微重力对拟南芥长势影响的代谢网络流平衡分析*
2022-11-09王芷涵张青叶张红雨
王芷涵 阮 耀 张青叶 张红雨
(华中农业大学信息学院 武汉 430070)
0 引言
为保证长期空间任务的顺利进行,必须有良好的生物再生生命保障系统(Bioregenative Life Support System, BLSS)。该系统能够将代谢废物变为可供利用的生物量,是人类长期空间生存的基础[1]。在该系统中,高等植物起着关键作用。一方面,植物可通过光合作用合成人类生存所需营养物质,如糖类、脂类、蛋白质、膳食纤维和维生素等;另一方面,光合作用还可以消耗航天员呼吸产生的二氧化碳,生成氧气,起到调节飞行器中二氧化碳与氧气浓度平衡的作用。此外,高等植物还可以为密闭、狭小和嘈杂的空间在轨系统提供勃勃生机和活力,起到缓解航天员紧张情绪的作用[2]。因此,了解空间环境对植物生长发育的影响是研发BLSS 必不可少的工作。航天器中的微重力环境对植物而言是一种生长逆境,微重力会影响植物细胞分裂、细胞壁合成、胚胎发生、蛋白质、淀粉及脂肪酸代谢的调节、次生代谢物等植物生理生化特征的各个方面和生命活动的许多关键环节[3]。同时,在微重力条件下,植物可以节省在地面上必须维持重力的位置动态平衡而被消耗的能量,这可能会影响ATP 的产生[4]。
植物在微重力环境下可进行快速代谢反应,瞬时微重力环境下的拟南芥被观察到对能量的需求增加,并改变了许多基因的转录水平,其中主要是与钙离子和活性氧相关的基因[5]。在长期的微重力条件下,植物会通过代谢变化适应环境的改变。在空间飞行14 天后,拟南芥愈伤组织中有关碳水化合物代谢途径的酶表达水平被观察到较大的上调,说明拟南芥借此保证处于微重力环境期间的细胞能量供应[6]。微重力除改变植物的能量代谢之外,还影响着植物的转录后翻译、细胞壁合成、生长素的运输以及抗氧化防御等过程[7]。
微重力环境对植物的影响是多方面的,而代谢分析是能够较为全面反映细胞过程的研究手段。作为系统级的生物网络分析方法,基因组规模的代谢网络(Genome-scale Metabolic Network Model)可根据化学计量平衡原理,基于基因-蛋白-反应三者关联,以化学计量矩阵的形式对包含细胞内部发生的全部已知生化反应建立数学模型,然后使用约束重建和分析 (COBRA) 方法[8]对模型进行约束分析,模拟生物体的代谢状态。同时,代谢网络还允许将不同的组学信息整合到网络中,建立特异性代谢模型,提高模型预测的准确性[9]。目前已测量了空间环境下拟南芥等高等植物的基因表达数据[10,11],本研究拟将基因表达数据整合到代谢网络中,构建地面环境和微重力环境下拟南芥的特异性代谢网络,结合流平衡分析(Flux Balance Analysis, FBA)等代谢网络优化算法,获得地面和微重力环境下拟南芥的代谢状态,研究微重力环境对植物代谢过程的影响机制。
1 材料与方法
1.1 基因表达数据
拟南芥的基因表达数据从GEO 数据库下载,注册号分别为GSE56658[10],GSE56659[11]。二者均为STS-131 任务期间在空间中培育的拟南芥基因芯片数据,控制变量为微重力大小。前者是在BRIC-16 飞行试验绕地飞行近13 天后测量的未分化的拟南芥细胞和拟南芥黄化幼苗的基因芯片数据。其中,空间飞行与地面控制条件各6 个样品,共24 组数据,这里选取其中拟南芥黄化幼苗的基因芯片数据进行分析。后者是TAGES 飞行试验期间,在国际空间站(ISS)生长了12 天后测量的拟南芥幼苗的芽、根、下胚轴以及整个植株的基因芯片数据,并使用相同条件在地面做了对比试验。全植株重复3 组,芽、根、下胚轴各重复5 组,共36 组数据。
为研究拟南芥黄化幼苗、幼苗以及拟南芥不同器官组织对微重力的代谢响应,收集整理了拟南芥黄化幼苗、幼苗、芽、根、下胚轴在微重力和地面环境下的转录组数据。在R(版本3.6.2)中使用R 包GEOquery(版本2.54.1)从GEO 数据库下载基因芯片数据,使用R 包affy(版本1.64.0)、affydata(版本1.34.0)和limma(版本3.42.2)对数据进行质控,使用RMA 方法[12]进行芯片数据标准化,参数设置为默认。为了将基因表达数据整合到代谢网络中,需将探针名称替换为代谢模型中的基因名。由于存在多个探针对应同一个基因的情况,对存在多个表达值的基因,为将其整合到代谢网络中并使其最接近真实转录值,将重复基因表达数据的均值作为网络的约束。
1.2 基因组规模代谢网络模型
AraGEM[13]是基因组规模的拟南芥代谢网络模型。该模型主要涵盖拟南芥的初级代谢,还包括了一部分次级代谢,共1601 个反应、1737 个代谢产物和4833 个基因-酶-反应的关联,并根据亚细胞定位分为胶质细胞、线粒体、质体和过氧化物酶体。该模型包含预设的约束条件,为将基因表达信息作为约束整合到代谢网络中,这里使用移除这些约束条件后的模型[14]。
1.3 基因表达数据约束代谢网络模型
利用代谢网络分析工具COBRApy[15]将基因表达数据整合到代谢网络中。已开发Python 库 py-TARG[16]**https://github.com/SergioBordel/pyTARG,通过对模型中所有反应施加最大边界,实现自动约束人类全基因组代谢网络。该方法允许计算代谢通量分布并评估限制酶催化反应速率的结果。因此,已被用于识别癌细胞系的代谢药物靶点[17]。该方法基于基因-蛋白-反应的关联规则,把基因表达数据作为模型中该基因对应反应的上限约束。为避免求解线性规划时的数值问题,按常规的处理方式取基因表达值的10 倍整数[17]作为该基因对应反应的上限。分别将拟南芥黄化幼苗、幼苗、芽、根和下胚轴在空间微重力与地面条件下的基因表达数据整合到拟南芥代谢网络模型AraGEM 中,得到不同植株和组织处于微重力环境和地面环境下的特异性拟南芥代谢网络模型。
1.4 流平衡分析
流平衡分析(Flux Balance Analysis,FBA)[18]是一个基于约束的模拟方法,通过计算代谢物通过该代谢网络的通量,预测生物体的生长速率或代谢物的产生速率。流平衡分析将代谢物与反应的关系表示为化学计量矩阵,求解由约束和目标函数组成的线性规划问题,得到目标函数的最优解和整个网络的代谢通量分布。在数学上,FBA 可表示为

其中,Z是代谢网络的目标函数,通常为最大化生物质产量,S代表化学计量矩阵,v表示反应通量,vlower和vupper分别为反应的下限和上限。等式约束确保了模型中代谢物产生的总量及其消耗的总量在稳态时是相等的,不等式约束规定了代谢反应的不可逆性以及允许的最大和最小通量。
这里以生物量为目标函数,计算拟南芥黄化幼苗、幼苗、芽、根、下胚轴在空间微重力与地面条件下的生物质产量、ATP 产量和代谢通量分布,并计算了生物质产量和ATP 产量的下降率,下降率为负值时代表产量上升。生物质产量代表拟南芥长势,生物质产量越高说明拟南芥长势越好。
1.5 KEGG 富集分析
为进一步研究微重力环境影响拟南芥代谢状态的生理机制,在流平衡分析得到拟南芥的生物质产量和代谢通量分布后,微重力与地面环境下拟南芥黄化幼苗、幼苗、芽、根、下胚轴的差异代谢反应所对应的基因被提交到DAVID 数据库[19]中进行KEGG 富集分析。KEGG 富集分析是生物信息研究中常用的方法,常用于研究测序数据中得到的一系列基因属于哪些通路。常用的富集分析由超几何分布计算,其数学表达式如下:其中,N代表总样本数;M代表总样本中特定类别的数量;n代表从总样本中随机抽取的数量;k代表抽取样本中属于特定类别的数量;p表示当原假设为真时得到的样本观察结果或更极端结果出现的概率,p值越小说明得到的结果越具有显著性,这里选择p值小于0.01 的通路作为富集结果。

2 结果
2.1 生物质产量以及ATP 相关反应的通量分析
对拟南芥黄化幼苗、幼苗、芽、根、下胚轴对应样本的基因表达值取平均,分别整合到拟南芥代谢网络模型AraGEM 中,得到拟南芥黄化幼苗、幼苗、芽、根、下胚轴的特异性代谢模型,利用流平衡分析得到其在微重力与地面环境下的生物质产量和代谢物产量。除了生物质外,ATP 的产量也被用来比较拟南芥的生长水平。在拟南芥的基因组规模代谢模型中,REner01_p 是叶绿体中产生ATP 的整体反应,RN04_m 是线粒体中NADH 呼吸链的ATP 反应产量,RN05_m 是线粒体中FADH 呼吸链的ATP 反应产量。
图1 为基于拟南芥黄化幼苗、幼苗、芽、根、下胚轴的代谢网络预测得到的生物质及叶绿体、线粒体NADH 呼吸链和FADH 呼吸链的ATP 产量,微重力环境下拟南芥黄化幼苗、幼苗、芽、根、下胚轴的生物质产量对比地面环境组均有所下降,分别下降了33.00%,51.52%,6.89%,12.53% 和11.70%。由于幼苗的叶绿体数量大于任何局部组织,拟南芥幼苗叶绿体的ATP 产量最高。作为叶绿体的主要分布部位,芽叶绿体的ATP 产量略低于幼苗。根与下胚轴不存在叶绿体,整合其转录组数据的拟南芥代谢模型叶绿体的ATP 产量位于同一水平,大大低于幼苗和芽叶绿体的ATP 产量。同时,由于拟南芥黄化幼苗叶绿体的生物发生受到抑制[20],拟南芥黄化幼苗叶绿体的ATP 产量远低于拟南芥幼苗。已有研究证明,微重力会使植物的叶绿素含量降低[21],因此微重力环境下拟南芥幼苗叶绿体的ATP 产量相比地面环境的下降程度远大于拟南芥黄化幼苗,微重力环境对拟南芥黄化幼苗叶绿体的ATP 产量影响相对更小。而微重力对线粒体电子传递链在拟南芥黄化幼苗、拟南芥幼苗以及拟南芥的芽、根、下胚轴中的影响大不相同,相比地面环境,拟南芥幼苗和黄化幼苗线粒体中FDAH 呼吸链在空间环境下的ATP 产量均有所提高,这可能是由于拟南芥节省了在地面上必须维持重力的位置动态平衡而消耗掉的能量,而根与下胚轴的情况却与之相反,这可能是由于拟南芥的根与下胚轴相对其他部位来说受到了更大、更复杂的微重力环境影响,根系在微重力环境和地面环境下表型相差不大的拟南芥应该更适应空间环境。从整个植株来看,虽然微重力条件下拟南芥黄化幼苗和幼苗的生物质产量均大幅度下降,但相比拟南芥幼苗,拟南芥黄化幼苗的生物质产量的降幅更小,因此拟南芥黄化幼苗更适应空间微重力环境,选择拟南芥黄化幼苗以及根与下胚轴短小的幼苗可能对构建生命保障系统更有帮助。

图1 拟南芥黄化幼苗(a)、幼苗(b)、芽(c)、根(d)和下胚轴(e)在微重力与地面环境下的生物质产量以及ATP 相关反应的通量对比Fig. 1 Mustard biomass production and ATP synthesis reaction fluxes of Arabidopsis thaliana etiolated seedlings(a), seedlings (b), shoots (c), roots (d), and hypocotyls (e) in microgravity and ground environment
目前已有研究表明,空间微重力条件下生长的拟南芥幼苗比地面生长的拟南芥幼苗矮小,在空间微重力条件下生长的瓦斯莱型(Ws)和哥伦比亚型(Col)型拟南芥比地面条件下短约3 mm[22]。从拟南芥各植株或组织来看,微重力环境对拟南芥根的长势影响最大,目前已有文献表明,相比地面条件,空间微重力条件下拟南芥的生长速度更慢。在空间微重力条件下Ws 型和Col 型拟南芥均表现出较短的芽和根;对于根,空间条件下生长的Ws 型拟南芥平均比地面条件下生长的拟南芥短5 mm,而Col 型拟南芥平均短14 mm[22]。
2.2 KEGG 富集分析
为了研究微重力与地面环境不同导致拟南芥生长差异的生理机制,需进一步比较两种环境中的主要差异代谢反应。由于大部分差异反应位于细胞质中,对位于细胞质上下调反应的相关基因进行KEGG 富集分析。
表1~5 分别是相对于地面环境、微重力环境中的拟南芥黄化幼苗、幼苗、芽、根和下胚轴下调反应对应的基因以0.01 为p值的阈值富集到最显著的10 个KEGG 通路。碳代谢、代谢途径、次生代谢产物的生物合成,光合生物中的碳固定,抗生素的生物合成,氨基酸的生物合成途径均出现在拟南芥黄化幼苗、拟南芥幼苗,拟南芥芽、根、下胚轴的富集结果中;乙醛酸和二羧酸酯代谢,丙酮酸代谢,柠檬酸盐循环(TCA 循环),精氨酸生物合成,氮素代谢,丙氨酸、天冬氨酸和谷氨酸代谢途径同时出现在拟南芥黄化幼苗和拟南芥幼苗的富集结果中;磷酸戊糖途径、糖酵解、果糖和甘露糖代谢、酪氨酸代谢同时出现在拟南芥芽、根、下胚轴的富集结果中,结果表明下调的通路在拟南芥黄化幼苗、幼苗和各器官组织中具有较高的一致性,即微重力环境对植物及植物不同部位的不利影响是相似的。详细结果列于表1~5。
相比地面环境,表6~9 分别列出了微重力环境中的拟南芥黄化幼苗、幼苗、根和下胚轴上调反应对应的基因被显著富集到的前10 个KEGG 功能通路。表10 是拟南芥芽上调反应对应的基因显著富集到的通路,相比拟南芥黄化幼苗、幼苗和其他器官组织,上调反应对应的基因富集到的通路较少,p值大于0.01 的通路仅有5 个,主要包含氧化磷酸化、吞噬体和抗坏血酸代谢等相关的代谢通路,说明拟南芥芽主要通过上调这些通路以应对微重力环境。而碳代谢、抗生素的生物合成、磷酸戊糖途径、光合生物中的碳固定、氨基酸的生物合成和糖酵解出现在拟南芥黄化幼苗和拟南芥幼苗的富集结果中,没有同时出现在拟南芥黄化幼苗,拟南芥幼苗,拟南芥芽、根、下胚轴的代谢途径,可能跟拟南芥芽的上调富集结果过少有关,这也说明相对于下调的通路,拟南芥黄化幼苗、幼苗和各器官组织上调通路的不一致性更高,即植物及植物不同部位采取不同策略应对微重力的影响。

表1 拟南芥黄化幼苗下调反应对应基因的富集途径Table 1 Enrichment pathways for genes corresponding to Arabidopsis thaliana etiolated seedlings down-regulation reactions
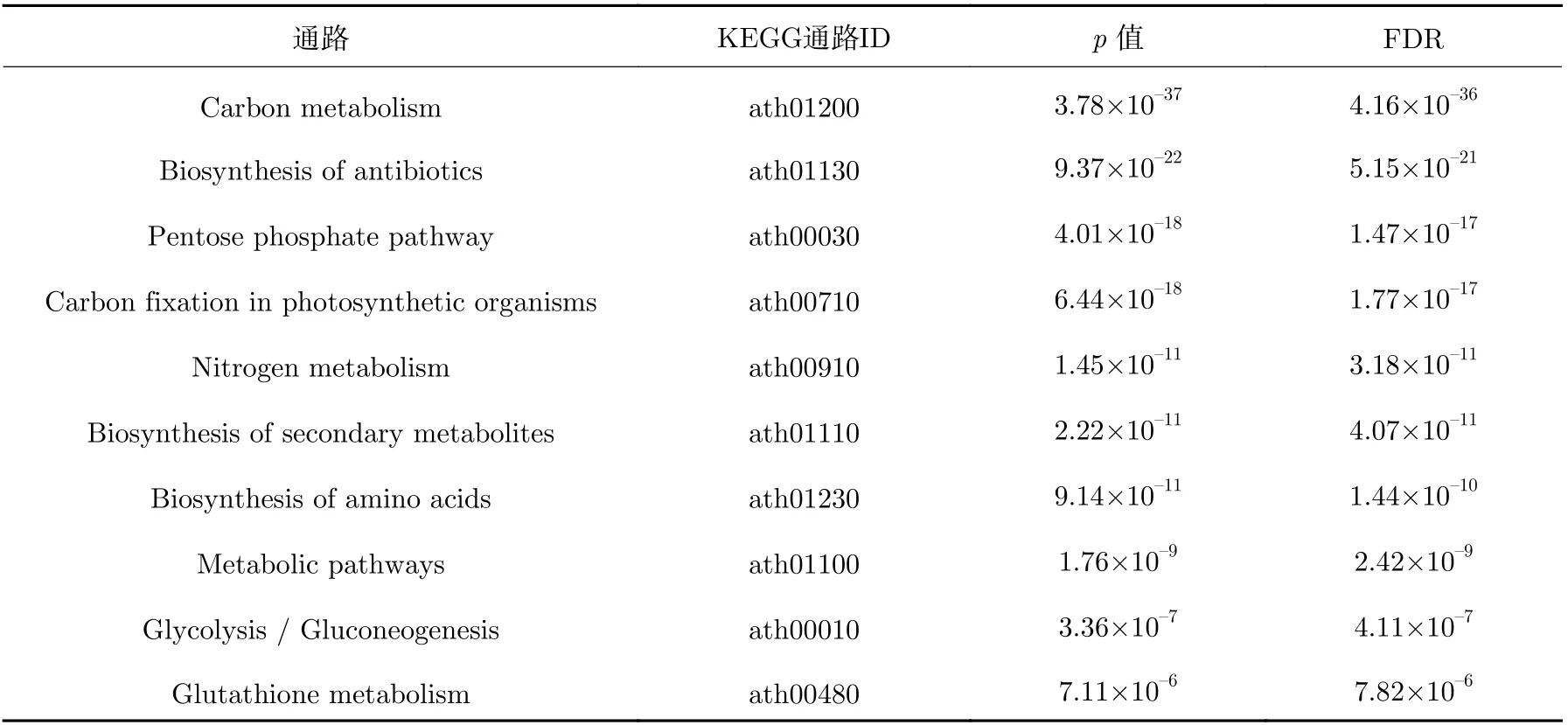
表6 拟南芥黄化幼苗上调反应对应基因的富集途径Table 6 Enrichment pathways for genes corresponding to Arabidopsis thaliana etiolated seedlings up-regulation reactions

表10 拟南芥芽上调反应对应基因的富集途径Table 10 Enrichment pathways for genes corresponding to Arabidopsis thaliana shoot up-regulation reactions

表2 拟南芥幼苗下调反应对应基因的富集途径Table 2 Enrichment pathways for genes corresponding to Arabidopsis thaliana seedlings down-regulation reactions
同时,富集结果表明,部分上调和下调反应对应的基因同时富集到同一通路中,例如柠檬酸循环出现在拟南芥幼苗的上调和下调反应的富集结果中,糖酵解出现在拟南芥根的上调和下调的富集结果中,说明这些通路在微重力环境下的代谢调控机制更加复杂。
柠檬酸循环是活细胞将有机物完全氧化为二氧化碳和水的主要途径之一。该循环提供了生命体活动的大部分能量,还提供了氨基酸、葡萄糖等合成所需的多种中间体[23]。分析拟南芥幼苗的上下调反应,发现微重力会下调拟南芥幼苗中柠檬酸循环过程中苹果酸生成草酰乙酸的反应和琥珀酸生成延胡索酸的反应,而上调柠檬酸生成草酰乙酸的反应。

表3 拟南芥芽下调反应对应基因的富集途径Table 3 Enrichment pathways for genes corresponding to Arabidopsis thaliana shoot down-regulation reactions

表4 拟南芥根下调反应对应基因的富集途径Table 4 Enrichment pathways for genes corresponding to Arabidopsis thaliana root down-regulation reactions
糖类最主要的生理功能是为机体提供生命活动需要的能量,糖酵解是所有生物体进行葡萄糖分解代谢必须经过的共同阶段。分析拟南芥根的上下调反应,发现微重力会下调β-葡萄糖的磷酸化反应和二磷酸果糖生成磷酸果糖的反应,上调α-葡萄糖磷酸化的反应。此前的研究也表明,糖酵解中氨基酸和糖代谢的异常会导致根发育的停滞[24]。
根据富集结果,光合作用中的碳固定通路被下调的情况也被发现。光合作用是绿色植物捕获光能并把二氧化碳和水合成有机物同时释放氧气的过程。其主要包括光反应、暗反应两个阶段,涉及光吸收、电子传递、光合磷酸化、碳同化等重要反应步骤。同时,光合作用是植物将无机物转换为人体能利用的有机物的重要途径,是长期空间任务中生物再生生命保障系统中不可缺少的组成部分。已有的研究证明,微重力会对植物的光合作用产生影响。例如在微重力环境中生长的小球藻类囊体膜堆积会减少,叶绿素和类胡萝卜素的含量减少35%~50%[25];在空间种植的豌豆和拟南芥中也观察到叶绿体中类囊体膜的结构变化[26,27];在空间生长的小麦也被观察到具有较低的光合速率[28];甘蓝型油菜植物在空间中的光化学活性降低30%等[29]。在本研究中同样观察到光合作用中用于碳固定通路被下调的情况,这可能说明微重力条件下拟南芥生长水平的下降可能是光合作用水平下降造成的。

表5 拟南芥下胚轴下调反应对应基因的富集途径Table 5 Enrichment pathways for genes corresponding to Arabidopsis thaliana hypocotyl down-regulation reactions
3 讨论
利用特异性代谢网络和流平衡分析研究了微重力环境对拟南芥长势的影响以及其可能的生理机制。流平衡分析结果表明微重力环境会影响拟南芥生物质的合成和ATP 的产生,导致拟南芥幼苗、黄化幼苗、根、芽和下胚轴的生长水平比地面偏低,这与实验结果一致[22]。但是,拟南芥黄化幼苗相比正常幼苗,在微重力环境下生物质产量下降的程度较低,因此黄化幼苗可能更能适应微重力环境。
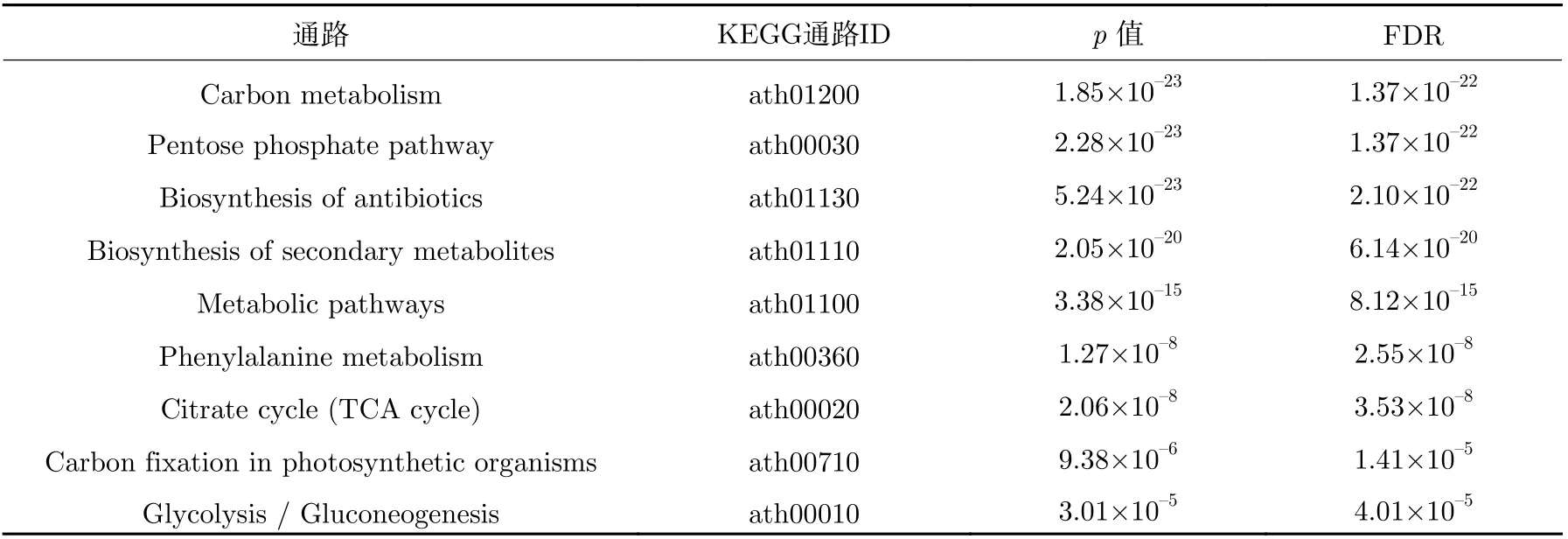
表7 拟南芥幼苗上调反应对应基因的富集途径Table 7 Enrichment pathways for genes corresponding to Arabidopsis thaliana seedlings up-regulation reactions
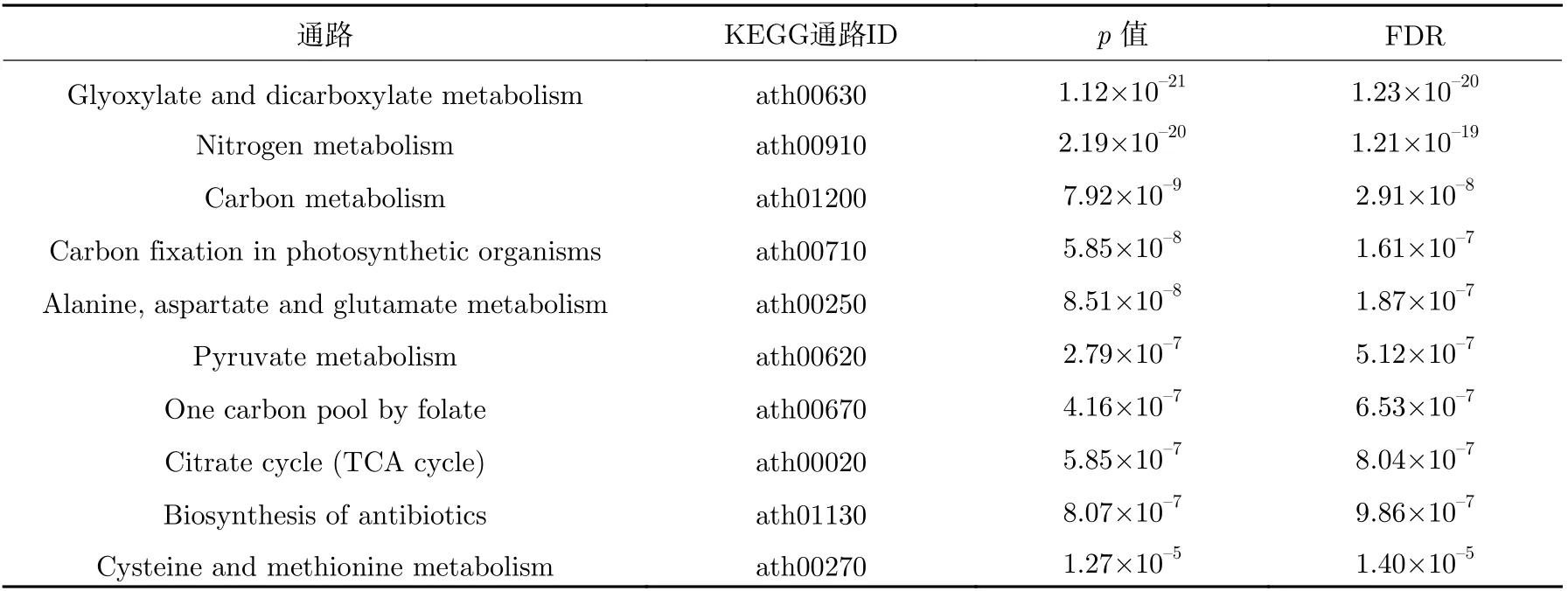
表8 拟南芥下胚轴上调反应对应基因的富集途径Table 8 Enrichment pathways for genes corresponding to Arabidopsis thaliana hypocotyl up-regulation reactions

表9 拟南芥根上调反应对应基因的富集途径Table 9 Enrichment pathways for genes corresponding to Arabidopsis thaliana root up-regulation reactions
拟南芥在微重力环境影响下的上下调代谢反应对应基因的KEGG 富集分析结果表明,微重力环境会影响拟南芥的多个代谢途径,并且导致拟南芥光合作用中碳固定通路的下调,这说明拟南芥生长水平的下降可能是由于光合作用水平下降造成的,这与已有的研究结果一致[25–29]。同时,拟南芥不同组织共同改变的代谢途径也可能为进一步的研究提供方向。
此前微重力环境对植物影响的研究多基于蛋白组或基因组,随着测序技术的发展,转录组数据相对容易获得,并且整合转录组数据的代谢网络可以系统研究植物在微重力条件下的代谢变化。研究表明,代谢网络模拟结果与实验观察结果有很高的一致性,并能够对实验结果进行机制解释。
4 结论
近年来,随着人类对空间探索的不断深入,空间长期生存所需的生物再生式生命保障系统成为空间生物学的研究热点。植物是该保障系统的关键要素,研究空间环境对植物生长发育的影响具有十分重要的科学意义。本研究通过流平衡分析初步探讨了微重力对拟南芥生长发育的影响机制,得到与实验观察一致的结果,即微重力环境会影响拟南芥的生长,使其生物质合成降低,导致拟南芥生长缓慢。同时,从拟南芥各器官组织来看,微重力环境对拟南芥根的影响最大,使其生物质和ATP 的产量较地面环境组均有所下降,这也与微重力环境下拟南芥出现较短的根的实验结果一致。流平衡分析表明,微重力环境下根较短的原因可能是线粒体电子传递链受到的影响较大导致其生物质合成降低。而导致拟南芥幼苗生物质合成下降的原因不是由于微重力对线粒体电子传递链的影响,关于其深层机制需要开展进一步分析研究。
