佛坪国家级自然保护区大熊猫与潜在捕食者的时空动态
2022-11-07马青青马亦生陈原玉刘红彩张振华淡中豪
马青青 马亦生 陈原玉 刘红彩 周 刚 张振华 杨 丽 淡中豪 刘 坤
(1.陕西省渭南市农业技术推广中心,渭南,714000;2.陕西佛坪国家级自然保护区管理局,佛坪,723400)
生态位分化理论常被认为是物种共存遵守的基本法则。种间竞争的物种一般通过调整栖息地利用、食性和活动节律等生态位分化实现共存[1-2],而食肉动物则通过与猎物在时空生态位的重叠以提高捕获率[3-4]。因此,掌握同域分布物种的生态位在时间和空间上的分化与重叠特征有助于理解同域分布物种的种间关系(包括竞争、捕食等)及其共存机制,为生物多样性保护与管理提供建议[5]。红外相机监测技术能够实现全天候24 h监测,已成为研究同域物种共存机制的有效手段[6-7],目前已被广泛应用于动物集群、活动节律和同域物种的时间生态位分化等方面的科学研究中[8-9]。
经济发展是造成人与野生动物冲突的直接原因[10]。大熊猫(Ailuropodamelanoleuca)是我国一级重点保护野生动物,同时作为旗舰物种和伞护物种,还面临着许多挑战,快速的人口增长及逐渐加剧的人类活动,对大熊猫的生存与繁衍产生严重影响[11-12]。已有研究表明,野生大熊猫性成熟较晚,繁殖率较低,而幼仔死亡率较高[13-14]。秦岭地区的雌性和雄性大熊猫性成熟年龄都是4.5岁,幼仔年出生率为0.654仔/a,幼仔的1岁存活率仅为59.5%[15]。由于初生幼仔发育极不完善,且母体大熊猫在育幼期间存在频繁更换育幼洞穴的现象,对幼仔存活可能是极大挑战[14]。因此,处在幼年期的大熊猫更易受到捕食者的威胁。
在秦岭地区与大熊猫同域分布的食肉动物中,金钱豹(Pantherapardus)、黄喉貂(Martesflavigula)和豹猫(Prionailurusbengalensis)均可对大熊猫的亚成体及幼仔构成威胁[16],但有关秦岭大熊猫与这些食肉动物的共存模式及生态位分化鲜有报道。本研究基于红外相机监测数据分析了陕西佛坪国家级自然保护区内的大熊猫及其主要潜在捕食动物金钱豹、黄喉貂和豹猫的日活动节律和空间共存特征,探讨这些物种的共存机制,以期为秦岭大熊猫及同域分布濒危物种的保护与管理提供科学依据。
1 研究区概况
陕西佛坪国家级自然保护区位于秦岭中段南麓陕西省佛坪县境内(33°33′—33°46′ N,107°41′—107°55′ E),总面积29 240 hm2,周边与陕西观音山、长青、周至、老县城和黄柏塬5个国家级自然保护区相连,处于秦岭自然保护区群中心位置。佛坪保护区建立于1978年,是以保护大熊猫为主的森林和野生动物类型国家级自然保护区,区内生物多样性丰富,有野生脊椎动物32目101科262属452种,其中兽类7目26科58属73种,鸟类17目54科153属316种[17]。根据第四次全国大熊猫调查,佛坪保护区分布有野生大熊猫成体67只,野外种群密度0.23只/km2,是秦岭地区野生大熊猫种群数量最多、密度最大的自然保护区[16]。
2 研究方法
2.1 红外相机布设
2017年1月—2019年12月,在佛坪保护区内大熊猫高密度分布区(核心区和缓冲区)28条巡护样线上布设了102台红外相机(型号为Ltl6210MC),即102个相机监测位点。每条样线上安装1到数台不等的红外相机,若1条样线上安装2台以上的红外相机,相机间距在500 m以上,并在海拔1 500~2 100 m大熊猫产仔育幼洞穴的集中分布区[18-19]适当增加相机布设位点(图1)。红外相机主要安放于大熊猫活动通道上及标记树附近的树干上,相机参数统一设定,拍摄数量为连拍3张,拍摄时间间隔为1 s,灵敏度为中等。相机安装好后,记录安装日期、时间、相机编号、GPS位点、海拔、坡度、坡向、水源距离和植被状况等信息。每隔3~4个月收取1次照片数据,检查并更换电池、存储卡以确保相机正常工作。
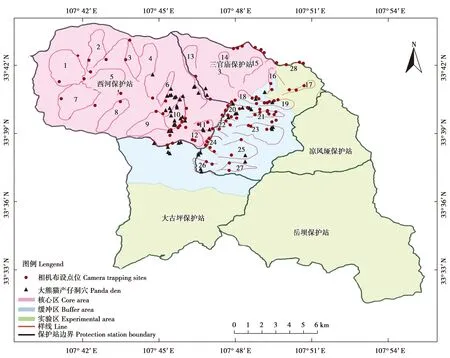
图1 佛坪国家级自然保护区红外相机布设位点Fig.1 Locations of infrared cameras in the Foping National Nature Reserve
2.2 数据分析
2.2.1 日活动节律及时间重叠性
所有分析均基于各物种的有效探测次数,即同一相机位点含同一物种的相邻有效照片间隔时间至少为30 min[20]。根据物种活动时间与日出日落的关系,分为昼间活动(主要在日出后1 h—日落前1 h)、夜间活动(主要在日落后1 h—日出前1 h)和晨昏活动(主要在日出日落前后1 h内)[21]。
2.2.2 空间重叠性
使用ArcGIS v10.2软件中的核密度空间分析功能对大熊猫、金钱豹、黄喉貂和豹猫的红外相机拍摄率进行空间分析,绘制各物种相机拍摄率空间分布格局图。将大熊猫的相机拍摄率空间分布格局与天敌动物金钱豹、黄喉貂和豹猫的相机拍摄率空间分布格局叠加,得到各物种高拍摄率区域空间重叠格局,利用ArcGIS空间统计学功能计算各物种间相机拍摄率的空间相关系数。
3 结果与分析
2017年1月—2019年12月,佛坪自然保护区内安放的102个红外相机位点的累计工作日达到52 835 d,其中97个相机位点拍摄到大熊猫,25个位点拍摄到金钱豹,79个位点拍摄到豹猫,85个位点拍摄到黄喉貂。
3.1 日活动节律
调查期间,大熊猫、金钱豹、黄喉貂和豹猫的有效探测次数分别为1 424、97、735、675次。大熊猫的日活动节律以昼行性为主,在夜间也有一定活动强度(图2A);金钱豹(图2B)和黄喉貂(图2C)均为昼间活动模式,而豹猫为典型夜行性动物(图2D)。

图2 大熊猫、金钱豹、黄喉貂和豹猫的日活动节律雷达Fig.2 Radial plots of diurnal activity patterns for Ailuropoda melanoleuca,Panthera pardus,Martes flavigula,and Prionailurus bengalensis
3.2 时间重叠特征

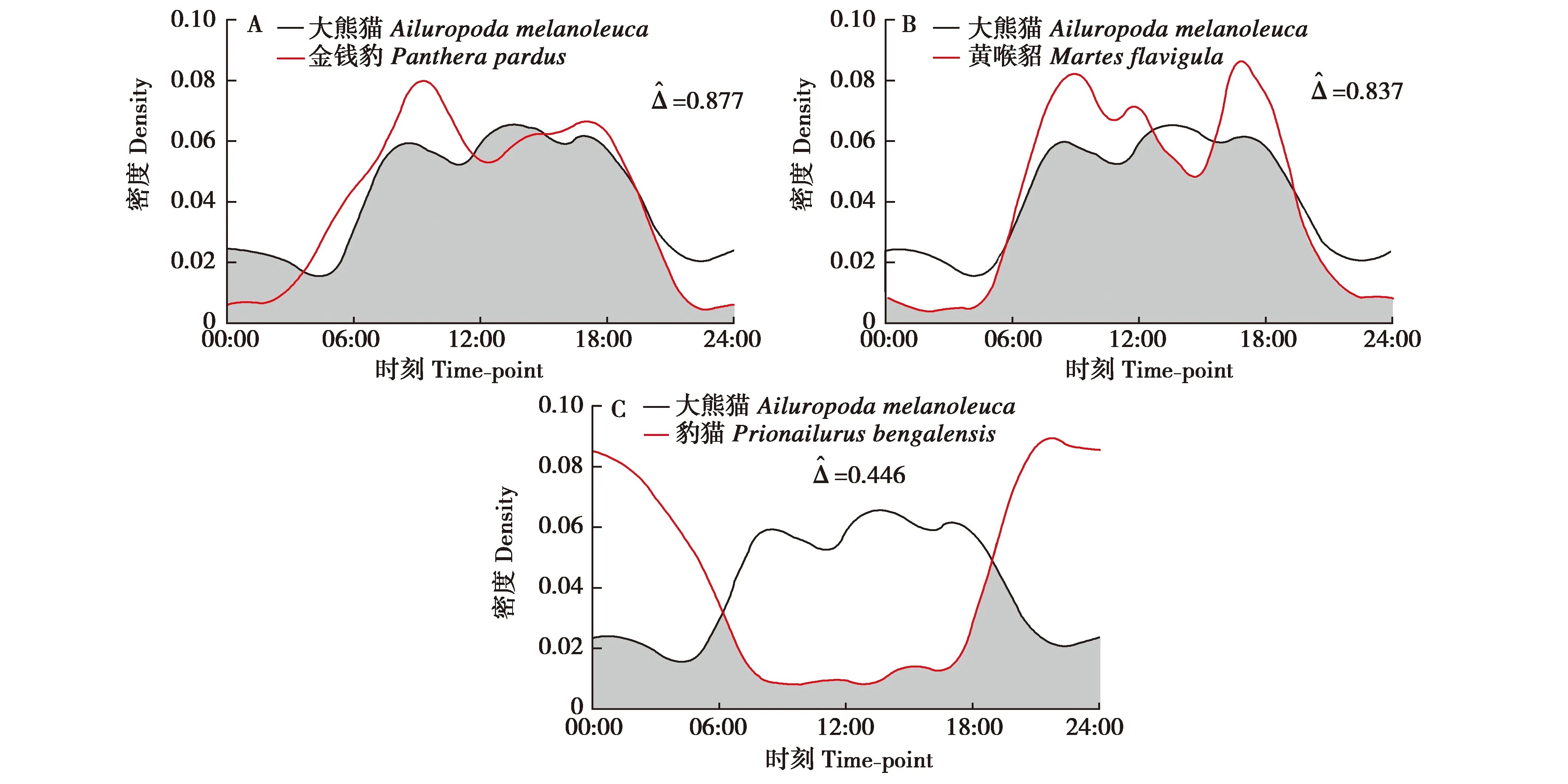
图3 大熊猫与金钱豹、黄喉貂、豹猫的日活动节律重叠分析Fig.3 Overlapping of the diurnal activity patterns between Ailuropoda melanoleuca and Panthera pardus,Martes flavigula,and Prionailurus bengalensis,respectively
3.3 空间重叠特征
图4所示为大熊猫的相机拍摄率空间格局及其与3种食肉动物的相机拍摄率空间关联特征。大熊猫主要活动于佛坪保护区核心区和缓冲交界的三官庙和西河保护站,2个保护站各有1个最高核密度区(图4A)。比较大熊猫与金钱豹核密度重叠情况,大熊猫和金钱豹的相机拍摄率空间相关系数为-0.09,核密度最高的区域亦不重叠(图4B),说明两者的核心活动区域趋向于分离。大熊猫核密度最高的区域在海拔1 500~1 900 m的冬居地,而金钱豹核密度最高的区域在保护区西北部的黄桶梁和北部的光头山高海拔地区,即大熊猫的夏居地。黄喉貂和豹猫的相机拍摄率与大熊猫的相机拍摄率的空间相关系数均大于0.70,说明与大熊猫的活动空间高度重叠。黄喉貂高密度区范围更广,几乎覆盖了所有红外相机安装点,且其高核密度区与大熊猫高核密度区全部重合,即在大熊猫的冬居地和大熊猫洞穴集中分布区(图4C)。类似地,豹猫和大熊猫核密度区大部分重叠即处在西部大熊猫高核密度区部分完全重叠,处在东部大熊猫高核密度区则部分重合(图4D)。
4 讨论
同域物种生态位的分化是多维度的,而行为活动的时间差异常常是共存物种的生态位分化形式之一[23-24]。活动节律既是物种本身进化适应的一部分,也是对环境因素变化采取生存对策的一种适应,在长期的自然选择中,每一个物种都形成了利于自己的最适活动节律[25]。在本研究中,金钱豹主要在昼间活动,偏向于晨昏活动模式,这与通常认为金钱豹常在夜间活动相左。虽然该结果有待进一步验证,已有研究显示豹为昼间活动,与虎(Pantheratigris)的活动节律形成时间活动上的分离[26-27],而历史上秦岭地区也有华南虎(Pantheratigrisamoyensis)分布。虽然自20世纪60年代后再未发现华南虎的踪迹,但秦岭地区的金钱豹在长期自然选择中已形成了利于自己的昼间活动节律并保留至今。金钱豹和黄喉貂与大熊猫的日活动节律重叠指数很高,说明两者对白天活动的大熊猫具有较大的潜在威胁。豹猫为典型夜间活动模式,与大熊猫的日活动节律重叠指数较低,但大熊猫在夜间也有一定活动强度,豹猫对大熊猫幼仔构成较大威胁。
物种间空间分布重叠分析结果显示,大熊猫与金钱豹空间分布的重叠程度很低,结合两者的日活动节律重合程度较高,反映出金钱豹和大熊猫通过空间生态位的分化实现共存。金钱豹高核密度区在保护区西北部的黄桶梁和北部高海拔地区的光头山,即大熊猫的夏居地。黄桶梁地区处于中山和亚高山地带,森林植被主要为针阔叶混交林、落叶阔叶混交林和亚高山针叶林带,其下优势灌木多为秦岭箭竹,在山脊附近和沟谷秦岭箭竹林生长稀疏,草本植物茂盛;光头山地区海拔较高,森林植被为亚高山针叶林带和高山灌丛草甸带[17],有利于金钱豹的活动与捕食。大熊猫高核密度区处于冬居地,每年9月至翌年5月长达9个月时间内,秦岭野生大熊猫主要活动在海拔2 200 m以下的竹林中,在秋冬季和初春主要采食落叶阔叶林和针阔叶混交林下巴山木竹(Bashaniafargesii)叶和茎[28-29],这里植被生长茂密,有利于大熊猫的觅食和躲避大型天敌。作为秦岭地区最大的食肉动物,金钱豹在高海拔区域与大熊猫共同生活栖息约4个月,是对大熊猫及其幼仔构成威胁的主要物种。研究中发现,金钱豹有时也会游荡在大熊猫冬居地,偶尔也会探访大熊猫产仔育幼洞穴,对新生大熊猫幼仔和迁移中的大熊猫幼仔威胁更大。
佛坪保护区野生动物种类丰富,许多野生动物相对多度指数都较高[30],为食肉动物提供了丰富的食物来源。根据最优取食理论分析,动物在食物量资源丰富时选择最优食谱[31]。黄喉貂和豹猫体型大小相近,这就促使两者在空间资源利用上的重叠,却又以时间生态位的分化巧妙缓解竞争,从而实现两者共存。在食物充足的条件下,食肉动物首先捕食自保能力相对较弱的啮齿目(Rodentia)、偶蹄目(Artiodactyla)和鸟类等动物,大熊猫这种大型兽就成为次要选择。据报道,2014年四川唐家河国家级自然保护区有1只3岁大熊猫疑遭黄喉貂围攻致肠子外露[32]。大熊猫高核密度区与黄喉貂高核密度区在大熊猫冬居地即大熊猫产仔育幼洞穴区域全部重合,且黄喉貂高核密度区范围更大几乎覆盖了所有大熊猫分布区,两者存在高度竞争关系。黄喉貂生态位更为广泛,适应性更强,又是群居动物,因此,黄喉貂是佛坪保护区大熊猫同域捕食者中最大的潜在威胁。豹猫高核密度区和大熊猫高核密度区大部分重叠,主要在大熊猫冬居地活动,豹猫冬春季节对大熊猫及其幼仔威胁大于其他季节。
大熊猫与金钱豹等3种食肉动物通过在时间生态位或空间生态位上的一定程度的分化达到共存,实现同域分布。大熊猫与黄喉貂的时空生态位重叠程度较高,是佛坪保护区大熊猫同域捕食者中最大的潜在威胁。在今后大熊猫的保护研究中应对处于食物链顶级的金钱豹及黄喉貂、金猫(Catopumatemminckii)、豹猫等主要捕食者和捕食对象深入研究,以获得更全面的物种共存信息。本研究有助于综合评价秦岭野生大熊猫潜在捕食风险,为野外大熊猫保护、繁殖和野外放归提供有效的科学管理意见。
致谢:感谢陕西佛坪国家级自然保护区管理局各保护站职工在野外工作中的支持,以及陕西师范大学叶新平教授在数据处理中给予的帮助。
