后7 d,随着丁酸钠添加量的增加,母羊泌乳量呈显著线性变化(P<0.05),A组与对照组差异不显著(P>0.05),B、C组显著高于对照组(P<0.05)。
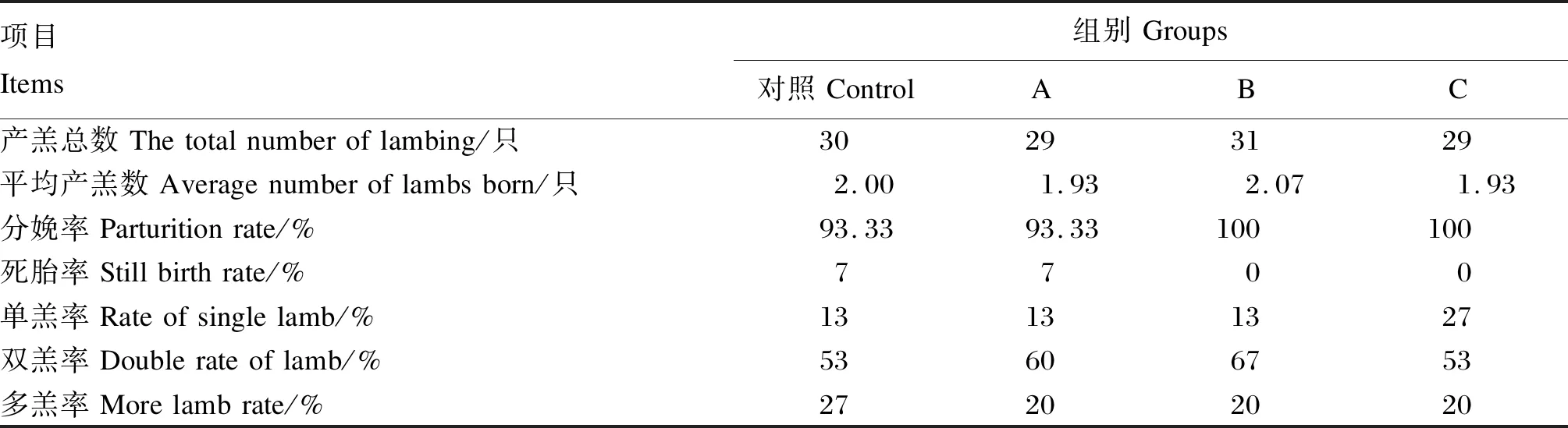
表3 丁酸钠对妊娠后期母羊产羔数的影响Table 3 Effects of sodium butyrate on lamb number of ewes in late gestation
2.3 妊娠后期湖羊母羊饲喂丁酸钠对新生羔羊生长性能的影响
由表5可知,妊娠后期母羊饲粮添加丁酸钠对羔羊初生重、1月龄重以及平均日增重有提高趋势,但各组间差异不显著(P>0.05)。
2.4 丁酸钠对妊娠后期湖羊母羊养分表观消化率的影响
由表6可知,B、C组干物质表观消化率显著高于对照组和A组(P<0.05);B组的总能表观消化率显著高于对照组和A、C组(P<0.05),对照组和A组之间差异不显著(P>0.05),呈显著线性和二次变化(P<0.05);A、B、C组粗蛋白质表观消化率差异不显著(P>0.05),但显著高于对照组(P<0.05);B组中性洗涤纤维和酸性洗涤纤维表观消化率与C组差异不显著(P>0.05),但显著高于对照组和A组(P<0.05),随着丁酸钠添加量的增加,母羊干物质、粗蛋白质、中性洗涤纤维和酸性洗涤纤维表观消化率呈显著线性变化(P<0.05);各组间粗脂肪和钙、磷表观消化率未达到显著水平(P>0.05)。
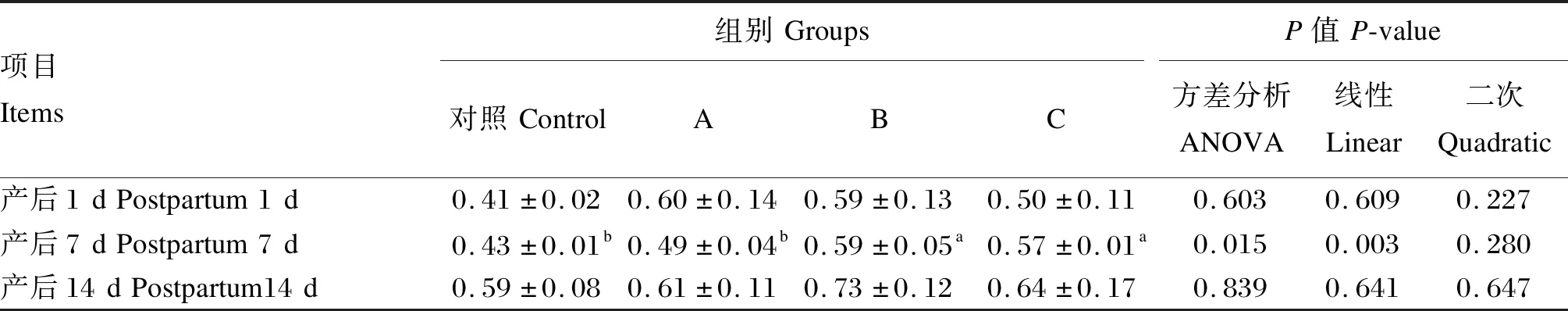
表4 丁酸钠对妊娠后期母羊泌乳量的影响Table 4 Effects of sodium butyrate on milk yield of ewes at late gestation kg

表5 妊娠后期母羊饲喂丁酸钠对新生羔羊生长性能的影响Table 5 Effects of feeding ewes sodium butyrate at late gestation on growth performance of newborn lambs

表6 丁酸钠对妊娠后期母羊养分表观消化率的影响Table 6 Effects of sodium butyrate on nutrient apparent digestibility of ewes at late gestation %
2.5 丁酸钠对妊娠后期湖羊母羊血清抗氧化指标的影响
由表7可知,在产前60 d,各组间血清SOD、GSH-Px活性,MDA含量,T-AOC没有显著差异(P>0.05)。在产前15 d,B组血清SOD活性、T-AOC显著高于A、C组和对照组(P<0.05),A、C组差异不显著(P>0.05);A、B、C组血清GSH-Px活性未达到显著水平(P>0.05),但这3组显著高于对照组(P<0.05);随着丁酸钠添加量的增加,血清SOD和GSH-Px活性、T-AOC呈显著线性和二次变化(P<0.05)。在产后14 d,B组血清SOD活性显著高于A组和对照组(P<0.05),与C组差异不显著(P>0.05),随着丁酸钠添加量的增加,呈显著线性变化(P<0.05);A、B、C组血清GSH-Px活性、T-AOC与对照组相比没有显著差异(P>0.05)。
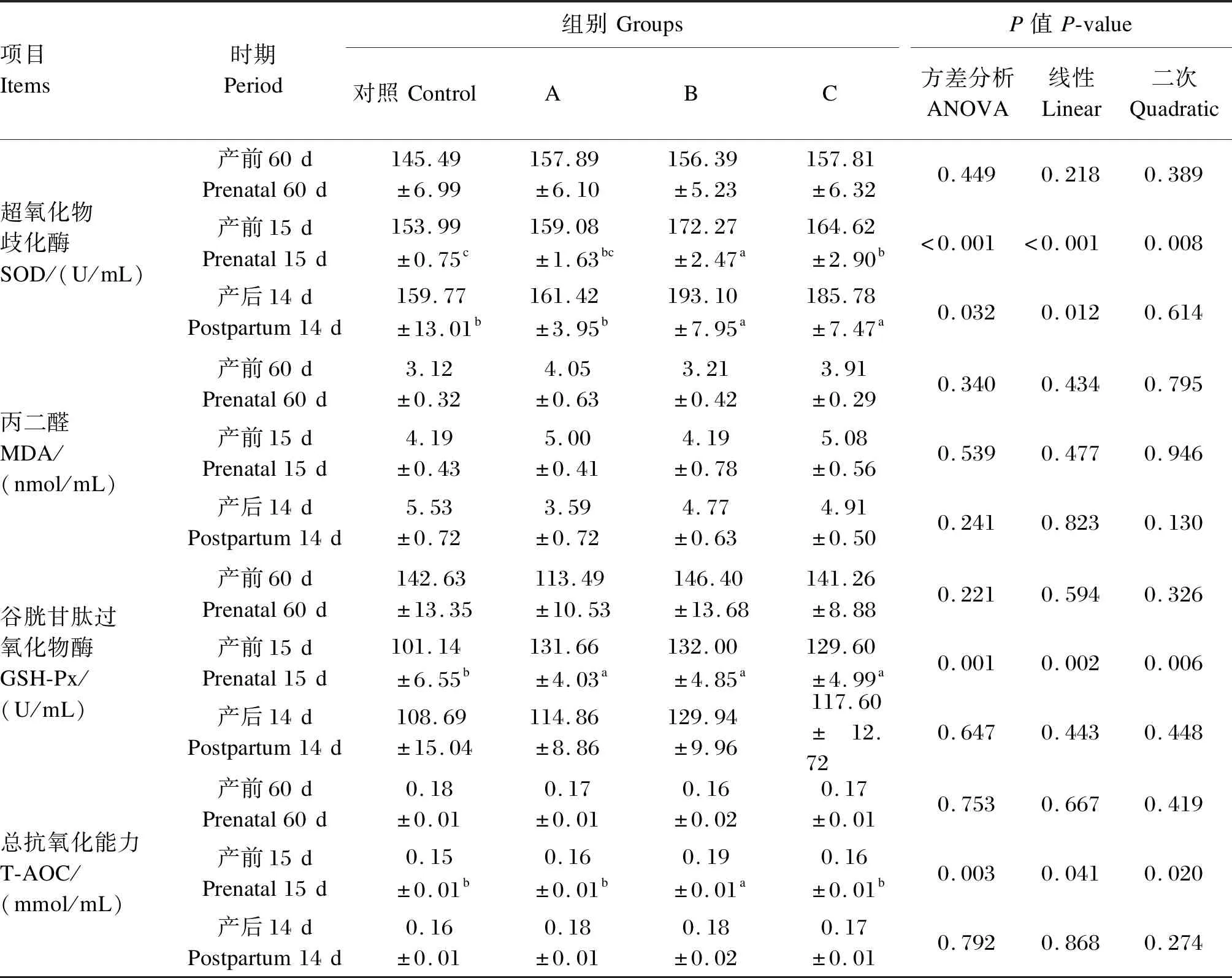
表7 丁酸钠对妊娠后期母羊血清抗氧化指标的影响Table 7 Effects of sodium butyrate on serum antioxidant indexes of ewes at late gestation
2.6 丁酸钠对妊娠后期湖羊母羊血清免疫指标的影响
由表8可知,在产前60 d,各组间血清IgG、IL-2、IL-4、IL-6含量没有达到显著水平(P>0.05)。在产前15 d,B组血清IgG含量显著高于对照组和C组(P<0.05),与A组差异不显著(P>0.05),呈显著线性和二次变化(P<0.05);B、C组血清IL-2、IL-6含量显著低于对照组和A组(P<0.05),呈显著线性变化(P<0.05)。在产后14 d,B、C组血清IL-6含量显著低于对照组和A组(P<0.05),呈显著线性变化(P<0.05);各组间的血清IgG、IL-2和IL-4含量差异不显著(P>0.05)。
3 讨 论
3.1 丁酸钠对妊娠后期湖羊母羊生长性能的影响
母畜在妊娠后期对葡萄糖、氨基酸、脂肪酸的需要急剧增加,但由于胎儿压迫胃肠道使母畜采食量降低,摄入能量不能满足机体需要,使母畜进入能量负平衡的应激状态。岳敏[16]在母猪饲粮中添加丁酸钠显著提高了妊娠后期的平均日采食量,说明丁酸钠对母猪有诱食作用。Guilloteau等[8]报道,在犊牛饲粮中添加丁酸钠提高了生长速度和体增重,说明丁酸钠对幼龄反刍动物有促生长作用,可能是因为刺激胰液体积和胰蛋白酶分泌,改变了胃肠道的形态和功能。本试验结果表明,在饲粮中添加2.5、5.0和7.5 g/(d·只)的丁酸钠可在一定程度上提高母羊妊娠后期DMI,推测原因可能是丁酸钠进入瘤胃后改变了瘤胃内渗透压,瘤胃丁酸含量增加刺激瘤胃壁代谢,促进瘤胃微生物的活动[17],从而提高DMI。研究表明,高剂量丁酸钠会导致动物血清中肿瘤坏死因子-α(TNF-α)和IL-6含量显著降低[18],TNF-α释放到脑室会抑制食欲[19],IL-6通过降低胰岛素样生长因子-1(IGF-1)的产生抑制线性生长[20],因此,高剂量的丁酸钠可降低动物采食量。本研究中,添加5.0 g/(d·只)丁酸钠组母羊的DMI高于添加2.5 g/(d·只)丁酸钠组;添加5.0 g/(d·只)丁酸钠组产前15 d母羊体重高于其他各组,与该组母羊DMI最高的结果相一致,说明本试验条件下湖羊母羊妊娠期添加丁酸钠的最佳添加量为5.0 g/(d·只)。

表8 丁酸钠对妊娠后期母羊血清免疫指标的影响Table 8 Effects of sodium butyrate on serum immune indexes of ewes at late gestation
绵羊的泌乳受品种、年龄、饲养条件等多种因素的影响[21]。研究表明,母牛妊娠期间饲喂高能量饲粮,可明显提高产犊后的产奶量[22]。妊娠后期营养水平的增加会使泌乳量逐步提高,初乳中的乳脂及乳蛋白含量逐步上升[23-24]。以上研究说明,妊娠后期饲粮的营养成分对母畜产后泌乳量及乳品质有影响。本试验中,添加丁酸钠的各试验组母羊产后7 d泌乳量分别较对照组提高了13.95%、37.21%、32.56%,妊娠后期DMI分别比对照组提高了1.19%、14.88%、3.57%,说明母羊泌乳量受妊娠后期营养摄入的影响,可能是由于丁酸钠在动物体内可解离出丁酸根离子,具有短链脂肪酸游离的特性,为动物提供快速的能量来源[25-26]。
3.2 妊娠后期湖羊母羊添加丁酸钠对新生羔羊生长性能的影响
妊娠期母羊的机体状况直接影响胎儿的发育状态,而羔羊哺乳前期所需的营养成分大部分来源于母乳[27],母乳具有适口性好、消化率高、营养全面等优点,是羔羊出生至断奶期间重要的营养来源[28]。研究发现,妊娠后期母羊饲喂高能量饲粮可提高泌乳期泌乳量,从而使羔羊90日龄体重增加[29]。岳敏等[16]在妊娠期和哺乳期母猪饲粮中添加丁酸钠,结果显示母猪的平均日采食量显著提高,仔猪初生均重、断奶均重和平均日增重均显著高于对照组,腹泻率显著降低。本试验中,妊娠后期母羊饲粮中添加2.5、5.0和7.5 g/(d·只)的丁酸钠使羔羊初生重分别提高了5.70%、7.69%和6.55%,这与母羊产前15 d的体重结果相一致,说明母羊妊娠后期添加丁酸钠可以通过改善能量摄入促进羔羊发育,并且各丁酸钠添加组羔羊1月龄的平均日增重较对照组分别提高0.31%、5.45%、4.84%,这与各丁酸钠添加组母羊泌乳量结果相一致。
本试验在妊娠后期添加丁酸钠,对母羊围产期健康水平影响较大,从母羊分娩率及死胎率结果看,添加5.0和7.5 g/(d·只)丁酸钠组相对于对照组和添加2.5 g/(d·只)丁酸钠组提高了母羊分娩率,可能是由于丁酸进入体内不需要通过复杂的三羧酸循环,能快速给胃肠道细胞供能,因此丁酸钠可通过促进能量代谢改善母羊围产期健康,降低死胎率。
3.3 丁酸钠对妊娠后期湖羊母羊养分表观消化率的影响
养分表观消化率是测定动物机体对饲料添加剂消化代谢的能力,养分表观消化率的高低与动物的生长性能密切相关。有研究显示,瘤胃挥发性脂肪酸(尤其是丁酸)可促进胰腺淀粉酶的分泌[30],在犊牛饲粮中添加丁酸钠可提高胰腺分泌和营养物质消化率[8]。研究表明,丁酸能够提高瘤胃微生物的生长代谢,促进纤维素降解酶的产生和分泌[31-32]。支链挥发性脂肪酸是纤维降解菌必需的生长因子[33],而饲粮中添加丁酸可通过提高支链脂肪酸的浓度,从而促进纤维降解菌的生长代谢,当纤维素降解酶活性提高后,有利于促进底物干物质的降解。本试验中,妊娠后期母羊饲粮中添加5.0和7.5 g/(d·只)丁酸钠组的干物质表观消化率高于对照组和添加2.5 g/(d·只)丁酸钠组,而2.5 g/(d·只)的添加量没有显著影响干物质表观消化率,这与Huhtanen等[34]研究结果一致,其研究发现提高奶牛瘤胃中丁酸的浓度,能够增加饲粮干物质的降解率。研究表明,丁酸盐对生长期的反刍动物或成熟期的反刍动物瘤胃上皮细胞的生长产生刺激作用[35-36],可增加瘤胃乳头的长度[37],使营养物质吸收的表面积增大[38],并且激活胃蛋白酶原,促进蛋白质消化[39]。本试验中,添加丁酸钠组可提高妊娠后期母羊粗蛋白质表观消化率,与前人的研究结果一致。添加2.5 g/(d·只)丁酸钠组和对照组中性洗涤纤维、酸性洗涤纤维表观消化率低于添加5.0 g/(d·只)丁酸钠组,推测原因可能是食糜在消化道中的流通速度加快,纤维在消化道停留时间变短而造成消化率下降[38]。研究表明,将丁酸盐直接注入瘤胃,可以增加生长期山羊瘤胃中短链脂肪酸吸收的效率[36]和奶牛总营养素的消化率[34],当短链脂肪酸氧化后可以为肠细胞提供能量,丁酸所提供的能量约为短链脂肪酸耗氧时产生能量的70%[40]。本试验结果表明,妊娠后期饲粮中添加5.0 g/(d·只)丁酸钠可提高总能表观消化率,这可能是因为丁酸钠的添加提高了短链脂肪酸的吸收效率,进而促进了机体对能量的利用率。有研究报道,在断奶仔猪饲粮中添加适量丁酸钠可提高仔猪粗脂肪表观消化率,而对钙、磷表观消化率没有产生显著影响[41]。本试验中,妊娠后期添加丁酸钠对母羊粗脂肪、钙和磷表观消化率没有显著影响。
3.4 丁酸钠对妊娠后期湖羊母羊血清抗氧化指标的影响
国内外关于丁酸钠对动物抗氧化性能的研究主要集中在单胃动物和家禽方面[42]。SOD、GSH-Px、T-AOC是动物机体主要的抗氧化酶,在清除自由基或减少自由基形成、防止生物分子产生损伤等方面发挥重要作用,它们的活性影响机体内活性氧自由基水平及脂质过氧化终末代谢产物MDA;而MDA含量的高低可反映细胞损伤的程度[43]。当MDA含量升高,T-AOC降低,表明促氧化因子压倒抗氧化因子,产生脂质过氧化[44]。MDA是脂质过氧化产生的剧毒副产物,其毒性取决于其与蛋白质和DNA的快速反应[45]。有研究报道,血清MDA含量降低,T-AOC升高,可能导致增加β羟丁酸含量并减轻氧化应激水平[46]。本试验中,添加丁酸钠对各组血清MDA含量没有造成显著影响,说明丁酸钠对血清MDA含量的产生无显著影响。母羊在妊娠后期处于能量负平衡状态,机体产生过量的活性氧,超出机体自身可清除的能力,往往抗氧化指标会有所降低[47]。但本试验中产前15 d,各试验组血清中GSH-Px活性显著高于对照组,产后14 d添加丁酸钠对血清GSH-Px活性有升高趋势,表明添加丁酸钠可以增强母羊产前产后血清GSH-Px活性,增强机体对自由基的清除能力,改善机体的抗氧化功能[48],与周腊枚等[49]研究丁酸钠可以促进机体抗氧化酶相关基因的表达研究结果一致。SOD可减轻自由基导致的氧化损伤,在保护细胞免受损伤方面具有重要作用,T-AOC是衡量动物机体抗氧化系统能力的综合指标[50]。本试验结果显示,妊娠后期添加5.0 g/(d·只)丁酸钠组产前15 d血清SOD活性、T-AOC显著高于对照组和添加2.5、7.5 g/(d·只)丁酸钠组,说明饲粮添加5.0 g/(d·只)丁酸钠对缓解母羊产前氧化应激效果较优。此外,与对照组相比,产后14 d 3个试验组血清SOD活性、T-AOC有升高趋势,说明妊娠后期饲粮中添加丁酸钠有利于阻断自由基对细胞造成的损伤[51],改善机体氧化应激状态。
3.5 丁酸钠对妊娠后期湖羊母羊血清免疫指标的影响
研究发现,丁酸钠具有加强肠道黏膜化学屏障的作用,可刺激肠道黏膜分泌免疫球蛋白,促进免疫细胞的增殖分化,并调节宿主免疫[52],能够抑制炎症细胞因子的产生和释放[53]。免疫球蛋白与动物的免疫能力密切相关,其中IgG是动物免疫系统中较为重要的一部分,在血清中的占比最大,可以达到血清中抗体的70%~80%[54],对维护动物自身的免疫屏障起到重要作用[55-56]。赵会利等[53]研究发现,丁酸钠有提高断奶犊牛血清IgG含量的趋势,从而提高机体免疫功能和抗病力。陈想等[57]研究表明,包膜丁酸钠可以提高羔羊血清中IgG的含量。本试验中,添加丁酸钠可提高母羊产前15 d血清中IgG含量,可能是因为丁酸钠能促进肠道黏膜内淋巴细胞增殖,使淋巴细胞分化成浆细胞的数量增多,提高了免疫系统的灵敏性[53],其中添加5.0 g/(d·只)丁酸钠组血清IgG含量高于添加2.5和7.5 g/(d·只)丁酸钠组,表明添加5.0 g/(d·只)对提高机体免疫功能效果较优,与前人研究结果[58]存在一致性。
白细胞介素具有涉及炎症反应的各种生物功能,可以激活调节免疫细胞和传递信息[59],已发现短链脂肪酸与炎症调节密切相关[60],已有研究证实,丁酸钠中的有效因子丁酸可以减少促炎因子的产生[61-64]。IL-2、IL-4、IL-6属于调节动物健康状态的重要细胞免疫因子,可调节机体炎症反应和细胞免疫应答[42]。刘馨忆[65]在饲粮中添加中剂量组的包被丁酸钠可以显著降低肉仔鸡血清中白细胞介素-1β(IL-1β)和IL-6的含量,增加肉仔鸡血清中白细胞介素-10(IL-10)的含量。本试验中,妊娠后期饲粮中添加丁酸钠可降低产前15 d母羊血清中IL-2和IL-6含量,尤其以添加5.0和7.5 g/(d·只)丁酸钠组效果较优,此外也可降低母羊产后14 d血清IL-6含量。推测原因是妊娠后期添加丁酸钠可减少IL-2、IL-6促炎因子的产生,防止免疫系统的过度活化[66]。
4 结 论
妊娠后期饲粮中添加丁酸钠可提高湖羊母羊的采食量、养分表观消化率,并且可有效改善母羊的抗氧化功能,提高机体的免疫力。在本试验条件下,湖羊母羊饲粮中丁酸钠最适添加量为5.0 g/(d·只)。
