4CL基因在海巴戟叶片莨菪亭累积过程中的功能研究
2022-11-11宫树森王青芬杨自云
宫树森,杨 霏,王青芬,杨自云,吴 田
基因在海巴戟叶片莨菪亭累积过程中的功能研究
宫树森,杨 霏,王青芬,杨自云,吴 田*
西南林业大学园林园艺学院/国家林业和草原局西南风景园林工程技术研究中心/云南省功能性花卉资源及产业化技术工程研究中心,云南昆明 650224
为明确海巴戟叶片中莨菪亭累积规律,探究基因在其累积过程中起到的作用。利用高效液相色谱仪对离体的海巴戟成熟叶0~48 h内莨菪亭含量进行测定,并测定经不同浓度的阿魏酸处理后莨菪亭含量的变化。以阿魏酸为底物,利用紫外分光光度计进行酶活反应的4CL总酶活性测定。对RNA-seq结果中7个基因进行qPCR,筛选出一个表达趋势与莨菪亭含量及酶活变化趋势一致的基因作为关键候选基因,进行全长克隆以及生物信息学分析,最后用qRT-PCR研究其表达特性。海巴戟叶片在采摘后48 h内,莨菪亭含量在12 h时最高,达0.35 mg/g,48 h最低,为0.18 mg/g。4CL总酶活性变化与莨菪亭含量趋势相近。阿魏酸处理后的叶片中4CL总酶活增加。通过qRT-PCR筛选到(DN15707)基因,克隆、测序后发现其长度为1632 bp,具备完整开放阅读框,在NCBI进行序列比对及系统进化树分析,确定该基因为基因家族中的基因,命名为。基因可能是莨菪亭累积过程中的关键基因,它能启动4CL酶活,充分利用阿魏酸底物进而导致莨菪亭累积。
海巴戟;莨菪亭;总酶活;4-香豆酸辅酶A连接酶(4CL);阿魏酸
海巴戟(L.)又称诺丽(noni),是一种茜草科巴戟天属的常绿植物,常为小乔木或灌木[1-2],是一种新兴热带果树[3]。海巴戟中富含莨菪亭,可以抑制骨关节炎,防止关节软骨退化和炎症的发生[4]。莨菪亭也是一种天然的抗癌成分[5],通过直接杀死癌细胞,防止新的癌细胞发展[6],可以用来治疗白血病。此外,海巴戟中的莨菪亭可以通过改善糖原储存和脂质、碳水化合物代谢来缓解疲劳[7],在保护肝脏、抗真菌、解热和降压方面也有重要的作用[8]。由于莨菪亭是海巴戟体内最为主要的活性成分之一,有学者将莨菪亭推荐作为海巴戟的鉴定认证参数、产品质量控制的有效评价参数和药代动力学研究的标记成分[9-10]。
莨菪亭是通过苯丙烷代谢途径而合成的。4-香豆酸辅酶A连接酶(4-coumarate-Co A ligase, 4CL),是苯丙氨酸类化合物合成过程中不可缺少的酶[11],作用于苯丙烷类代谢途径的第3个步骤,是该途径的关键调控位点[12],催化肉桂酸及其衍生物(香豆酸、咖啡酸、阿魏酸、5-羟基阿魏酸和芥子酸)生成相应的CoA[13],从而进入苯丙烷途径下游的不同分支,最终生成木质素、香豆素、黄酮、类黄酮、单宁等物质[14]。4CL可能是莨菪亭合成过程中的关键酶,目前在海巴戟中鲜有报道,其在海巴戟莨菪亭累积过程中的具体功能尚未明确。因此,本研究在探究海巴戟莨菪亭累积规律的基础上,分析4CL酶活与莨菪亭累积的相关性。由于研究海巴戟叶片的最终目的为其次生代谢物的提取和利用,且次生代谢物是评价海巴戟品质的重要标准,根据目前的研究,如袁婷婷等[15]观察阿魏酸对蚕豆()根系细胞组织结构变化、金晶等[16]探究阿魏酸对马缨杜鹃()和露珠杜鹃()种子萌发时的化感作用等使用活体进行阿魏酸处理,而品质类的研究更适合离体处理[17],故本研究选择基于离体条件下进行阿魏酸处理。在此基础上筛选出表达量与酶活及莨菪亭变化趋势一致的关键基因,为后续深入研究该基因功能及解析海巴戟莨菪亭累积机制奠定基础。
1 材料与方法
1.1 材料
海巴戟叶片样品于2019年8月采自云南省玉溪市元江县海巴戟栽培基地,随机摘取不同植株中间层、向阳面成熟叶片,于冰盒中暂存,立即送往实验室待用。
1.2 方法
1.2.1 海巴戟叶片中莨菪亭含量检测 将海巴戟成熟叶片于室温(25℃)下进行取样。使用1、3、5、7、10、15 mmol/L阿魏酸(coolaber,北京)均匀喷施离体海巴戟成熟叶片表面,将叶片放置于玻璃容器中并用薄膜覆盖密封,室温保存,并设置喷水的叶片为空白对照,2 h后进行取样。所有样品取样后均切碎,于60℃烘干箱烘干至恒重,并研磨至粉末后同硅胶干燥剂一起放置于室温避光保存待用。莨菪亭检测参照沈宋利等[18]的方法略有改动:称取样品1 g,按照液料比(g∶mL)1∶32加入53%的乙醇,以频率为45 kHz,温度为50℃进行超声提取33 min,12 000 r/min离心15 min,取上清液过0.45 µm微孔滤膜后进样分析。每个样品设3个重复并且平行测定3次。以乙腈∶水(∶,20∶80)作为流动相进行等度淋洗,以1.0 mL/min的流速,25℃的柱温,20 μL的进样量,检测波长348 nm。莨菪亭标准品(Sigma,美国,HPLC≥99%)的峰保留时间为5 min左右。
1.2.2 线性关系及标准曲线 将配制好的莨菪亭对照品溶液注入高效液相色谱仪进行测定,将得到的峰面积与对照品溶液浓度数据进行整理并绘制标准曲线,得到线性方程=77.043+6.3003(2=0.9998),表明在0.5~20 μg/mL浓度范围的莨菪亭与其色谱峰峰面积线性关系良好。
1.2.3 4CL酶活测定 参照岳凯等[19]和WEI等[20]的方法进行4CL酶活测定,称取样品0.5 g,加入液氮迅速磨成粉末,加入预冷的0.2 mol/L(pH 7.8)的Tris-HCl缓冲液5 mL(含10%的甘油和15 mol/L的巯基乙醇),充分混合,在4℃下10 000 r/min离心20 min,取上清液用于酶活性测定。反应体系3 mL包括:0.5 mL粗酶液,0.2 mmol/L 阿魏酸,5 mmol/L ATP,0.33 mmol/L辅酶A,500 mmol/L Tris-HCl(pH 7.8)和25 mmol/L MgCl2。反应液混合后,于40℃水浴锅反应10 min后加入0.5 mL 6 mol/L盐酸终止反应,将0.5 mL粗酶液换成0.5 mL蒸馏水作为空白对照,反应前和反应后迅速分别测定345 nm处的吸光值,以每克鲜重每分钟吸光度变化0.01为1个酶活性单位,并按以下公式计算酶活性:
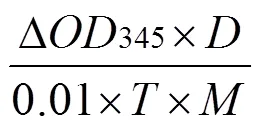
式中,Δ345为反应液在反应时间内吸光度的差值;为稀释倍数,即所提取的总粗酶液与反应酶液的倍数;为反应时间,单位为min;为样品鲜重,单位为g。
1.2.4 基因克隆 实验使用RNA提取试剂盒(Magen,广州)进行海巴戟叶片RNA的提取。将提取得到的RNA进行逆转录合成cDNA第一链,参考TransGene逆转录试剂盒说明书操作。采用NCBI中的Primer-Blast设计基因扩增全长引物4CL(DN15707)-F/R(表1),PCR反应程序:95℃预变性3 min;95℃变性15 s,52 ℃退火20 s,72℃延伸2 min,共35个循环;72℃再延伸10 min。扩增完成后用1%琼脂糖凝胶进行电泳检测,切胶回收,连接到pMD18-T载体上,转入大肠杆菌进行筛选培养,将检测呈阳性的菌液送至生工生物科技(上海)有限公司进行测序。
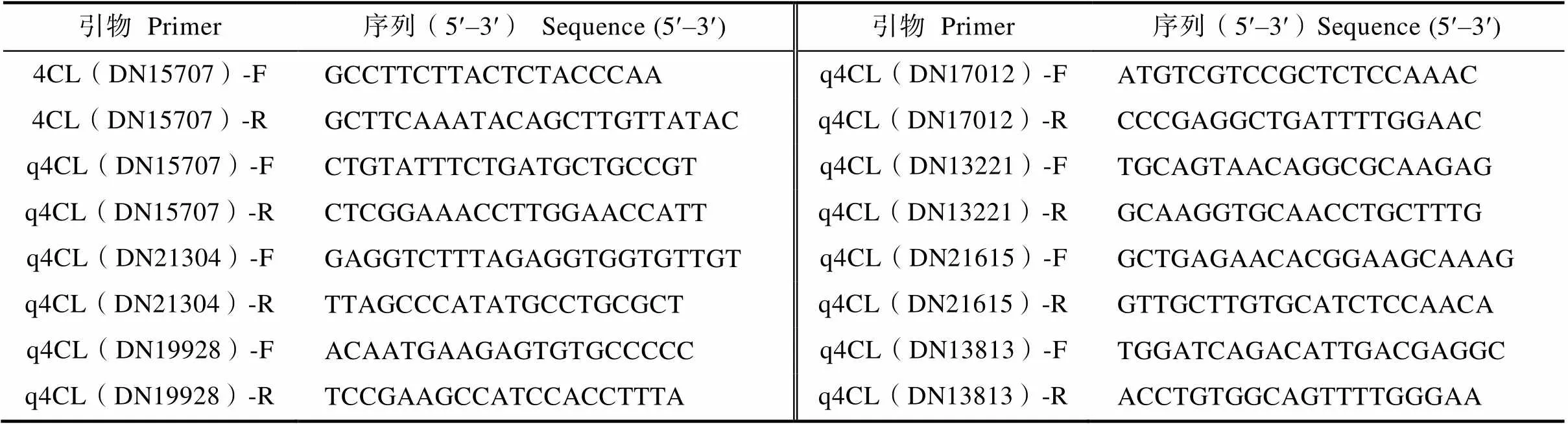
表1 qRT-PCR引物序列
1.2.5 荧光定量 以海巴戟叶片cDNA为模板,基因[21]为内参基因,采用NCBI中的Primer-Blast设计4CL基因特异性引物(表1),根据本课题组基于采后0 h及48 h的条件下所测海巴戟转录组[22],在RNA-seq数据中以“4CL”、“4-coumarate-CoA ligase”及“4-hydroxycinnamoyl-CoA ligase”作为关键词搜索并剔除表达量极低的(表达量为1以下)情况后获得7个基因,选中4个表达量较高且差异较大的基因进行相对定量分析,并使用2‒∆∆CT公式计算其相对表达量。筛选出和莨菪亭含量及酶活变化趋势一致的基因再进行克隆并且进行荧光定量分析。
1.2.6 海巴戟基因的生物信息学分析 将得到的海巴戟(DN15707)序列使用NCBI数据库、ExPASy(https://prosite.expasy.org/)和DNAMAN软件进行序列分析和保守功能结构域分析,利用MEGA-X软件进行基因系统进化树的构建。
2 结果与分析
2.1 海巴戟叶片莨菪亭累积规律
2.1.1 不同浓度的阿魏酸处理海巴戟叶片后莨菪亭含量的变化 海巴戟叶片经不同浓度的阿魏酸处理后除15 mmol/L阿魏酸处理的叶片莨菪亭含量与对照无显著差异外,其他浓度均可刺激叶片的莨菪亭含量增加,其中经5 mmol/L阿魏酸处理后的果实莨菪亭含量显著高于其他浓度条件处理叶片,高达2.5 mg/g,其莨菪亭含量的数值是对照的5倍(图1A)。此实验确定5 mmol/L阿魏酸是诱导海巴戟莨菪亭大量累积的最佳浓度,使用5 mmol/L阿魏酸对海巴戟叶片处理进行下一步筛选处理最佳时间条件的实验。
2.1.2 5 mmol/L阿魏酸处理海巴戟叶片48 h内莨菪亭含量的变化 使用5 mmol/L阿魏酸处理海巴戟叶片测定其0~48 h期间莨菪亭的含量变化,结果如图1B,经过最佳浓度的阿魏酸处理叶片后,随着时间的增加其莨菪亭含量有一定的变化规律,期间莨菪亭含量先升高后下降。处理叶片后2 h便有明显的反应,其莨菪亭含量开始显著增加,到8 h为最高峰,含量增加到最大值0.78 mg/g,随后其莨菪亭含量逐渐减少,叶片中的莨菪亭含量到48 h降低至最低值0.36 mg/g,并且显著比对照值低。
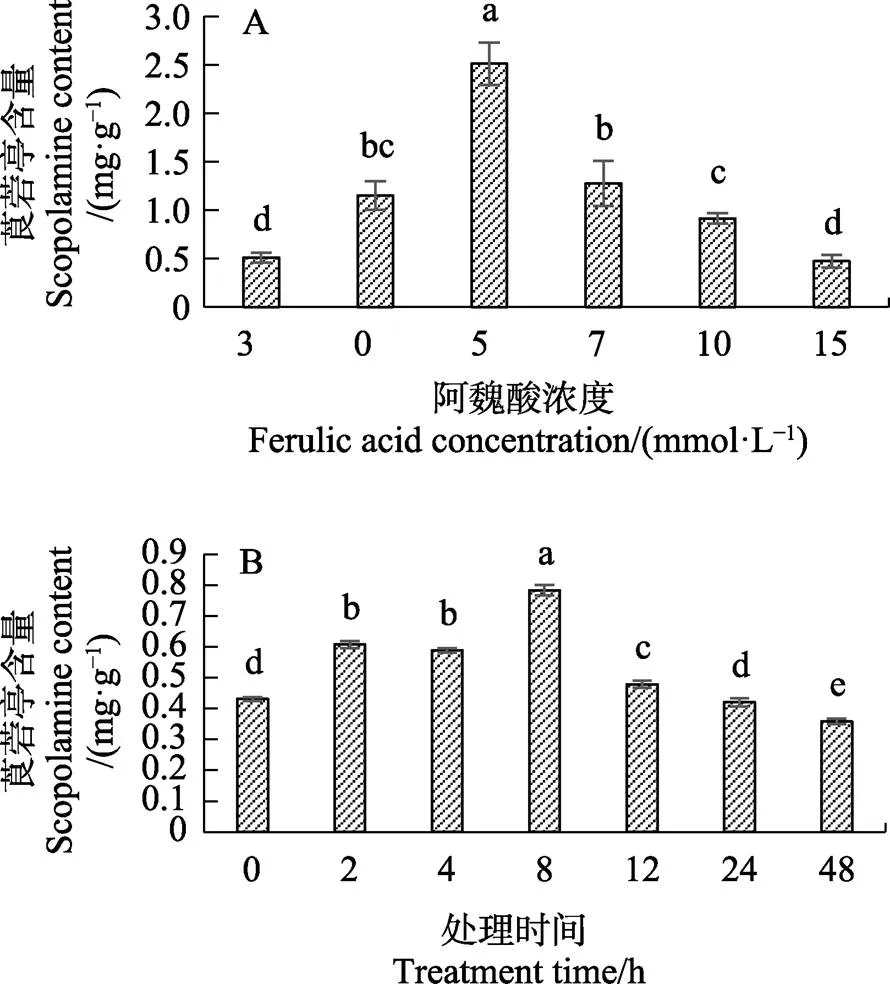
不同小写字母表示处理间差异显著(P<0.05)。
2.2 不同浓度的阿魏酸处理海巴戟后4CL总酶活性变化
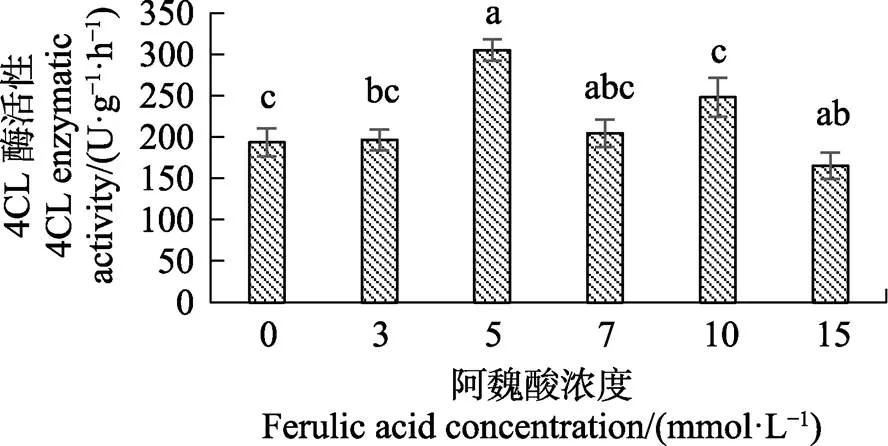
不同浓度的阿魏酸处理海巴戟叶片后,5 mmol/L和10 mmol/L阿魏酸处理显著高于对照,其中5 mmol/L阿魏酸处理使叶片中4CL总酶活性为最大值305 U/g×h,显著高于对照的酶活值193 U/g·h,而15 mmol/L阿魏酸处理的叶片中4CL总酶活性与对照相比显著降低(图2)。
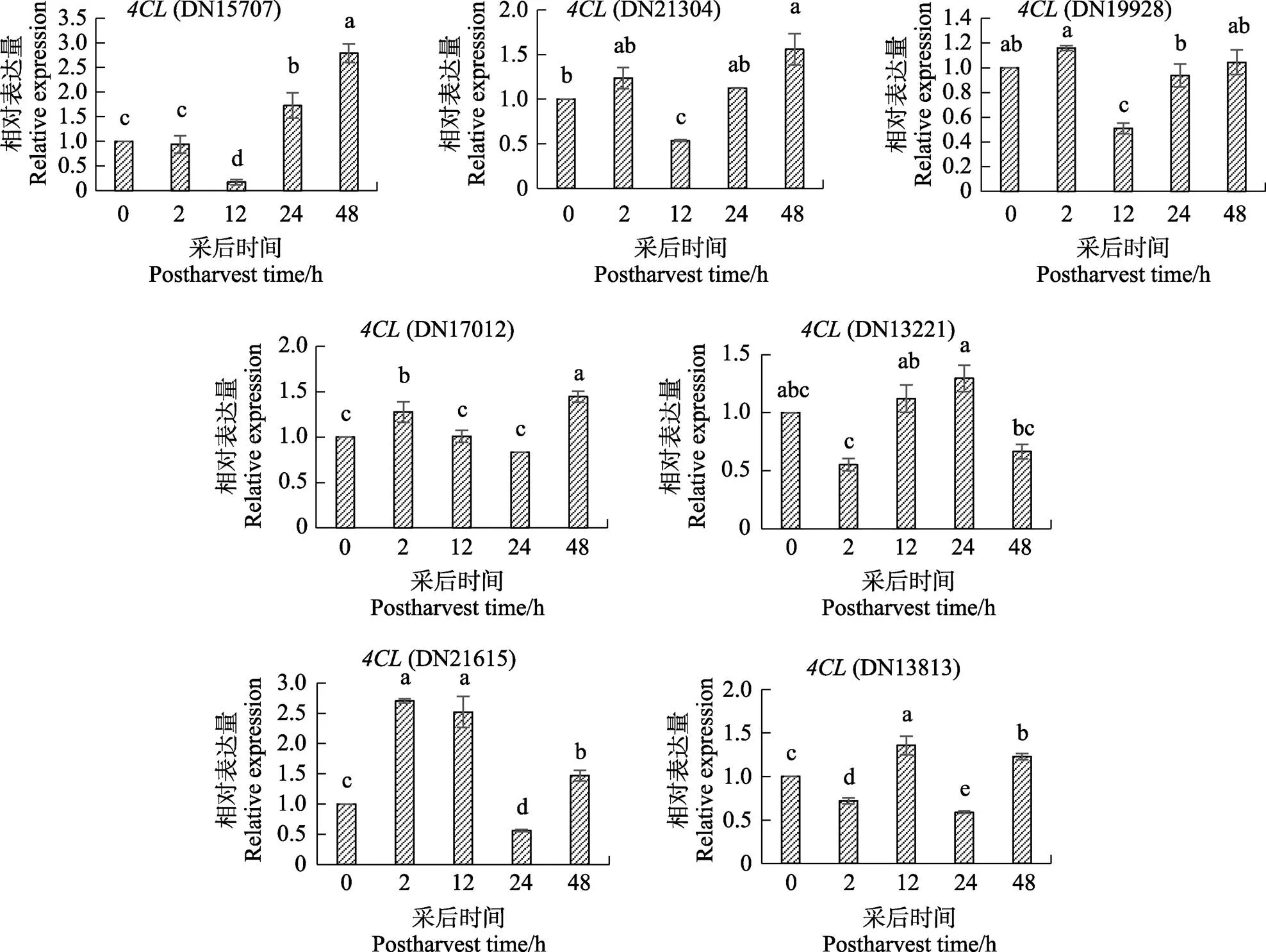
2.3 4CL基因在叶片中48 h内的表达分析
在海巴戟叶片采后0~48 h过程中,基因的表达情况各异(图3)。(DN21615)、(DN13813)、(DN17012)、(DN15707)、(DN21304)的表达量为上调,并且(DN21615)和(DN17012)的表达量在前期迅速反应与0 h相比显著上升;其中(DN21615)在2 h和12 h的表达量是0 h的表达量的2.5倍左右,而且(DN17012)表达量在后熟过程中并未比0 h表达量显著下降。(DN13221)表达量为下调,虽12 h和24 h表达量有所上升,但是上升幅度不大,基本与0 h没有太大的差异,并且最终在48 h表达量又显著下降了。(DN13813)和(DN19928)的表达量与0 h相比始终没有大幅度上调的显著差异,所以选择(DN21615)、(DN17012)、(DN15707)、(DN21304)进行后续最佳浓度阿魏酸处理不同时间叶片的荧光定量表达实验。
不同小写字母表示处理间差异显著(P<0.05)。
2.4 4CL基因在最佳浓度阿魏酸处理叶片48 h内的表达分析
在最佳浓度5 mmol/L阿魏酸的处理下,海巴戟叶片中基因在48 h内的表达情况见图4。(DN21615)的表达量在前期并没有迅速上升,反而相对空白对照其表达量显著下降,仅仅在12 h和24 h显著上升,其中24 h的表达量显著增加了1倍,但是最终其表达量在48 h又显著下降并且显著低于对照。(DN17012)仅仅在4 h表达量显著上升了,但是其余时间点如2、8、12 h其表达量与对照相比是下降的,后期表达量又恢复平稳与对照无显著差异。(DN15707)自前期开始其表达量是渐渐上升的,在8 h表达量显著增加,是对照表达量约3倍,而后表达量下降,自12 h开始逐渐降低,该基因的表达量变化趋势与其对应莨菪亭含量变化趋势较为一致。(DN21304)除了在2 h与对照保持平稳的表达量以外,从4 h开始表达量显著增加,而后逐渐降低后又稍有升高,但是总体自4 h到最终的48 h其表达量始终显著高于对照。
不同小写字母表示处理间差异显著(P<0.05)。
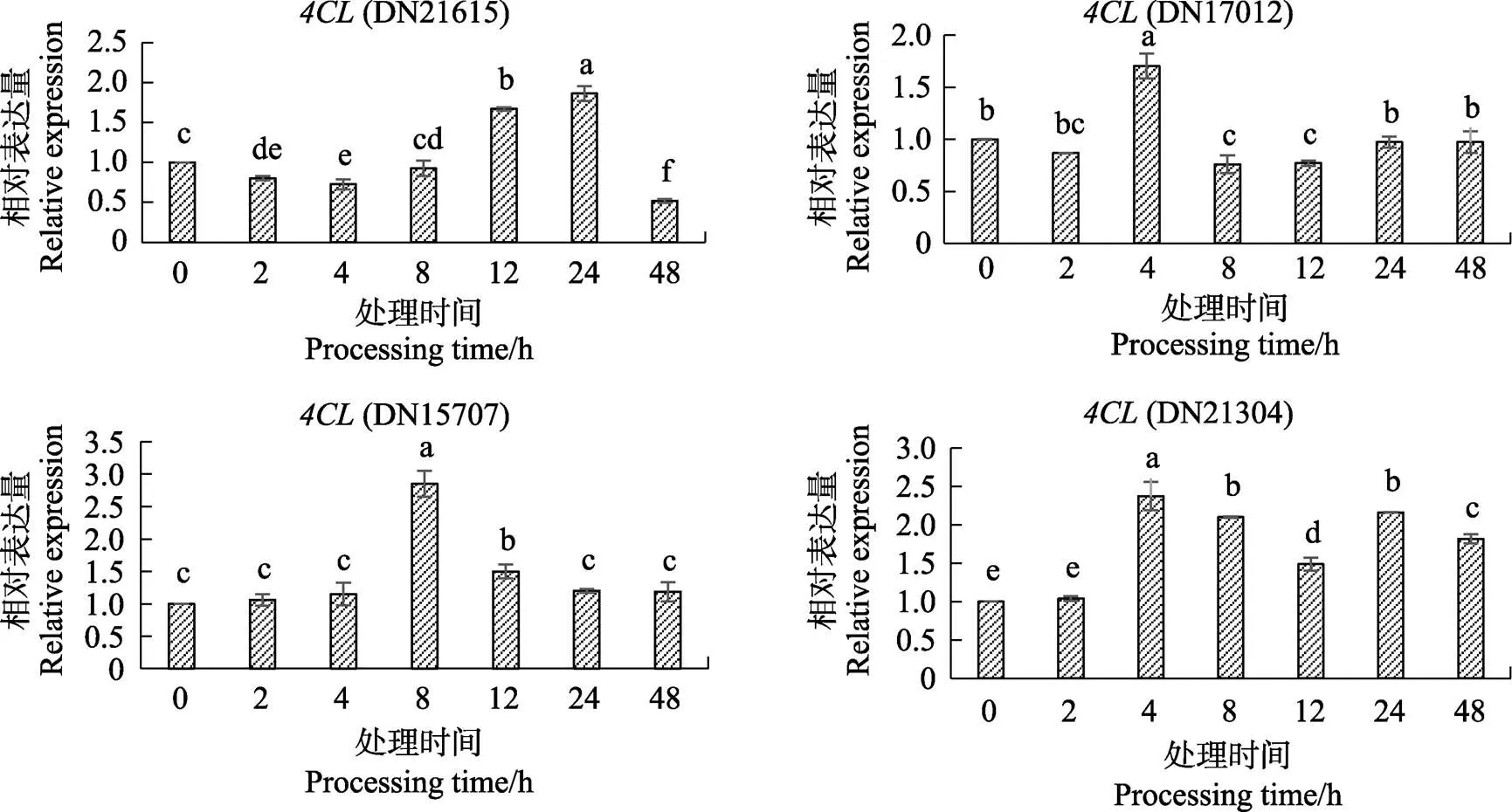
不同小写字母表示处理间差异显著(P<0.05)。
2.5 海巴戟4CL(DN15707)全长克隆结果
将(DN15707)的测序结果经过NCBI序列比对分析得知该序列与目标基因相符,(DN15707)长度为1632 bp,具有1632 bp完整开放阅读框,编码543个氨基酸。在NCBI进行序列比对,发现海巴戟(DN15707)与咖啡()、茶树()、橡胶树()和麻疯树()等植物的基因具有较高的相似性和一致性,初步确定该基因为4CL基因家族中的基因。系统进化树分析结果显示(图5),海巴戟(DN15707)与咖啡聚为一支,说明该基因与同为茜草科的咖啡基因亲缘关系较近,可能与咖啡基因具有相似的功能和结构。根据进化树进一步确定该基因为基因,将其命名为。将序列提交至GenBank,登录号为MZ676989。
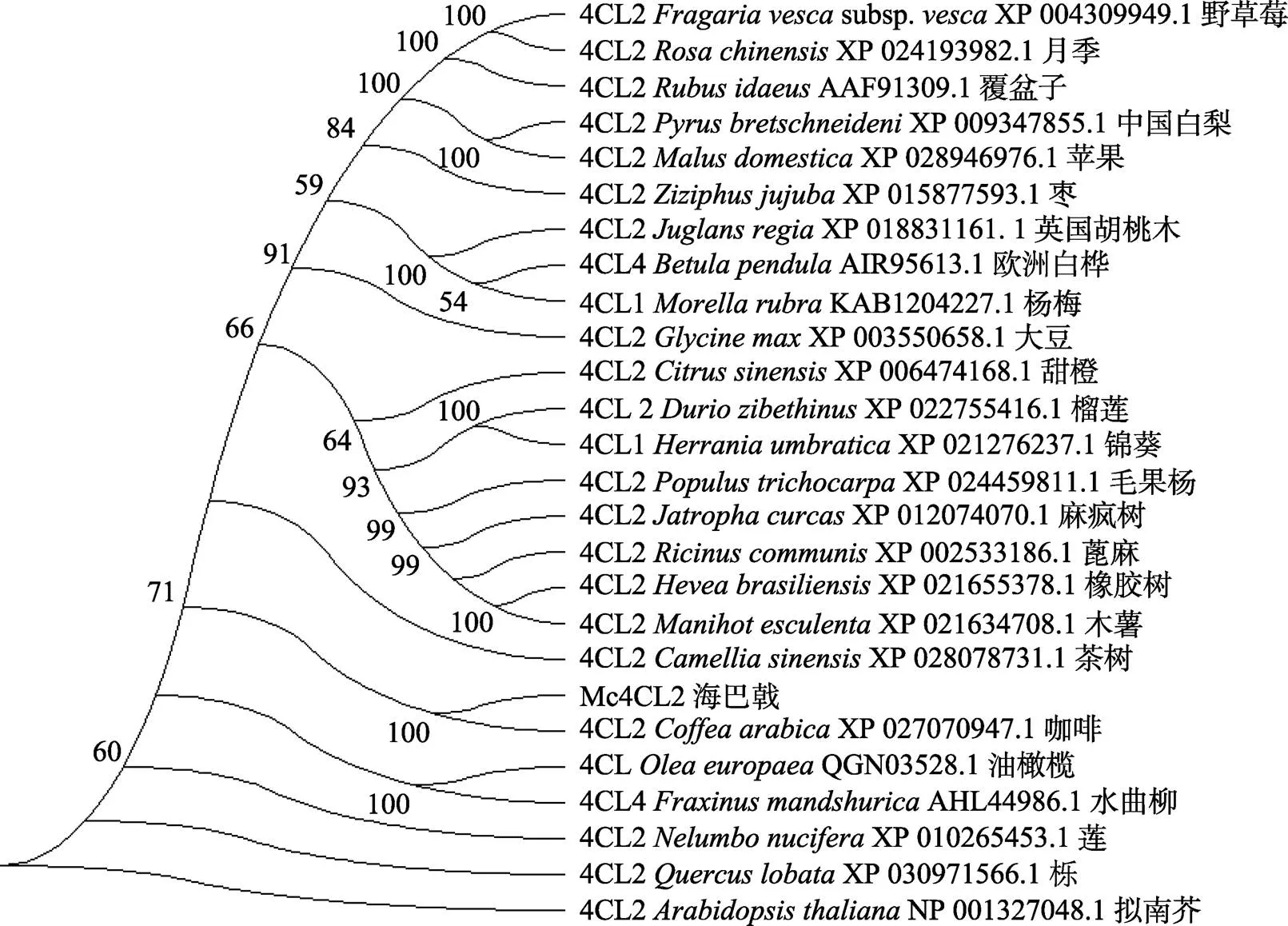
图5 Mc4CL2系统进化树分析
Mc4CL2分别在186~197、386~392氨基酸处具有4CL的特征序列LPYSSGTTGLPK(保守特征)和GEICIRG(催化功能特征),并且其保守功能结构域包括CaiC、AMP-binding、PLN02246、4CL以及pimA结构域。其中PLN02246和4CL是典型的4香豆酸辅酶A连接酶蛋白的标签;CaiC是酰基辅酶A合成酶,主要进行脂质运输与代谢、次级代谢产物生物合成、运输与分解代谢等作用;PimA是二羧酸辅酶A连接酶,属于酰基辅酶A连接酶大家族的成员之一;AMP-binding是AMP结合酶,主要通过依赖ATP的AMP与底物的共价结合而发挥作用,这些预测的结构功能与4CL参与次级代谢催化反应中需要结合AMP最终生成酰基辅酶A酯的特点相符,另外,Mc4CL2具有组成保守特征的活性位点、CoA结合位点、AMP结合位点以及对于ATP和AMP的结合是绝对必要的酰基激活酶(AAE)共识基序,以上充分说明该基因属于4CL基因家族,具有其催化合成酰基辅酶A酯反应功能的特征。
3 讨论
阿魏酸通过影响不同酶的活性从而导致植物的生长发生变化,由此可以利用此特性来促进海巴戟合成莨菪亭,更重要的是,阿魏酸是合成莨菪亭代谢途中的重要中间物质,这样的外源中间体的添加可以有效地促进次生代谢终端产物的合成[23],所以将阿魏酸作为前体物质进行前体饲喂,是一种促进海巴戟合成莨菪亭的有效手段。随着处理时间的增加阿魏酸的刺激作用也会有变化,海巴戟叶片随着阿魏酸处理时间的增加,其莨菪亭合成量增加的效果逐渐明显,但是超过8 h后其效果会逐渐降低,说明阿魏酸诱导海巴戟增加莨菪亭含量的效果可能具有时效性。阿魏酸处理叶片48 h其莨菪亭含量比对照值低,该时间点可能是阿魏酸对莨菪亭诱导作用的失效节点。然而,只是单一的莨菪亭含量变化的现象无法充分证明是因为阿魏酸的作用效果变化导致的,可能有离体材料自身的代谢活动或者其他原因导致的,阿魏酸对海巴戟合成莨菪亭的诱导作用机制需要联系酶活性、基因表达、代谢反应等相关方面做深入的研究。
阿魏酸处理对不同植物的酶活有不同的影响,宛国伟等[24]使用不同浓度的阿魏酸浸泡丹参()根部,对苯丙氨酸解氨酶的活性均产生抑制作用但是却对多酚氧化酶的活性均有促进作用。用阿魏酸处理甘蔗(),对苯丙氨酸解氨酶的活性有促进作用,却对多酚氧化酶的活性产生抑制作用[25]。同时,不同浓度的阿魏酸刺激植物调控其体内代谢活动的强弱程度是不同的,使用10 mmol/L阿魏酸浸泡番茄()促进其酶活性提高、抗病性能增强的效果比5 mmol/L阿魏酸的效果好[26]。不同浓度的阿魏酸处理海巴戟叶片后,发现阿魏酸处理可以提高4CL总酶活,其中5 mmol/L的浓度能最大限度地提高,而15 mmol/L阿魏酸处理叶片会降低其4CL总酶活性。本研究中在一定浓度下阿魏酸对海巴戟中4CL酶活性表现为促进作用,可能主要是因为阿魏酸是4CL的反应底物,当底物增多时就会促使4CL转化更多的阿魏酸进行催化反应。另外,本研究是以阿魏酸为底物进行4CL总酶活性的测定,由于4CL催化阿魏酸生成阿魏酰辅酶A是4CL催化反应中最靠近生成莨菪亭终端步骤的一个转化反应,所以能较准确地反映出4CL总酶活性变化与阿魏酸诱导促进莨菪亭积累之间的作用关系。根据本研究中阿魏酸处理能诱导莨菪亭合成量增加、4CL总酶活性增强,并且在5 mmol/L阿魏酸处理条件下诱导二者效果最佳的一致性结果来看,可说明4CL活性的提高在合成莨菪亭途径中有着正向的促进作用。
通过阿魏酸外源处理海巴戟叶片,可以诱导基因不同程度的表达变化。其中(DN15707)和(DN21304)的表达量均比对照高,说明阿魏酸可以诱导(DN15707)和(DN21304)高度表达,可能是因为(DN15707)和(DN21304)对阿魏酸作为底物的亲和力要高一些,所以受到阿魏酸的刺激其表达量较为突出,但是在12~48 h莨菪亭含量是下降的,而(DN21304)在此期间的表达量始终显著高于对照,整体来看(DN15707)的表达趋势与其莨菪亭含量变化趋势较为一致,因此(DN15707)在合成莨菪亭途径中的作用较为重要。鉴于4CL在莨菪亭合成途径中的作用点较多,具体(DN15707)参与合成莨菪亭的哪个环节还需要后续进一步探讨。在合成莨菪亭过程中具体的作用底物还需要结合其纯化蛋白利用不同底物的酶活性、超表达或沉默该基因等实验进一步验证和研究。
本研究克隆所得的基因可能是莨菪亭累积过程中的关键基因,它能启动4CL酶活,充分利用阿魏酸底物进而导致莨菪亭累积。
[1] MOTSHAKERI M, GHAZALI H M. Nutritional, phytochemical and commercial quality of noni fruit: a multi-beneficial gift from nature[J]. Trends in Food Science and Technology, 2015, 45(1): 118-129.
[2] 杨小波. 海南植物名录[M]. 北京: 科学出版社, 2013.
YANG X B. Plant list of Hainan[M]. Beijing: Science Press, 2013. (in Chinese)
[3] 刘德兵, 魏军亚, 王樱莅, 李绍鹏. 海南省野生及栽培果树资源(续)[J]. 中国南方果树, 2008, 37(6): 26-28.
LIU D B, WEI J Y, WANG Y L, LI S P. Wild and cultivated fruit tree resources in Hainan Province (continued)[J]. South China Fruits, 2008, 37(6): 26-28. (in Chinese)
[4] OSMAN N F W, TANTOWI N A C A, LAU S F, MOHAME S. Epicatechin and scopoletin rich(Noni) leaf extract supplementation, mitigated osteoarthritis via anti-inflammatory, anti-oxidative, and anti-protease pathways[J]. Journal of Food Biochemistry, 2019, 43(3): e12755.
[5] YUAN C, WANG M H, WANG F, CHEN P Y, KE X G. Network pharmacology and molecular docking reveal the mechanism of scopoletin against non-small cell lung cancer[J]. Life Sciences, 2021, 270: 119105.
[6] AHMADI N, MOHAMED S, SULAIMAN RAHMAN H, ROSLI R. Epicatechin and scopoletin-richleaf ameliorated leukemia via anti-inflammatory, anti-angiogenesis, and apoptosis pathwaysand[J]. Journal of Food Biochemistry, 2019, 43(7): e12868.
[7] OSMAN W N W, MOHAMED S. StandardizedL. andL. leaf extracts alleviated fatigue by improving glycogen storage and lipid/carbohydrate metabolism[J]. Phytotherapy Research, 2018, 32(10): 2078-2085.
[8] PETER P I, PETER K V. Noni (L.): research and development[M]. Berlin: Springer, 2018.
[9] DENG S, WEST B J, JENSEN C J. A quantitative comparison of phytochemical components in global noni fruits and their commercial products[J]. Food Chemistry, 2010, 122(1): 267-270.
[10] PANDY V, NARASINGAM M, KUNASEGARAN T, MURUGAN D D, MOHAMED Z. Effect of noni (Linn.) fruit and its bioactive principles scopoletin and rutin on rat vas deferens contractility: an ex vivo study[J]. The Scientific World Journal, 2014, 2014:909586.
[11] 谢海娟, 范希德, 叶广继, 周 云, 王 舰. 马铃薯的克隆及表达分析[J]. 生物技术通报, 2019, 35(11): 1-8.
XIE H J, FAN X D, YE G J, ZHOU Y, WANG J. Cloning and expression analysis ofgene in[J]. Biotechnology Bulletin, 2019, 35(11): 1-8. (in Chinese)
[12] 董丽丽, 阿不都热扎克·依沙克, 户 倩, 李川微, 张水明. 石榴4-香豆酸辅酶A连接酶基因的克隆和表达分析[J]. 西北植物学报, 2016, 36(11): 2146-2151.
DONG L L, ABDURAZAK I, HU Q, LI C W, ZHANG S M. Cloning and expression analysis of pomegranate 4-coumarate-CoA ligase[J].Acta Botanica Boreali-Occidentalia Sinica, 2016, 36(11): 2146-2151. (in Chinese)
[13] 宋洁洁, 罗红梅, 朱珣之, 张 玉, 高 婷. 珊瑚菜基因克隆与生物信息学分析[J]. 世界科学技术-中医药现代化, 2017, 19(4): 610-617.
SONG J J, LUO H M, ZHU X Z, ZHANG Y, GAO T. Cloning and bioinformatics analysis ofgene in[J]. Modernization of Traditional Chinese Medicine and Materia Medica-World Science and Technology, 2017, 19(4): 610-617. (in Chinese)
[14] 张 蕾, 林 晓, 罗 赟, 汪佳易, 徐 川. RNAi沉默基因对草莓果实花色苷代谢的影响[J]. 果树学报, 2015, 32(3): 434-439, 522.
ZHANG L, LIN X, LUO Y, WANG J Y, XU C. Influences of RNAi-inducedsilencing on anthocyanin metabolism in strawberry fruit[J].Journal of Fruit Science, 2015, 32(3): 434-439, 522. (in Chinese)
[15] 袁婷婷, 董 坤, 郭增鹏, 董 艳. 阿魏酸诱导蚕豆枯萎病发生及根系组织结构损伤的化感效应[J]. 植物营养与肥料学报, 2020, 26(5): 914-923.
YUAN T T, DONG K, GUO Z P, DONG Y.Allelopathic effects of ferulic acid inducingwilt occurrence and abnormal root tissue structure of faba bean[J].Journal of Plant Nutrition and Fertilizers, 2020, 26(5): 914-923. (in Chinese)
[16] 金 晶, 付远洪, 李朝婵, 唐凤华. 阿魏酸对2种野生杜鹃种子萌发的化感作用[J]. 种子, 2017, 36(6): 22-25.
JIN J, FU Y H, LI C C, TANG F H. Allelopathy of ferulaic acid on seed germination of two wild rhododendron[J]. Seed, 2017, 36(6): 22-25. (in Chinese)
[17] 胡 斌, 潘 洁, 陈 丽, 郑小林, 李博强. 阿魏酸处理对番茄采后果实品质和青霉病害的影响[J]. 保鲜与加工, 2019, 19(1): 14-18, 24.
HU B, PAN J, CHEN L, ZHENG X L, LI B Q. Effect of ferulic acid treatment on postharvest quality and blue mold in tomato fruit[J]. Storage and Process, 2019, 19(1): 14-18, 24. (in Chinese)
[18] 沈宋利, 杨 健, 沈燕飞, 唐泽辉, 陈梅兰. 超声萃取高效液相色谱法测定诺丽果粉中的莨菪亭、芦丁和槲皮素含量[J]. 食品科学, 2017, 38(6): 191-196.
SHEN S L, YANG J, SHEN Y F, TANG Z H, CHEN M L. Determination of scopoletin, rutin, and quercetin in noni () fruit powder by ultrasonic extraction coupled with high performance liquid chromatography[J]. Food Science, 2017, 38(6): 191-196. (in Chinese)
[19] 岳 凯, 刘文瑜, 魏小红. 干旱胁迫对不同品系藜麦内黄酮和抗氧化性的影响[J]. 分子植物育种, 2019, 17(3): 956-962.
YUE K, LIU W H, WEI X H. Effect of flavonoids content and key enzymes and antioxidant activities of flavonoids in different quinoa on drought stress[J]. Molecular Plant Breeding, 2019, 17(3): 956-962. (in Chinese)
[20] WEI H, XU C, MOVAHEDI A, SUN W B, ZHUGE Q. Characterization, expression profiling, and functional analyses of a-like gene of[J]. Processes, 2019, 7(1): 45.
[21] 吴 田, 蓝增全, 王华芳. 诺丽基因片段克隆及实时荧光定量PCR方法的建立[J]. 中南林业科技大学学报, 2018, 38(2): 16-22.
WU T, LAN Z Q, WANG H F. Cloning and development of real-time fluorescence quantitative PCR assay ofgene fragment from noni[J].Journal of Central South University of Forestry & Technology, 2018, 38(2): 16-22. (in Chinese)
[22] 胡丹焱, 张正雪, 刘 娟, 蓝增全, 吴 田. 基于转录组测序的海巴戟果实中乙烯调控相关基因表达分析[J]. 四川农业大学学报, 2020, 38(5): 528-537.
HU D Y, ZHANG Z X, LIU J, LAN Z Q, WU T. Expression analysis of ethylene regulate related genes infruit based on transcriptome sequencing[J]. Journal of Sichuan Agricultural University, 2020, 38(5): 528-537. (in Chinese)
[23] 吕春茂, 范海延, 姜 河, 孟宪军. 植物细胞培养技术合成次生代谢物质研究进展[J]. 云南农业大学学报, 2007, (1): 1-7.
LV C M, FAN H Y, JIANG H, MENG X J. Research advances on synthesis of secondary metabolities by plant cell culture[J]. Journal of Yunnan Agricultural University (Natural Science), 2007, (1): 1-7. (in Chinese)
[24] 宛国伟, 董娟娥, 梁宗锁,李进瞳, 王武军. 培养条件对离体丹参根苯丙氨酸解氨酶和多酚氧化酶活性的影响[J]. 西北植物学报, 2007(12): 2471-2477.
WAN G W, DONG J E, LIANG Z S, LI J T, WANG W J. Phenylalanine ammonia lysae (PAL) and polyphenol oxidase (PPO) activities of in vitro root under different culturing conditions in[J]. Acta Botanica Boreali-Occidentalia Sinica, 2007(12): 2471-2477. (in Chinese)
[25] 大 东. 鲜切甘蔗和芋头抗褐变剂筛选及其抗褐变机理研究[D]. 武汉: 华中农业大学, 2019.
MAMOUN ADAM H D. Study on screening anti-browning agents and mechanism of inhibiting the browning on fresh-cut sugarcane and taro[D]. Wuhan: Huazhong Agricultural University, 2019. (in Chinese)
[26] 潘 洁. 阿魏酸处理对采后番茄果实品质影响及青霉病控制的研究[D]. 杭州: 浙江工商大学, 2017.
PAN J. Effects of post-harvest ferulic acid treatment on storage quality and control of blue mold caused byin tomato fruit[D]. Hangzhou: Zhejiang Gongshang University, 2017. (in Chinese)
Function of GeneDuring the Accumulation of Scopoletin inLeaves
GONG Shusen, YANG Fei, WANG Qingfen, YANG Ziyun, WU Tian*
College of Landscape Architecture and Horticulture Sciences, Southwest Forestry University / Southwest Landscape Architecture Engineering Research Center of State Forestry and Grassland Administration / Yunnan Engineering Research Center for Functional Flower Resources and Industrialization, Kunming, Yunnan 650224, China
In order to clarify the accumulation of the scopoletin inleaves, and to explore the role of genein the accumulation process, the content of scopoletin in the leaves ofwas determined within 0‒48 h by high performance liquid chromatography, and the changes of the scopoletin content after treatment with different concentrations of ferulic acid were determined. An AUV spectrophotometer was used to measure the total enzyme activity of 4CL using ferulic acid as a substrate for the enzyme activity reaction. The qPCR of sevengenes was performed based on the RNA-seq results, and onegene was screened out as a key candidate gene because its expression trend was consistent with the change trend of the scopoletin content and enzyme activity. Its full-length was cloned and bioinformatics analysis was performed, and finally its expressive characteristics was studied by qRT-PCR.leaves had the highest content of scopoletin at 12 h, reaching 0.35 mg/g, and the lowest at 48 h with 0.18 mg/g within 48 hours after picking. The change of 4CL total enzyme activity was similar to the trend of the scopoletin content. The total enzyme activity of 4CL in the leaves treated with ferulic acid increased. The(DN15707) gene was screened out by qPCR. The gene was 1632 bp in length, with a complete open reading frame. Sequence alignment and phylogenetic tree analysis revealed the gene was agene in thegene family, named asgene. Thegene may be a key gene in the accumulation of scopoletin. It can initiate the activity of 4CL enzyme, make full use of ferulic acid substrate and lead to the accumulation of scopoletin.
; scopoletin; total enzyme activity; 4-coumarate-Co A ligase (4CL); ferulic acid
S567.19
A
10.3969/j.issn.1000-2561.2022.10.004
2022-01-02;
2022-03-05
国家林业和草原局项目(No. [2019]27);云南省科技厅重点研发计划项目(No. 2019IB011)。
宫树森(1997—),男,硕士研究生,研究方向:植物分子生物学。*通信作者(Corresponding author):吴 田(WU Tian),E-mail:wutianpotato@swfu.edu.cn。
