冰封期湖泊溶解氧变化特征及代谢速率*
2022-11-09杨文焕冯栋栋李卫平王乐乐
杨文焕,冯栋栋,杨 芳,李卫平,周 鑫,姚 植,王乐乐
(1:内蒙古科技大学能源与环境学院,包头 014010) (2:中国环境科学研究院,北京 100012) (3:内蒙古自治区凉城县岱海综合治理管理委员会,乌兰察布 013750)
湖泊水生系统中溶解氧(DO)浓度对湖泊生物多样性、营养盐生物地球化学循环、温室气体排放和饮用水质量等有重要影响[1-3]. DO一般随着水温升高而降低,且湖泊DO连续变化轨迹难以预测[4], 其变化过程受气温、水温、水生生物光合作用和呼吸作用、沉积物氧化还原反应等多种因素共同影响[5-7]. 世界上有1.17亿个湖泊,其中有近一半在冬季会进入到冰封期[8]. 与大量关于湖泊夏季研究文献相比,只有2%的文献报道了湖泊冰下溶解氧、热分布、有效光照及盐分等地球物理化学变化过程[9]. 其中,DO是影响冰下生态环境特征的重要因素,冰封期受冰盖阻碍,浅水湖泊与大气间的气体交换影响湖泊DO通量,进而改变湖泊新陈代谢及冬季水环境[10-11]. 在覆雪条件下微生物活动较剧烈的浅水湖泊,甚至出现缺氧现象[12-14],造成沉积物营养盐释放[15-16]并在冬季和早春引起氮、磷等营养物质的形态和浓度变化[17-18]. 冰封期湖泊冰下DO水平与新陈代谢具有一定响应关系,且受冰下水环境的净初级生产力(NEP)和生态系统呼吸速率(R)之间动态平衡的直接影响[19-20],Meding收集了23个北美温带湖泊的数据,指出在湖泊进入冰封期后平均40天内,溶解氧浓度保持不变或略有下降,然后迅速下降,当溶解氧浓度下降到1~3 mg/L时,溶解氧消耗速率逐渐减缓[21]. 与非冰封期相比,冰封期湖泊冰下水体的热分层颠倒、水温较低、水体承受的压强增大,以及风的混合作用和空气-水体交换基本停止,因此,冰封期溶解氧浓度的补充和消耗与非冰封期不同,且对冰封期湖泊生态系统的动植物种群和水质变化都有直接或间接的影响[9,11].
蒙新高原湖区存在着冰封期与非冰封期的交替变化,随着全球气温稳步上升,冬季覆冰湖泊生物化学循环过程越来越受到关注[22]. 在湖泊新陈代谢速率的计算中,溶解氧的平流传输、光呼吸和溶解有机碳的光氧化等非新陈代谢过程可能在短时间内引起DO的波动[23]. Obertegger等[24]、Song[20]在对浅水湖泊代谢速率计算中每30 min取一次溶解氧数据来计算时新陈代谢速率,再以此估算日新陈代谢速率,但较短的时间间隔存在非新陈代谢因素引起DO变化的可能,可能对日新陈代谢速率计算结果有一定影响. 传统的新陈代谢速率测定方法如黑白瓶法、叶绿素a法等,具有操作简单的优势,但易受天气影响[25-26],且难以获得观测期内逐日连续变化特征及逐小时生产力变化规律[27]. 本研究在典型寒旱区浅水湖泊岱海布设高频溶解氧仪和环境光度仪,通过不同深度下溶解氧高频数据分析岱海冰封期溶解氧的变化规律,并对冰封期湖泊新陈代谢速率进行估算,研究岱海冰封期溶解氧变化特征及其与环境因子之间的关系及冰封期浮游植物和微生物与环境的能量交换过程,揭示岱海冰封期溶解氧动态变化特征,对冰封期湖泊新陈代谢速率的计算方法进行进一步改进,与传统测定方法相比具有全天候、高频次的优势,可为冰封期湖泊缺氧治理提供新理论和技术参考.
1 材料和方法
1.1 研究区域概况
岱海(40°29′27″~40°37′6″N,112°33′31″~112°46′40″E)隶属于蒙新高原湖区,位于内蒙古自治区乌兰察布市凉城县境内,是内蒙古自治区三大内陆湖之一,也是我国“两屏三带”生态安全战略格局中“北方防沙带”的重要组成部分. 岱海东西长约25 km,南北宽约10 km,平均水深4 m,湖面面积约48 km2(2020年). 岱海流域四季分明,湖水主要补给水源为降水,多年平均年降水量为403.5 mm,平均年蒸发量为813 mm,年内降水量少而不均,蒸发强烈,夏季温和短促,冬季寒冷漫长. 岱海11月至翌年4月为冰封期,观测期间气温在-20~14℃之间,平均气温为-12.2℃,平均太阳总辐射为7.32 kcal/cm2,近年来,岱海水域面积不断萎缩,生物多样性下降,矿化度高,水质长期处于劣Ⅴ类[28].
1.2 检测指标
以6 km2为控制单元将岱海湖区划分为9个网格分区,结合岱海入湖河流的位置,在每个分区中心设置采样点(图1). 岱海浮游植物生物量较低,结合岱海水质逐月监测数据,在9个采样点中叶绿素a浓度最高的DH-4点(水深3.9 m)表层、中层、底层(表层0.8 m、中层1.8 m、底层3.7 m)处布置溶解氧仪(美国PME mini DOT),并在表层另布置环境光度仪(美国PME mini PAR),两种仪器每15 min记录一次数据(表1). 为提取冰封期更丰富的数据,2021年1月3日同步采集水温、pH、溶解氧等参数,透明度用赛氏盘法现场直接测定,并采用柱状分层采水器采集表层(水面以下0~1 m处)、中层(水面以下1~3 m处)、底层(湖底上方0~1 m处)水样各5 L,带回实验室完成总氮、总磷等水质指标检测(表2).

图1 岱海研究区位图Fig.1 Map of Lake Daihai research area

表1 观测仪器及精度
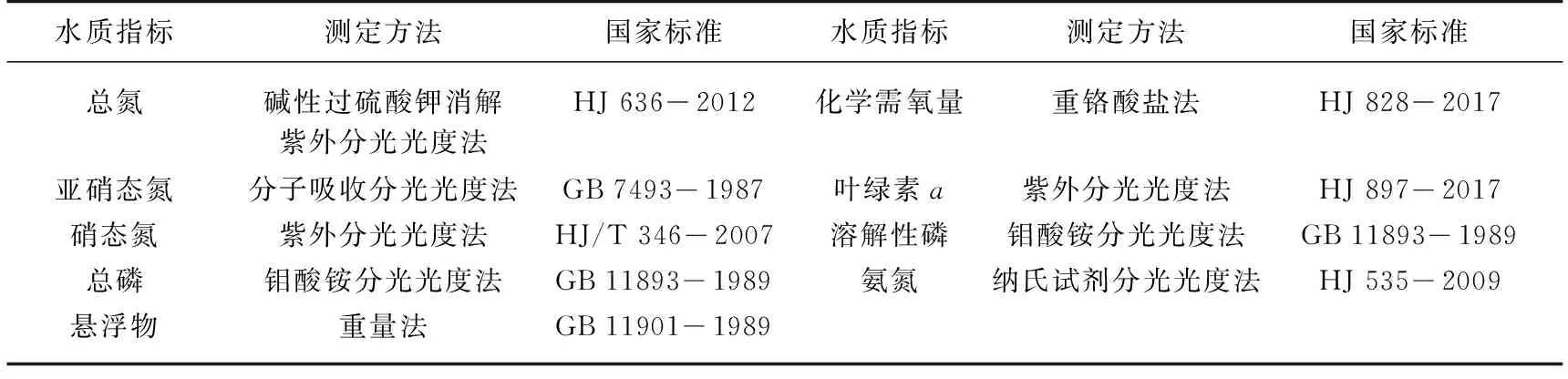
表2 实验室水质指标测定方法及国家标准
1.3 岱海冰封期新陈代谢速率的计算方法
生态系统的新陈代谢是指光合作用固定生物碳和呼吸作用氧化有机碳相关的基本过程,因此可以用产氧速率来衡量. 湖泊新陈代谢速率可以由总初级生产力(GPP)、净初级生产力(NEP)和呼吸速率(R)来表示[29].
Odum[30]首先建立了以DO来估算水中新陈代谢速率的计算方法:
ΔO2/Δt=GPP-R-F-A
(1)
式中,ΔO2/Δt为单位时间内DO变化速率;F为湖泊与大气的氧气交换;A代表除光合作用、呼吸作用和湖泊与大气的氧气交换之外其他引起DO变化的因素,在计算时每15 min记录一次DO值,然后计算整个水柱不同深度的DO平均值,因此其影响可忽略. 假设单位时间里白天呼吸作用所消耗的氧气(R昼)和夜晚呼吸作用所消耗的氧气(R夜)相同;以氧气溶于水的方向为正,则呼吸作用为负,光合作用为正. 夜间DO变化(ΔO2夜)通过查询当地逐日的日出日落时刻,再以当日日出时刻DO减去前一日日落时刻DO计算得出,由夜晚的呼吸作用与湖泊与大气的氧气交换共同作用:
ΔO2夜=R夜+F
(2)
全天呼吸作用所消耗的氧气(R)可以由夜间呼吸作用所消耗的氧气(R夜)和夜晚的时长(T夜)确定,则有:
R=R昼+R夜=24(R夜/T夜)
(3)
白天溶解氧的变化(ΔO2昼)由光合作用、呼吸作用和湖泊与大气的氧气交换共同作用影响,其值为当日日照最大值时DO减去当日日照0值时的DO,日净初级生产力(NEP日)与时净初级生产力(NEP时)可以由方程(4)、(5)得出:
NEP日=ΔO2昼+F
(4)
NEP时=ΔO2时+F
(5)
则GPP可以由公式(6)估算出:
GPP=NEP+R
(6)
F由公式(7)计算得出:
F=K(O2mean-O2sat)
(7)
式中,K是扩散系数;O2means是实际溶解氧浓度, 由溶解氧饱和度(O2%)乘以饱和溶解氧浓度(O2sat)得出.
K计算公式为:
K=K600(SC/600)-0.5
(8)
式中,K600是施密特数在600时的系数值,由公式(9)得出:
(9)
式中,U10是湖面上方10 m处的风速;SC为湖泊与大气的氧气交换系数,由公式(10)计算得出:
SC=0.0476T3+3.7818T2-120.1T+1800.6
(10)
式中,T为水温,当湖泊进入冰封期,冰盖的存在隔绝了空气和水体氧气的交换,可以将SC计为0.
2 结果与分析
2.1 岱海冰封期溶解氧分布特征
2.1.1 岱海冰封期不同水深溶解氧变化特征 2021年1月3日岱海湖区冰下表层水体DO浓度在13.06~14.78 mg/L之间,平均值为14.23 mg/L,是2020年夏季(2020年8月15日)的3.48倍,DO浓度在水平方向呈现出东北高西南低的特点(图2). 岱海整个冰封期内DO浓度在沿水深方向上出现分层现象,呈现表层>中层>底层的规律(图3),表层、中层和底层平均DO浓度分别为15.77、15.54和15.17 mg/L.
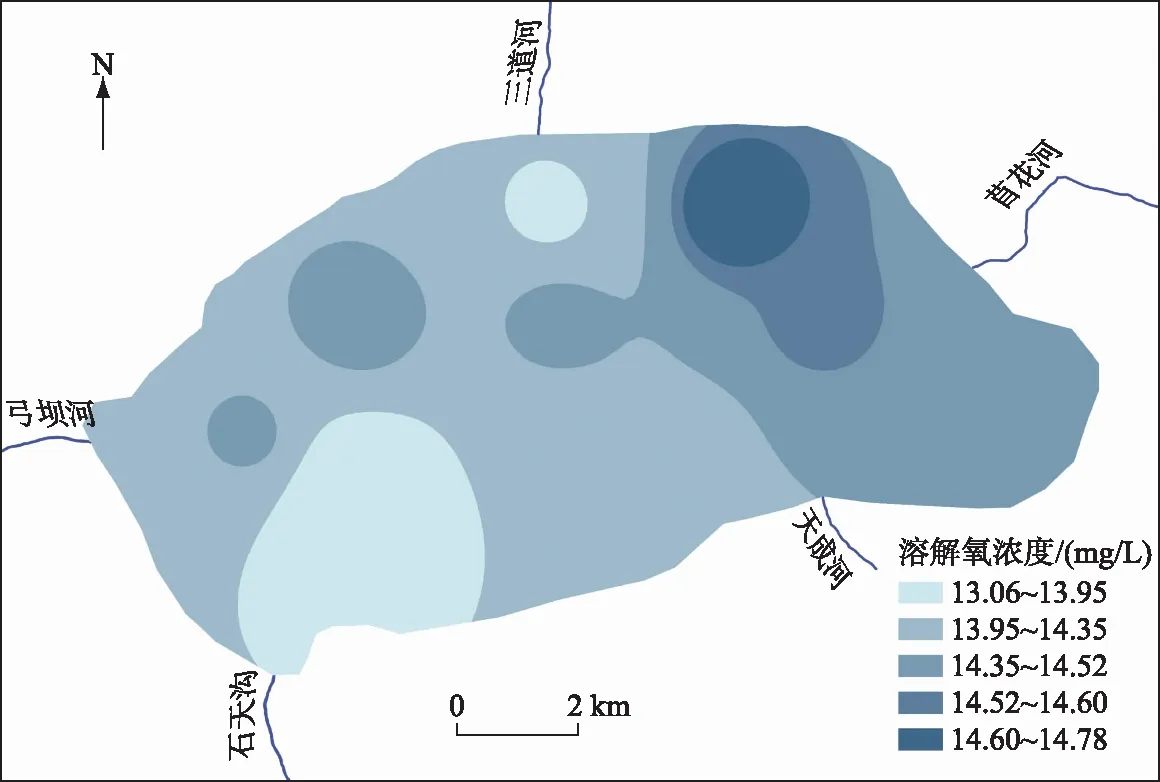
图2 2021年1月3日岱海DO浓度空间分布Fig.2 Distribution of dissolved oxygen concentration in Lake Daihai on January 3, 2021
岱海冰封期水体DO日变化表现为: 表层、中层、底层均表现出昼间上升,夜间下降的规律(图3). 其中昼间表层DO峰值比中层高0.25%,在夜间表层与中层基本相同,底层DO与表层和中层相比有较大的差距,与中层DO峰值相比下降了1.26%. 岱海冰封期表层DO昼夜变化幅度明显大于中层,这是因为浮游植物在冰封季节依然保持活性[17],且PAR会随水的深度增加而减小,因此表层浮游植物的生物量要比中层和底层更加丰富,初级生产力更强,表现出白天表层DO大于中层的现象[31];与表层和中层相比,湖泊底层DO变化曲线存在明显波动(图3),DO与水温波动存在一致性,这种现象可能与湖底表层的沉积物的氧化[32]及热通量释放有关[33].

图3 岱海冰封期溶解氧表层(a)、中层(b)和底层(c)浓度变化Fig.3 Changes of dissolved oxygen concentrations in the surface (a), middle (b) and bottom (c) of Lake Daihai during ice-covered period

图4 岱海冰封期溶解氧(a)和水温(b)剖面结构Fig.4 Profile structure of dissolved oxygen (a) and water temperature (b)during ice-covered period in Lake Daihai
2.1.2 岱海冰封期溶解氧浓度随时间的变化特征 2021年1月10日至3月11日,根据DO浓度和水温的变化规律(图4,图5)可知: 岱海表层、中层和底层水温在2021年1月10至2021年2月5日基本保持恒定,水温为-0.16~-0.13℃,表层、中层和底层日均DO浓度从14.41、14.49和14.29 mg/L上升至15.78、15.75和15.41 mg/L,期间DO浓度昼夜变化特征明显,昼夜DO浓度均有上升趋势,产生这种现象的原因是产氧代谢使DO不断积累. 2021年2月6日至2021年3月1日各层水温开始缓慢上升并出现昼夜水温差异,各层DO浓度基本维持不变,其原因是光合作用继续产氧但水温上升,水体的氧气溶解能力下降[6],DO在一定浓度范围内处于动态平衡;当岱海进入融化阶段(2021年3月2日至2021年3月11日)时,不同深度下水温差值明显增大,DO浓度昼夜差异逐渐消失,各层DO浓度变化混乱无序,此阶段DO浓度变化主要受水温影响.

图5 岱海冰封期气温(a)、PAR(b)和水温(c)变化Fig.5 Changes of air temperature (a), PAR (b) and water temperature (c) during ice-covered period in Lake Daihai
2.2 岱海冰封期新陈代谢速率的计算及变化特征
2021年1月10日至3月11日岱海冰封期产氧速率总体为正值,总初级生产力平均值为0.20 mg/(L·d),最高为0.57 mg/(L·d),最低为-0.30 mg/(L·d). 净初级生产力平均值为0.11 mg/(L·d),最高为0.46 mg/(L·d),最低为-0.09 mg/(L·d),总初级生产力和净初级生产力变化趋势一致.
冰封期湖泊在没有积雪覆盖时产氧代谢比较旺盛,PAR通过影响水-沉积物界面的温度来影响湖泊动植物呼吸速率[9,34],PAR对冰封期湖泊代谢速率具有决定性影响. 岱海产氧速率受天气变化影响明显(图6),2021年1月13日岱海出现降雪天气,降雪过后岱海冰下光合有效辐射显著降低,光合有效辐射峰值从12.62 μmol/(m2·s)下降到6.07 μmol/(m2·s),总初级生产力和净初级生产力下降明显(图6、图7). 相比于2021年1月13日,2021年1月14日PAR峰值有所回升,NEP回升显著,原因可能是2021年1月14日监测区域内仍存在积雪导致PAR并未显著回升,而湖面其他区域已无积雪覆盖,湖泊NEP总体上已经恢复.

图6 湖面有积雪(a)和无积雪(b)岱海冰封期净初级生产力的比较Fig.6 Comparison of net primary productivity between snow (a) and no snow (b)in Lake Daihai area during ice-covered period
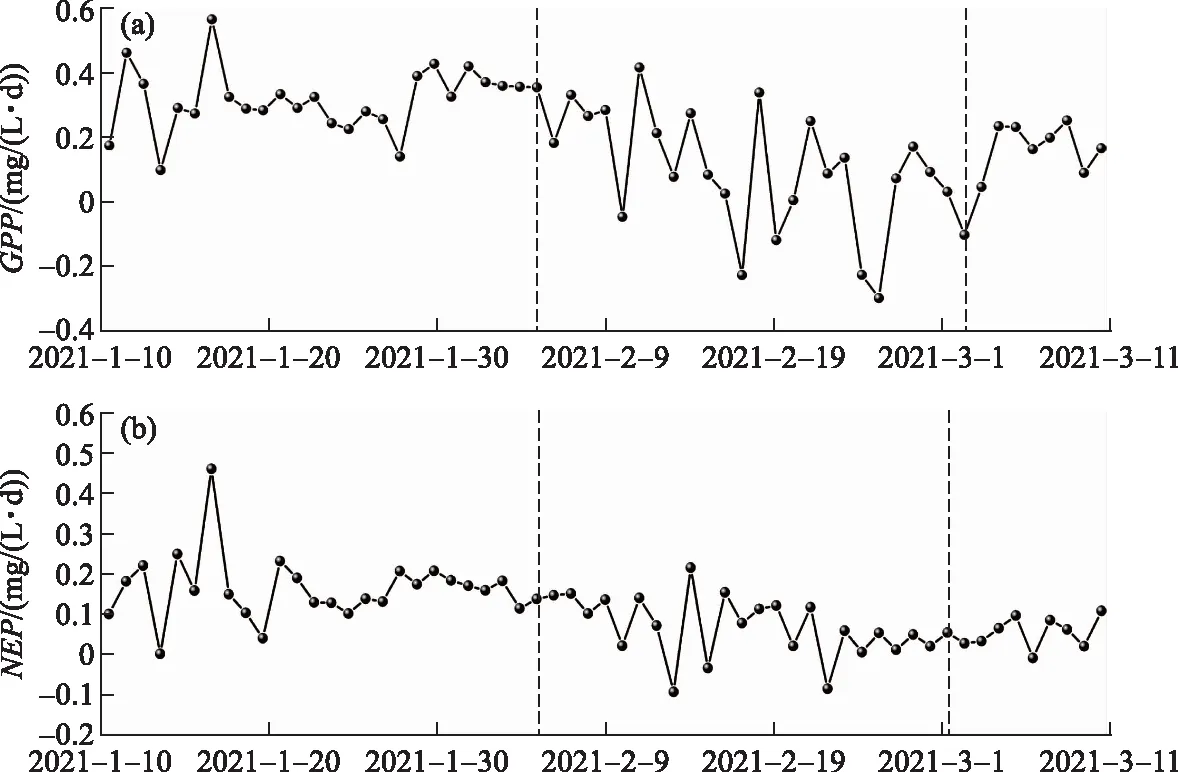
图7 岱海冰封期总初级生产力(a)与净初级生产力(b)Fig.7 Total primary productivity (a) and net primary productivity (b) during the ice-covered period of Lake Daihai
2021年1月10日至2月13日岱海的新陈代谢速率较为稳定,净初级生产力平均值为0.15 mg/(L·d),总初级生产力和净初级生产力都处于较高水平. 在2021年2月14日经历降雪后,总初级生产力与净初级生产力均有明显下降;之后水温升高,呼吸作用增强,2021年3月3日开始,净初级生产力恢复缓慢,总初级生产力恢复相对迅速.
除昼夜水温差异外(图5),湖体平流混合及其他水力作用也会影响监测点的DO,且湖体白天呼吸作用会大于夜间的呼吸作用,部分藻类在光线消失后仍可以进行产氧代谢[35],造成GPP和R的估算值偏小,因此,排除天气和仪器自身对DO扰动的可能,DO变化曲线也并不平滑(图3),可能出现部分负值(GPP不能为负值)及GPP变化曲线中个别GPP值过高的现象[36],但不会对NEP的估算产生影响[37].
一般认为水体中氮磷比(TN/TP)高于7则磷为藻类生长的限制因子,若低于7则氮为限制因子[38]. 2021年1月3日,岱海水体平均氮磷比为36.57,且整个冰封期内氮磷比均远大于7,同时DO与总氮呈显著负相关,与溶解性磷呈显著正相关,充分说明磷是影响岱海冰封期初级生产力的主要限制因子.
2.3 湖泊溶解氧与主要环境因子影响分析
2020年4月至2021年1月,对岱海逐月水质进行取样检测,其中冰封期总磷浓度与非冰封期相比未见明显变化,而溶解性磷则显著升高,比非冰封期高86%;冰封期pH在8.98~9.61之间,平均值为9.27,比非冰封期高4%;岱海冰封期水体总氮浓度平均值达4.06 mg/L,比非冰封期高8%,四季均处于劣Ⅴ类水平;冰封期电导率和盐度在16.51~24.58 mS/cm和9.68×10-6~14.89×10-6mg/L之间,与非冰封期相比未见明显变化. 与非冰封期相比,冰封期的溶解性磷、总氮、pH等指标值均有一定升高,水质呈现恶化趋势.
开展非冰封期DO与主要环境因子的冗余分析(RDA),以DO作为主轴的平面内,轴1和轴2的特征值分别为0.1078和0.8301(表3,图8),在岱海非冰封期的8个月中,10、11月的DO与溶解性磷、COD呈正相关,其余呈负相关;4、5、6月DO与硝态氮、总氮、氨氮、总磷、叶绿素a、电导率、pH均呈正相关,与COD、溶解性磷呈负相关;7、8、9月的DO与溶解性磷、电导率呈现明显的负相关性.

表3 岱海非冰封期溶解氧和环境因子RDA分析结果
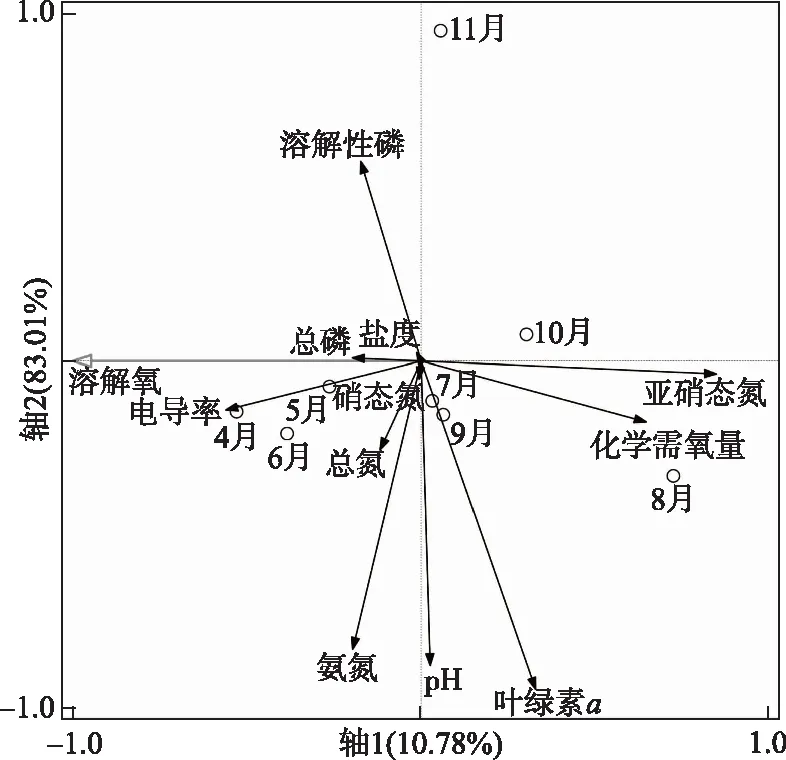
图8 岱海非冰封期DO和环境因子RDA排序图Fig.8 RDA sequence diagram of dissolved oxygen and environmental factors innon ice-covered period of Lake Daihai
对岱海冰封期DO与物理因子(pH、氧化还原电位、电导率、盐度、水温、光合有效辐射)和主要营养盐(总氮、氨氮、硝态氮、亚硝态氮、总磷、溶解性磷)等环境因子进行相关性分析. 相关系数矩阵结果显示(表4),DO与总氮、pH、水温呈负相关,与溶解性磷和光合有效辐射呈正相关,其中光合有效辐射、水温、总氮、pH与DO相关性极显著,溶解性磷通过影响水体初级生产力[4]来影响岱海冰封期DO.
在非冰封期,湖泊表层水体DO主要来源于大气与水界面的气体交换,中下层水体DO主要通过水体垂直对流扩散作用获得,与水温、水体热力分层、pH、悬浮物、高锰酸盐指数等有重要关系[39],而冰封期湖泊DO主要是由结冰前水体DO、冰封持续时间和底层有机物分解消耗等因素所决定[40]. 非冰封期不同时段岱海DO与物理因子、环境因子的相关性出现明显的差异,这可能是水温与生物生长周期共同作用的结果[17];而冰封期DO则主要是受水温与光合有效辐射等因素的影响[13],水体中氮磷等营养盐通过影响生产力来影响水体的产氧代谢速率[4,38].
3 讨论
3.1 湖泊冰封期溶解氧过饱和的原因
一般认为湖泊底层缺氧的原因是上层水域通过光合作用产生的有机物在底层水域分解,导致底层DO的消耗. 在水温分层的季节,温度梯度限制了表层水域向底层水域的氧气供给,造成底层氧气浓度持续下降,甚至出现缺氧现象[41-43]. 在岱海冰封期连续监测中发现,部分时段出现DO分层现象,平均DO达15.49 mg/L,表层DO处于过饱和状态,原因如下:
在淡水中不同温度所对应的DO理论值计算公式为[44]:
CDO=477.8/(T+32.26)
(11)
式中,CDO表示溶解氧浓度;T表示水温.
2021年1月10日至2月5日实际监测DO与计算的理论值极为接近,实际值比理论值高0.02~0.34 mg/L左右,说明该理论值计算公式可用于描述岱海冰封期DO的动态变化,结合水温与DO间的相关性分析,结果发现水温与DO相关性极显著,相关系数为0.833(P<0.001),即岱海进入冰封期时水温下降,氧气溶解能力增强,表现为冰封期DO浓度升高甚至过饱和状态.
冰封期内浮游植物光合作用生产的氧气在冰水界面间不断积累使表层DO不断升高. 岱海在冰封期净初级生产力总体上为正值,2021年2月6日至3月1日实际DO浓度比理论DO浓度高1.09 mg/L,与2021年1月10日至3月11日实际DO浓度和理论DO浓度的差值相比有明显升高,且期间DO高频观测结果也表现出明显的光合产氧特征,充分说明岱海冰封期仍然具有光合作用,并且光合产氧量大于呼吸作用耗氧量,因此在冰层未化的冬末初春时期,出现冰下水体DO积累现象.
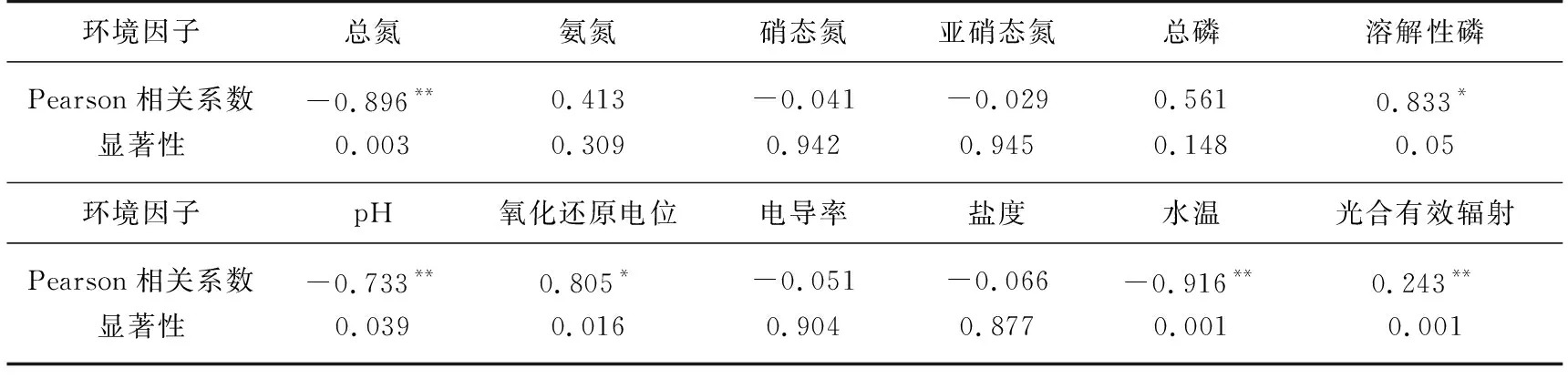
表4 岱海冰封期溶解氧与环境因子的Pearson相关系数
3.2 同纬度下湖泊冰封期溶解氧变化趋势的差异
岱海在冰封期内,湖体表层、中层和底层DO始终保持着较高水平,整体上呈现出先逐渐升高,达到一定浓度后保持不变,进入消融阶段DO分层现象消失,表层、中层和底层DO趋于一致的变化规律,并未发生底部缺氧现象,这与意大利贫营养湖泊Tovel湖观测结果相同[24]. 而Bai等[13]、Song[20]]、Zhai等[40]在对与岱海纬度相近且同属于蒙新高原湖区的乌梁素海进行冰封期DO研究时,发现乌梁素海底层(2.06 m)DO在整个冰封期内基本处于缺氧状态,表层(0.7 m)和中层(1.06 m)的DO呈现出逐渐下降的趋势,降雪后冰下DO急剧下降,冰盖消融后DO又逐渐上升,与岱海情况略有不同.
同纬度湖泊冰封期DO变化趋势出现差异的原因有:大部分浅水湖泊(水深1~3 m)水生植物可以覆盖整个湖底,具有高密度和高生物量的特点,这些水生植物和浮游植物在冰封期衰亡,将释放大量溶解态和颗粒态的有机物,其中冰封期湖泊DO消耗速率主要取决于湖泊中颗粒态有机物(POM)的氧化分解[45]. 岱海浮游植物生物量远低于乌梁素海,尤其沉水植物生物量极低,而乌梁素海沉水植物生长区域占其湖区面积的96.2%[46-47],每年沉积湖底的大型水生植物残骸约为20.5×104t,是典型重度富营养化的草型湖泊[48],冰封期缺氧的风险较大. 岱海净初级生产力高于乌梁素海且生物量远低于乌梁素海,因此岱海冰封期湖底未发生缺氧现象. 其次,乌梁素海湖泊面积大,部分湖区积雪覆盖长达20天以上,导致冰下光合有效辐射在积雪覆盖期显著下降,净初级生产力大部分时间为负值,产氧速率受到严重影响;但是岱海冬季漫长而寒冷,多风且干燥,难以积雪,经历降雪后,冰下光合有效辐射1~2天即可恢复至降雪前水平,积雪对岱海产氧代谢影响有限.
湖泊水深、总磷和叶绿素a浓度、水-沉积物界面的热通量、硝化作用等与冰封期湖泊耗氧过程同样有密切联系. Barica和Mathias研究发现,冰封湖泊单位体积的耗氧量与平均深度呈显著负相关(R2=0.92)[49],这可能是由于水深不断增加,光合辐射被逐渐削弱,不利于水生植物的生长和POM的积累,岱海平均深度约4 m,乌梁素海平均水深为1.5 m,因此乌梁素海生物量和冰封期耗氧量均高于岱海;叶绿素a和总磷浓度主要通过影响冰封期湖泊新陈代谢速率来影响DO[50-51],岱海全年叶绿素a浓度极低,冰封期仅有1.2 mg/L,且冰封期中较低的磷浓度限制了岱海新陈代谢速率的发展,这些造成了岱海冰封期产氧速率处于较低水平;秋冬季湖泊水柱迅速变冷,沉积物会保存一定热量[52]维持冬季底层水温,岱海面积只有乌梁素海的1/6,存储热量远小于乌梁素海(冰封期底层水温可达7℃),而岱海冰封期底层水温在1℃左右,表明岱海水-沉积物界面的热量交换作用不强,不是影响DO变化的主要因素;在冰封期缺氧的湖泊中,硝化细菌耗氧量占总耗氧量的1%~25%[53],而岱海冰封期未检测到氨氮浓度的下降和硝态氮的积累,表明岱海冰封期硝化作用并不明显. 因此,湖泊深度、POM浓度、当地气候及总磷和叶绿素a浓度的不同是导致岱海与乌梁素海冰封期DO出现差异的主要原因.
4 结论
岱海冰封阶段(2021年1月10日至2月9日)DO从14.39 mg/L上升至15.65 mg/L,之后DO在15.6 mg/L保持动态平衡,进入融化阶段(2021年3月2日后)DO分层现象逐渐消失;水温垂向上呈现均匀混合状态(2021年1月10日至2月5日)向分层状态(2021年2月6日至3月11日)转变的特征.
岱海冰封期净初级生产力平均值为0.11 mg/(L·d),水温和积雪对新陈代谢速率有明显的影响:水温升高导致呼吸作用增强,从而影响氧气的积累;积雪通过影响冰下光合有效辐射来影响新陈代谢速率.
岱海冰封期DO与溶解性磷和光合有效辐射呈正相关,与总氮、pH、水温呈负相关,物理因子中以水温和光合有效辐射对DO影响最大,营养盐中总氮与DO变化关系密切.
