模拟降水变化下黄土丘陵区草地群落特征及其物候期响应
2022-11-09盛翰苑翟林博曹家仪
盛翰苑, 翟林博, 曹家仪, 焦 峰,2
(1.西北农林科技大学 资源环境学院, 陕西 杨凌 712100; 2.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100)
黄土丘陵区的生态环境比较脆弱,长期以来由于自然和人为因素,导致该区的植被遭到严重破坏,水土流失严重,对人民的生活与生产建设造成了严重的影响[1]。因此,该区的植被恢复与重建问题变得尤为迫切。土壤水分作为制约干旱、半干旱黄土丘陵区植物生长的重要因素[2],是黄土丘陵区植被恢复与重建的关键所在。大气降水是黄土丘陵区土壤水分的主要来源,亦是驱动该地区植物物候期和群落特征改变的重要因子之一。在全球气候变暖背景下,我国黄土高原降水格局呈现出季节波动增强和极端降水事件增加趋势,而降水格局变化势必会引起草地群落特征和物候期发生重要改变。特别对于生态脆弱的黄土高原地区来说,植被对降水的变化响应较为敏感,降水格局改变会显著影响植物物候、群落分布格局以及生理生态特征,进而影响到生态系统的功能和稳定性[3-4]。
植物物候是植物受其内因和所处环境(如气候、水文、地形等)影响产生的以年为周期的自然现象[5],是植物长期适应环境的季节性变化而形成的生长发育节律[6]。有研究表明,干旱处理能使某些植物花期提前[7]。也有研究表明年降水减少可延迟灌木植物的开花和衰老的开始时间[8]。不同物种开花时间、峰值期和开花持续时间等过程也会对群落多样性、结构和功能产生影响[9]。有学者研究一年生植物Brassicalapa(芜菁)在持续干旱的环境中的物候特征,发现植物的生命周期缩短,干旱促使植物开花提前,更早地进入繁殖物候,从而在资源匮乏的环境中完成生活史[10]。我国对草本植物物候动态变化及其对气候变化的响应、不同物候期与环境因子(包括温度、光周期、土壤水分、CO2及冬天的寒冷程度等)的相互关系研究颇多。不论是长期观测研究还是试验研究,大多结果表明,草地群落物种物候期与降水量的存在不确定性[11-12]。草地是干旱半干旱地区黄土高原植被组成的主体,而草本植物作为草地生态系统的优势群落,在黄土高原恢复植被和水土保持建设方面存在着不可替代的优势,是研究物种多样性和土壤水分相互作用机制的理想对象和区域[13]。鉴于此,为揭示植物物候期和群落特征对降水变化的响应规律,需要开展定点观测降水控制试验方面的研究。本研究选择黄土丘陵区退耕草地为研究对象,设置野外定点观测试验站,利用遮雨棚装置模拟不同降水梯度处理,通过连续观测植物物候变化特征和群落结构变化,以揭示植物物候和群落特征对降水变化的响应规律,以期为应对未来全球气候变化下黄土丘陵区植被恢复和重建提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于中国科学院安塞水土保持综合试验站山地试验场(109°19′23″E, 36°51′30″N),系典型的黄土高原丘陵沟壑区。气候类型为暖温带半干旱气候。海拔1 068~1 309 m,年平均气温8.8℃,气温日较差较大;年平均降水量505.3 mm,平均相对变率为16.3%,最多为645 mm,降水分布不平衡,多集中在6—9月份。主要土壤类型为黄绵土,土质绵软,结构疏松,属极强度侵蚀区。
1.2 研究方法
1.2.1 样地布设 选择退耕草地为研究对象,2015年对样地进行前期整地和除杂处理,在样地四周布设围栏保护试验不受人类和动物的干扰。样地内主要的植物物种为达乌里胡枝子(Salespeoplecaricature)、狗尾草(Secretariats)、早熟禾(BoaanneaL.)、茵陈蒿(ArterialSpillaneThumb.)、铁杆蒿(Arterialsacrilegious)等。
本实验设置7个降雨梯度分别为DP60(-60%),DP40(-40%),DP20(-20%),CK(0%),IP20(+20%),IP40(+40%),IP60(+60%),每个降水梯度设置5个重复,共35个实验小区,每个试验小区面积为3 m×3 m,各试验小区间设2 m的间隔缓冲区域。通过搭建遮雨棚装置来模拟降水梯度变化,该装置由3部分组成:支撑钢结构、透明挡雨V型槽(透光率>92%的有机玻璃,厚2 mm,长1.8 m,V型口处宽0.1 m,与水平面夹角20°)、集水塑料桶(图1)。根据减少降雨的梯度对挡雨V型槽的数量进行增减,在雨后将集水塑料桶中集水均匀的喷洒在对应增雨的实验小区内。为了保证实验小区免受边际效应、坡位地形、气象因子等扰动,所有实验小区均位于距其它类型的土地利用类型50 m以上的同等地形下。降水实验自实验小区布设的第一年起连续进行。

图1 降雨控制试验装置示意图
1.2.2 物候观测 物候观测于2019年4月初—10月中旬进行,每次隔7 d观测1次,在植物生长关键期增加观测次数。调查方法是在试验小区中心划分出1 m×1 m调查样方,记录样方内的每种植物的物候期,每次调查时样方的位置保持不变。为了方便数据处理,本文利用儒略历计数法(即1月1日至该日期的日数)得到各物候期的时间序列进行统计分析。物候期划分方法:植物物候期包括返青期、蕾期(抽穗期)、始花期、花末期、初果期、果熟期、果后营养期、枯黄期。
1.2.3 群落特征调查 群落特征调查与物候期观测的样方大小及位置一致,每间隔15 d左右调查1次样方内草本植物的物种、株数、株高、覆盖度和上生物量。其中物种、株数、株高通过人工调查方式获取;覆盖度采取垂直拍摄样方照片,用Image-J软件进行灰度处理,计算面积获取;为避免对试验小区内植物生长的扰动,地上生物量采取估算法进行获得,具体步骤如下:根据样方调查的物种的株高分布,选取3个有代表性的株高在试验外的小区内找对应株高的植物,从地表剪下,按照物种和株高对分别装入不同的信封,带回室内在75℃下烘干48 h至恒重(0.01 g),然后计算出单物种的地上生物量,样方内各物种的生物量之和为地上生物量。
本文采用物种丰富度、物种均匀度指数和Shannon-Wiener多样性指数共同表征多样性,计算方法如下:
① 物种重要值:
Pi=(a+b+c)/3
(1)
② 物种丰富度:
M=(S-1)/lnN
(2)
③ Shannon-Wiener多样性指数:
(3)
④ Pielou均匀度指数:
J=H/lgS
(4)
式中:Pi为物种的重要值;a为相对频度;b为相对盖度;c为相对密度;M为Margalef指数;S为样方内总物种数;N为所有物种的个体总数。
1.2.4 数据处理 运用WPS 2019对数据进行简单预处理,用SPSS 23.0进行单因素方差分析(one-way ANOVA)与Dunces test多重比较,分析不同降水梯度下物种物候期、繁殖期和生长季长度和群落结构特征(物种组成、重要值、群落盖度、地上生物量、多样性)参数差异。在进行数据分析之前对各类数据进行正态分布和方差齐性检验。采用Origin 2021绘图。
2 结果与分析
2.1 降水改变对自然恢复草地群落物种组成的影响
半干旱黄土丘陵区草地群落生长季为4—10月,8月是植物生长达到高峰的时期,8月物种重要值变化可以较好地反映草地群落的物种组成与结构状况的变化情况。故以8月物种重要值为依据,对草地群落在不同降水梯度下物种组成差异进行分析。由表1可知,群落中共出现24个物种,一年生草本6种,多年生草本18种。24个物种分属9科20属,其中以豆科(7种)和菊科(7种)为主,以及禾本科(4种)、藜科、大戟科、紫葳科、牦牛儿苗科、堇菜科和唇形科。群落中达乌里胡枝子、茵陈蒿、狗尾草和早熟禾等为主要优势物种,黄鹌菜、草木樨状黄耆、披针叶黄华、猪毛菜和苣买菜等为主要伴生物种。不同降水处理下群落物种组成存在较明显差异。从表1可以看出,增雨处理和减雨处理均使群落物种种类增加,其中增雨60%和减雨60%时使群落物种种类增加最多,均为18种。增雨处理与减雨处理时增加群落物种种类无明显差异。与对照相比,减雨20%,40%,60%处理均使达乌里胡枝子的重要值明显增加,早熟禾、茵陈蒿、旱芦苇、蒙古蒿和猪毛菜等物种的重要值明显降低,且部分物种重要值呈随减雨程度增加而增加的趋势。其中牦牛儿苗和沙打旺等旱中生物种为减雨处理中所特有物种,表明减雨环境一定程度上会促进耐干旱植物生长。与对照相比,增雨20%,40%,60%处理均使茵陈蒿的重要值明显降低,达乌里胡枝子、蒙古蒿和猪毛菜等物种重要值亦有不同程度的降低,增雨20%和40%使狗尾草重要值明显增加。增雨20%~60%处理时,苦豆子、野豌豆、苣荬菜和铁杆蒿等物种重要值均随着增雨程度增加而降低,表明增雨量过多在一定程度上降低了物种的生长优势。

表1 不同降水处理下草地群落物种组成及其重要值
2.2 降水改变对草地群落结构的影响
图2为草地群落在7个不同降水梯度处理下群落盖度和地上生物量5—10月动态变化情况。总体上,群落地上生物量呈先增后减的趋势,8月达到最大值,9—10月又逐渐降低。增雨处理明显高于减雨处理,对照组地上生物量在5—8月均显著低于减雨20%、减雨40%、增雨20%、增雨40%和增雨60%,减雨20%在5—8月显著高于减雨40%,60%,5—10月增雨20%处理明显高于其他各组。出现这种结果的原因可能是,8月之后降雨减少,多数1年生草本植物完成生活史,生命周期仅有教短的40~60 d,导致群落地上生物量大幅减小。原因可能为,当降水不断增加,水分胁迫现象得以缓解,草本群落生物量随之增加;但当降水增加植物可以利用的水分上限时,反而会抑制植被呼吸等生理生化反应,导致植被生物量出现下降趋势。群落盖度呈先增加后降低趋势,这与植物生理生长规律相一致。增雨20%处理下,5—10月的群落地上生物量均显著高于其他各处理。相比于对照,增雨20%、增雨40%均使盖度显著增大,减雨60%、减雨40%、减雨20%、对照、增雨40%和增雨60%处理下盖度均显著低于增雨20%处理。7—10月群落盖度对降水变化敏感,不同降水处理之间盖度差异不断加大,而5月和6月组间差异较小。

注:不用字母表示不同降雨梯度之间的差异显著(p<0.05),下同。图2 不同降水处理下草地群落盖度和地上生物量差异
2.3 降雨改变对草地群落物种多样性的影响
由表2可知,群落物种Shannon-Wiener指数和Pielou均匀度指数在不同降雨处理下的存在显著差异。单因素方差分析结果不同降水处理下群落物种丰富度无显著差异。与对照相比,丰富度和Shannon-Wiener指数总体上都呈增加趋势,减雨20%,40%,60%均使Pielou均匀度指数降低。群落中增雨60%处理时物种丰富度最大(2.07±0.33);减雨40%处理时群落Shannon-Wiener指数最大(2.16±0.04),减雨20%最小(1.75±0.02);减雨40%处理时群落Pielou均匀度指数最大(1.21±0.04),增雨40%时最小(0.91±0.04)。
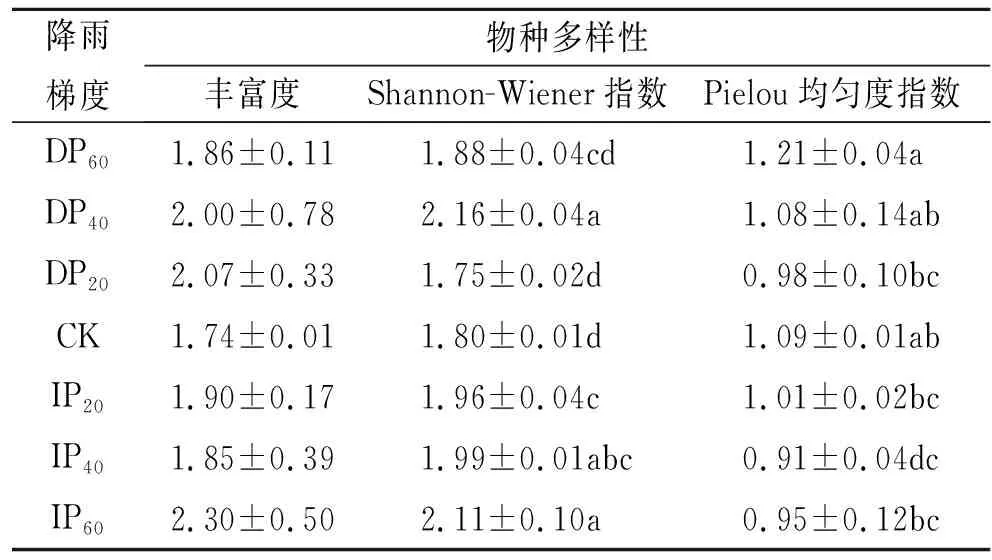
表2 不同降水处理下群落物种丰富度和多样性指数
2.4 草地群落物种物候期对降水改变的响应
综合不同降雨处理对群落各物种的生长优势的影响,分析群落中茵陈蒿、早熟禾、达乌里胡枝子和狗尾草的物候期差异,发现植物返青期、繁殖期和枯黄期具有明显的物种间差异,不同降水条件也会改变植物的物候期,且具有显著的影响(p<0.05)。由图3可知,与对照相比,增雨20%,40%,60%处理均使达乌里胡枝子和茵陈蒿的返青期、蕾期、果期和果后营养期有所提前。增雨处理使达乌里胡枝子返青期提前2~3 d,蕾期提前2~3 d,果期提前1~2 d,果后营养期提前2~3 d,枯黄期提前2~4 d。减雨处理使达乌里胡枝子返青期提前5~7 d,枯黄期显著提前3~5 d,说明减雨对促进其返青期提前更加显著。增雨和减雨处理使早熟禾返青期推迟2~3 d,对蕾期无影响,增雨各处理使黄枯期提前1~3 d。茵陈蒿返青期在增雨和减雨各处理下均发生提前,减雨提前4~5 d,增雨提前2~3 d。而狗尾草在各降雨处理下的返青期差异表现并不显著。

图3 不同降水处理下草地群落各物种物候期
由图4可知,减雨20%,40%,60%处理分别显著提前达乌里胡枝子繁殖期7,6,5 d,增雨60%使其繁殖期显著延迟3 d,增雨20%,40%与对照无显著差异。减雨对早熟禾和茵陈蒿的繁殖期无显著影响,增雨20%,40%显著提前早熟禾繁殖期5 d,增雨60%使茵陈蒿繁殖期显著延迟7 d。减雨各处理、增雨20%和增雨40%均使达乌里胡枝子的生长季长度显著增加;减雨各处理、增雨40%和增雨60%均使茵陈蒿的生长季长度显著增加;增雨40%和60%使狗尾草生长季长度降低,其他处理均与对照无异;减雨40%和增雨20%显著降低早熟禾生长季长度,减雨20%显著提高其生长季长度。
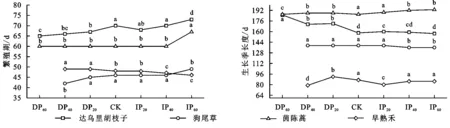
图4 不同降水处理下草地群落各物种繁殖期和生长季长度
3 讨 论
3.1 降水变化对草地群落物种组成的影响
本研究发现,群落物种组成在不同降水梯度中的响应规律存在明显差异。群落中达乌里胡枝子、茵陈蒿、狗尾草和早熟禾等为主要优势物种,减雨使达乌里胡枝子、牦牛儿苗和沙打旺等的重要值增加,增雨增加了狗尾草和苣荬菜的重要值,这表明降水变化不仅影响群落物种组成而且能够改变群落优势物种,群落中多年生草本物种在面对干旱环境时更能体现出优势,这与多年生物种较发达的根系以及保守型资源获取策略有关[14]。降水增加时,生长迅速的一年生物种根系较浅,生长快,高度依赖降水,因此增雨反而利于这些进攻型资源获取植物生长[15]。增雨条件下,植物生长不再受水分的限制,此时群落中所有植物均开始迅速吸收水分以促进其生长,导致物种间竞争加剧,物种生态位不断扩展重合,大部分优势物种和多年生物种重要值因此而降低。相比于降水增加,降水减少对荒漠草原植物群落生长特征的负面影响更为显著。在黄土丘陵区,草本植物的种类、多度和高度会受到降水格局的影响,从而影响植物群落的分布与结构。
3.2 降水变化对草地群落结构的影响
地上生物量是植物地上部分光合同化产物的质量,是植物当年净初级生产力的重要指标,对预示群落功能和稳定性具有重要作用[16]。本研究发现,群落增雨处理下的地上生物量均明显高于减雨处理,说明降水增多促进植物地上部分有机质积累,与过往研究结果一致[17-18]。降水增多对草地生产力具有显著的正向促进作用,降水增加提高了土壤水分含量,植物不再受到水分限制,竞争由地下转为地上竞争,促进植物地上部分生物量的积累[19]。群落地上生物量随减雨和增雨梯度增加而逐渐降低,而增雨20%和减雨20%能够明显提高群落生产力,这与群落盖度规律相一致。极端降水会导致土壤水分过饱和,根系呼吸作用和吸收作用均受到抑制,对植物生产力产生明显的负向效应。
群落盖度反映了植被覆盖地表的范围,是植物利于光能的重要指标,控制着群落生产力基本状况。5—8月,增雨20%处理下群落盖度均显著提高群落盖度,减雨20%处理仅次于增雨20%,而对照组群落盖度却始终较低或最低,这表明轻度干旱和适当增加降雨能够显著提高群落盖度[20]。降水增加超过20%以及干旱程度超过20%将会降低群落盖度,这与孙一梅等[21-22]的研究结果相一致,表明草地群落对降水的响应存在一定的阈值,重度干旱和过度降水都不利于植物生长。重度干旱对植物生理过程产生明显的抑制作用,减少叶片面积来减少蒸发,降低光合速率,在群落层面表现为群落盖度减少,草地生产力大大降低[23]。过度降水可能引起土壤水分过饱和,抑制植物根系的有氧呼吸,植物养分吸收和光合能力受限,涝害严重将导致植物萎蔫或死亡[24]。
3.3 降雨变化对草地群落物种多样性的影响
植物物种多样性研究是生物多样性研究中植物适应特定环境最直接和最适合的表达层次。本研究发现,丰富度和Shannon-Wiener指数总体上都呈增加趋势,减雨20%,40%,60%均使Pielou均匀度指数降低。结果出现的原因可能是,随着降水量的增加,植物可以利用的水分增加,物种间出现生态位互补效应,种间竞争呈现积极相互作用,使物种共存概率增大,群落物种多样性随之增加。而降水量的减少导致样地水分资源的紧缺,尤其是干旱程度严重时,物种生态位重叠程度加大,物种间的竞争加剧,弱势物种生长受到较大抑制,甚至被淘汰。
3.4 不同物种物候期对降水变化的响应
由于不同物种的植物功能性状及其对生长环境的响应和适应不同,不同植物物候期对环境变化的响应存在明显的差异[25]。本研究观测了4种主要物种(分别为达乌里胡枝子、狗尾草、早熟禾、茵陈蒿)物候期,发现达乌里胡枝子返青期对降水变化比其他4种植物更为敏感。降水变化对物候的影响的可能原因有两个:① 降水增加或减少使得土壤含水量也随着升高或减少,提前或推迟达到植物特定物候所需的水分条件,从而造成草地植物特定植物物候的提前或推迟。② 降水变化导致土壤水分发生改变,影响植物生长发育过程中与水分相关的一系列生理生化反应,从而间接影响植物物候[26]。因此对于温度变化而言,植物物候期对降水变化更为敏感,这与在青藏高原高寒草甸的研究结果一致[27]。
在物候的观测记录中仅发现降水变化对黄土丘陵区的自然恢复群落中的达乌里胡枝子物候产生显著影响。大量研究表明,影响植物物候期的主要环境因素是气温、降雨和日照时间[28]。对于光照充足的干旱-半干旱黄土丘陵区,气温和降水量的变化是影响植物物候期的重要因素[29]。在本研究中,在植物生长季的前期,降水量较少,植物物候前期对降水量的改变较为敏感,雨季较多的降水量而且降水强度较大,不利于植物生长旺盛期物候的发生,而且生长季末期降水量的增多对植物枯黄期的影响也不显著。但是在其他不同地区的研究结果表明,植物的返青和枯黄等物候期与前期的温度和降水显著相关[30]。本研究发现不同植物物种对不同降雨水平的响应是存在差异的,产生这种差异的原因是由于植物物种本身在繁殖期的适应机制和生存策略不同,这是一个复杂的过程[31]。
4 结 论
(1) 自然恢复群落达乌里胡枝子的返青期、繁殖期和枯黄期在减雨处理下显著提前,而增雨处理则推迟了其返青期和繁殖期。增雨和减雨处理显著推迟狗尾草和茵陈蒿的繁殖期,但对其他物种无显著影响。不同降水处理使自然恢复群落物种枯黄期发生延迟,从而延长了各物种生长季长度。
(2) 自然恢复群落物种组成、植被盖度和地上生物量在不同降水处理下表现出显著差异,而物种多样性对降水变化响应不敏感。增雨和减雨处理对物种组成影响主要体现在物种重要值上,减雨处理提高了群落中多年生优势物种重要值,而增雨处理提高了1,2年生物种重要值。草地群落植被盖度和地上生物量对降水变化的响应规律相似,5—8月生长季对降水变化较为敏感,而生长季末期对降水变化不敏感,增雨20%处理下群落盖度和地上生物量最大,过度增雨或减雨均会引起草地群落盖度和地上生物量降低。
