一种松材线虫醛脱氢酶的基因克隆及其生化性质
2022-11-05李文硕王林松杜桂彩郭群群张廷婷杨宏李荣贵
李文硕 王林松 杜桂彩 郭群群 张廷婷 杨宏 李荣贵
(青岛大学生命科学学院,青岛 266071)
由松材线虫(Bursaphelenchus xylophilus)引起的松萎蔫病已经在世界各地造成严重的生态环境破坏和巨大的经济损失,是松树的一种毁灭性灾害[1]。中国于1982年在江苏省南京市首次发现了松萎蔫病,随后该病迅速扩散至其他省份[2]。该病的发生和传播涉及多种因素,如松树的抗性、昆虫传播媒介及线虫携带的细菌等[3]。松萎蔫病的致病机制尚不清楚,目前尚未开发出针对该病有效且廉价的药物[3]。
当松材线虫感染松树的木质部时,树木的受损细胞可以释放化学物质来刺激模式识别受体并激活寄主松树的防御系统,使松树产生增厚细胞壁,产生活性氧,并积累醇、醛和萜烯等次生代谢产物现象[4-6]。松材线虫以松树的管胞和射线细胞为食,并依靠体内的酶类抵御入侵宿主过程中遇到的防御阻碍[7-8]。松材线虫体内各种酶类的研究已经成为松萎蔫病致病机制研究的热点,过氧化物酶是松材线虫应对寄主防御反应重要酶类,该酶由线虫口针分泌至体外发挥作用,降低由寄主防御系统产生的活性氧对松材线虫的伤害[9]。Fu等[10]克隆了一个在松材线虫尾部表达的Bx-Prx基因,该基因表达的产物同属于过氧化酶类,通过RNA干扰Bx-Prx的表达可以降低松材线虫的繁殖率。细胞色素P450通路被认为是松材线虫的主要解毒途径[3,11-13],干扰色素 P450家族 中 CYP33C9、CYP33C4、CYP33D3三个基因的表达,松材线虫的活性和致病性均显著降低[14]。
在之前的研究中,为了探究松材线虫体内一种醇脱氢酶的生理功能,我们使用醇脱氢酶的特异性抑制剂甲吡唑处理松材线虫,结果显示松材线虫的取食能力和运动能力均明显降低,表明醇代谢途径对松材线虫生长发育发挥着重要作用[15]。醛脱氢酶是醇类物质代谢途径中重要的一环,承担着将毒性较大的醛类转化为酸的重要功能[16]。转录组分析显示,甲吡唑处理的松材线虫,其体内一种编码醛脱氢酶的基因表达水平显著升高。本文通过研究该醛脱氢酶的理化性质,为深入了解该酶在松萎蔫病病理进程中的作用打下基础,也为以醛脱氢酶为靶点的杀线药物的开发提供参考。
1 材料与方法
1.1 材料
野生松材线虫取自崂山(中国,青岛市)上的枯萎黑松,将松枝剪成1 cm3左右的小块,采用贝尔曼漏斗法分离野生松材线虫[15]。将分离出的线虫接种于长满灰葡萄孢的马铃薯淀粉琼脂培养基中,在25℃下避光培养一周左右[17]。本实验所用的大肠杆菌DH5α和BL21(DE3)菌株及表达载体pET-15b购自Novagen公司并由本实验室保存。实验所用的限制核酸内切酶和T4 DNA连接酶购自NEB公司(北京);Taq DNA聚合酶、琼脂糖凝胶 DNA 回收试剂盒、质粒提取试剂盒购自天根生化科技有限公司(北京);pMD-18T载体、RNA提取试剂RNAiso购自宝日医生物技术(北京)有限公司;乙醛购自上海阿拉丁生化科技股份有限公司;烟酰胺腺嘌呤二核苷酸(NAD)购自上海源叶生物科技有限公司;其他化学试剂为分析纯级别,购自上海沪试实验室器材股份有限公司;引物合成和质粒测序工作由上海生工公司完成。
1.2 方法
1.2.1 松材线虫总RNA的提取及cDNA的合成 使用RNAiso试剂从松材线虫中提取总RNA,并用1%的琼脂糖凝胶电泳分析。以松材线虫总RNA模板,按照宝日医生物技术(北京)有限公司cDNA第一链合成试剂盒说明书操作,反转录mRNA生成cDNA第一链。
1.2.2 目的基因克隆及表达载体构建 根据转录组的测序结果,分别设计一对带有Nde I和BamH I酶切位点的特异引物(引物序列见表1),以松材线虫cDNA第一链为模板按如下程序扩增目的基因:94℃预变性 5 min;94℃变性 50 s,57℃退火 40 s,72℃延伸90 s,共进行35个循环。PCR产物通过1%琼脂糖凝胶电泳分离,用琼脂糖凝胶 DNA 回收试剂盒回收目的基因,16℃下与pMD-18T载体连接过夜,连接产物转化E. coli DH5α,在含有氨苄青霉素(100 μg/mL)LB培养基筛选阳性克隆,挑取单克隆培养后提取质粒并测序,序列正确的质粒命名为pMD-18T-aldh。

表1 本试验所用引物Table 1 PCR primers used in this study
pMD-18T-aldh经Nde I/BamH I双酶切,琼脂糖凝胶电泳分离载体pMD-18T与aldh,用琼脂糖凝胶 DNA 回收试剂盒回收aldh。用NdeI/BamHI双酶切表达载体pET-15b后,经琼脂糖凝胶电泳分离后,用琼脂糖凝胶 DNA 回收试剂盒回收pET-15b。回收的载体pET-15b和aldh混合后用T4DNA连接酶于16℃下连接过夜,连接产物转化E. coli DH5α,在含有氨苄青霉素(100 μg/mL)LB培养基筛选阳性克隆,挑取单克隆培养后提取质粒并测序,序列正确的质粒命名为pET-15b-aldh。
1.2.3 醛脱氢酶结构预测 醛脱氢酶的三级结构通过在线网站http:// www.sbg.bio. ic. ac.uk/ phyre2 /html/page..cgi?id=index预测。通过在线网站https:// www.novopro.cn /tools/ signalp.html预测醛脱氢酶是否含有信号肽。利用NPS@ SOPMA工具分析重组醛脱氢酶的二级结构。
1.2.4 表达及重组蛋白纯化 将测序正确的表达载体pET-15b-aldh转入大肠杆菌BL21(DE3)感受态细胞中,在含有氨苄青霉素(100 μg/mL)LB培养基筛选阳性克隆,阳性克隆即为构建的工程菌,挑取单菌落于37℃、150 r/min 的摇床中培养过夜。按1.5%(体积比)的接种量将培养液接种至4 L LB液体培养基中(含100 μg/mL 氨苄青霉素)进行菌体扩大培养。当OD600等于1时加入β-异丙基硫代半乳糖苷(麦克林,上海)至终浓度为0.4 mmol/L 诱导目的蛋白表达,28℃诱导4 h。4℃,8 000×g,离心20 min收集菌体。菌体用结合缓冲溶液(20 mmol/L Tris-Cl,pH 8.3,5 mmol/L 咪唑,500 mmol/L NaCl)悬浮后,用超声波破碎细胞,4℃,12 000×g,离心30 min获得粗酶液。Ni-NTA树脂用平衡缓冲液平衡处理后装柱(1.2 cm×5.0 cm),加入粗酶液,然后用2种含有不同浓度咪唑的洗涤缓冲液进行逐步洗脱杂蛋白(洗涤缓冲液1:20 mmol/L Tris-Cl,pH8.3,30 mmol/L 咪唑,500 mmol/L NaCl;洗涤缓冲液 2 :20 mmol/L Tris-Cl,pH 8.3,63 mmol/L 咪唑,500 mmol/L NaCl),最后用洗脱缓冲液(20 mmol/L Tris-Cl,pH8.3,500 mmol/L 咪唑,500 mmol/L NaCl)洗脱目的蛋白,4℃条件下洗脱蛋白溶液对Tris-Cl缓冲液(10 mmol/L,pH 8.0)透析脱盐。用考马斯亮蓝法测定蛋白浓度,并用分离胶浓度为12% 的SDS-PAGE 分析纯化结果[18]。
1.2.5 重组醛脱氢酶米氏常数的测定 在25℃下,以甲醛为底物测定重组醛脱氢酶的米氏常数。反应根据Okibe的方法稍作更改,反应总体积为 2 mL,含 5 mmol/L Tris-Cl 缓冲液(pH 8.3),3 mmol/L MgCl2,10 mmol/L 氧化型NAD和0.32 mg重组醛脱氢酶,以及不同浓度的甲醛(15 mmol/L-40 mmol/L)[19]。反应时间10 min,通过测定反应体系在340 nm 吸光度变化求出反应速度,采用双倒数作图法求出醛脱氢酶的Km值,还原型NADH在340 nm 的毫摩尔消光系数为6.22 mmol/L/cm[20]。使吸光度增加0.01所需的酶量定义为1 U。
1.2.6 不同pH值对醛脱氢酶反应速度的影响 在不同pH值下分别测定醛脱氢酶反应速度。在25℃下,反应时间5 min。采用如下6种不同缓冲液体系创造不同的pH环境:0.2 mol/L NaAC-HAC缓冲液(pH5.5);0.2 mol/L Na2HPO4-NaH2PO4缓冲液(pH6.5);0.2 mol/L Tris-Cl缓冲液(pH7.5);0.2 mol/L Tris-Cl缓冲液(pH8.5);0.2 mol/L Gly-NaOH 缓冲液(pH9.5);0.2 mol/L Gly-NaOH缓冲液(pH10.5)。反应总体积为2 mL,向上述不同的反应缓冲液分别加入0.3 mmol/L KCl、5 mmol/L 氧化型 NAD、50 mmol/L 乙醛和0.63 mg重组醛脱氢酶。在340 nm下测定反应体系吸光度的变化,计算反应速度。
1.2.7 温度对醛脱氢酶反应速度的影响 在不同温度条件下(15℃-50℃)分别测定醛脱氢酶的反应速度。反应时间为5 min的反应体系含有0.2 mol/L Na2HPO4-NaH2PO4缓冲液(pH6.5),0.3 mmol/L KCl,5 mmol/L 氧化型NAD,50 mmol/L 乙醛和0.63 mg重组醛脱氢酶,反应总体积为2 mL。在340 nm下测定反应体系吸光度的变化,计算反应速度。
1.2.8 金属离子对醛脱氢酶反应速度的影响 在25℃下,在5 mmol/L Tris-Cl缓冲液(pH8.3)中分别加入10 mmol/L 氧化型NAD,0.32 mg重组醛脱氢酶,35 mmol/L 甲醛,及终浓度为3 mmol/L 不同金属离子(Mn2+、Na+、Ca2+、Ni2+、K+、Fe3+),反应体系为2 mL,反应时间5 min。在340 nm下测定反应体系吸光度的变化,计算反应速度。
1.2.9 醛脱氢酶对不同底物的催化能力 在25℃下,在5 mmol/L Tris-Cl缓冲液(pH8.3)中,测定以不同醛类为底物时酶的反应速度,用于测试的醛类有:香草醛、戊二醛、甲醛、乙醛和4-(二甲氨基)-苯甲醛(简写为PDAB)。反应总体积为2 mL,反应时间5 min,反应体系中分别加入终浓度为35 mmol/L上述不同的醛,10 mmol/L 氧化型NAD和0.32 mg重组醛脱氢酶,3 mmol/L MgCl2。在340 nm下测定反应体系吸光度的变化,计算反应速度。
2 结果
2.1 目的基因的克隆及表达载体的构建
使用RNAiso试剂成功从松材线虫中提取了总RNA并反转录获得了cDNA第一链。以此为模板通过RT-PCR成功扩增出一段约1.3 kb的片段,将该片段连入pMD-18T构建pMD-18T-aldh。测序结果显示,扩增得到的片段与GenBank序列号为CAD5152421.1的序列一致。该片段有完整的开放阅读框,大小为1 353 bp,编码451 个氨基酸残基(图1),根据EXPASY 分析,醛脱氢酶理论大小约为51 kD,理论等电点(pI)为4.8。
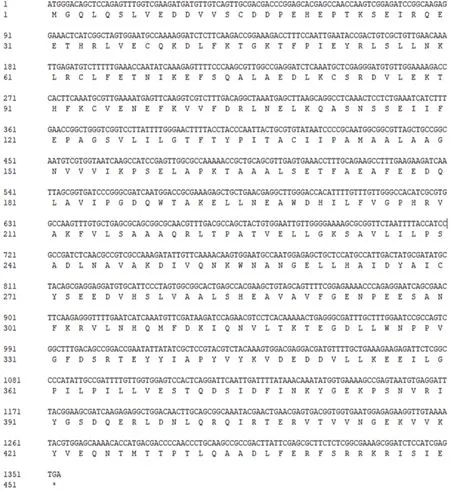
图1 松材线虫aldh基因序列和推测的氨基酸序列Fig.1 Sequences and predicted amino acid sequences of gene aldh from PWN(pine wilt disease)
2.2 重组醛脱氢酶的结构预测
使用在线工具预测得到的重组醛脱氢酶的三维结构如图2-A所示。通过NPS@ SOPMA工具分析,结果如图2-B显示,醛脱氢酶二级结构有较多的α螺旋结构(蓝色),占47.33%,其次是紫红代表的无规卷曲占29.78%。利用在线工具预测重组醛脱氢酶信号肽,重组醛脱氢酶不具有信号肽结构。
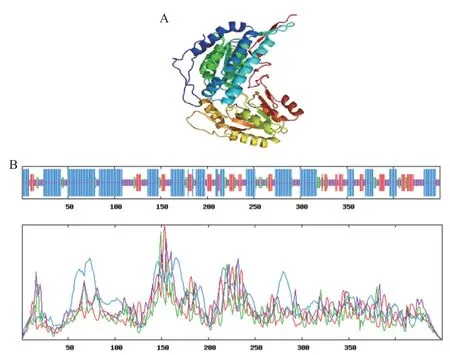
图2 醛脱氢酶三维结构(A)和二级结构预测(B)Fig.2 Prediction of the 3-dimensional structure(A)and 2 -dimensional structure(B)of aldehyde dehydrogenase
2.3 重组醛脱氢酶的表达及纯化
转化表达载体pET-15b-aldh的大肠杆菌BL21(DE3)经IPTG诱导后积累大量目的蛋白(图3),经SDS-PAGE分析,结果显示,利用镍柱亲和层析纯化出一种相对分子量约为51 kD的蛋白,与aldh编码的醛脱氢酶的理论大小一致。
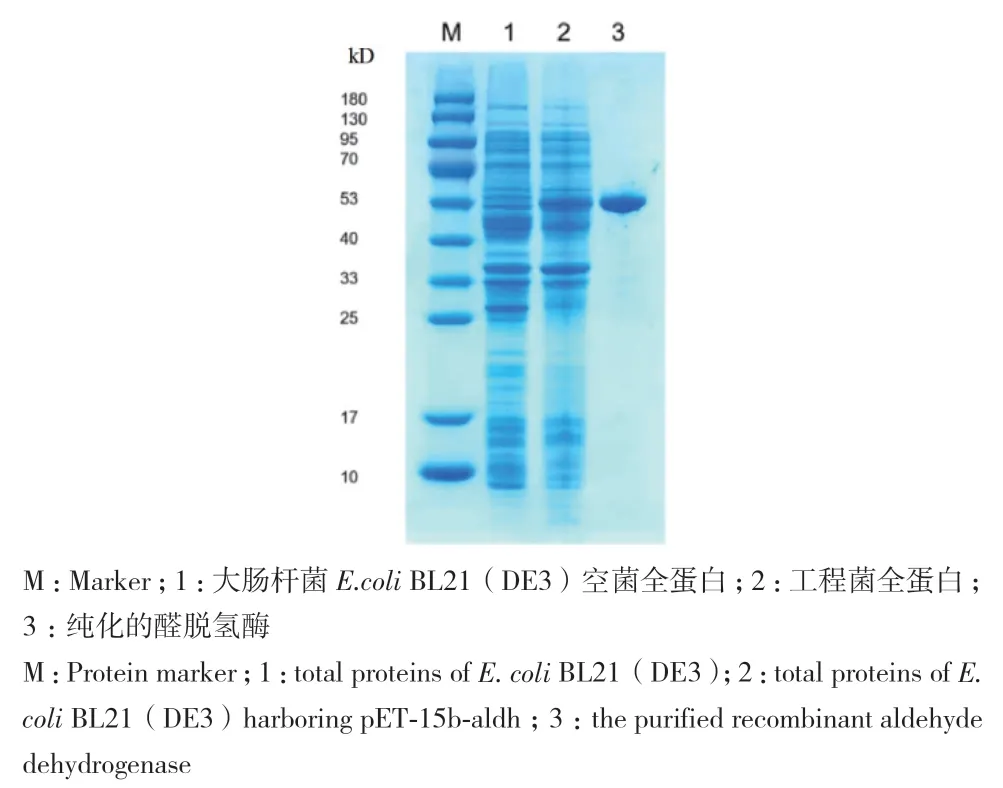
图3 重组醛脱氢酶的表达与纯化Fig. 3 Expression and purification of the recombinant alde-hyde dehydrogenase
2.4 重组醛脱氢酶米氏常数的测定
以反应速度的倒数(1/v)对底物浓度的倒数(1/[s])作图,如图4所示,根据Lineweaver-Burk方程式求得醛脱氢酶的Km值。当用甲醛为底物,氧化型NAD为辅因子时,Km为27.87 mmol/L。
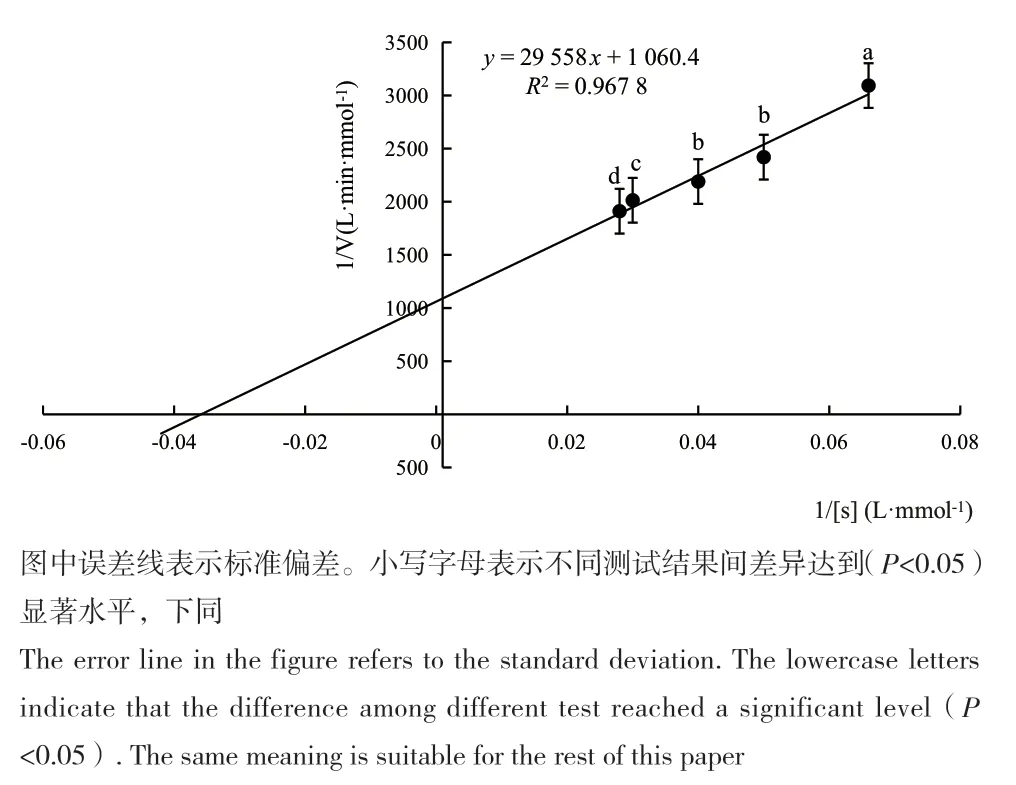
图4 通过Lineweaver-Burk计算醛脱氢酶的Km值Fig. 4 Km of the recombinant aldehyde dehydrogenase calculated by Lineweaver-Burk plot
2.5 pH对重组醛脱氢酶反应速度的影响
反应体系的pH值对醛脱氢酶反应速度的影响如图5所示,pH值的变化范围从5.5-10.5,重组醛脱氢酶在不同环境中表现出了不同反应速度,从图中可知重组醛脱氢酶的最适pH值为7.5,当pH值为10.5时,醛脱氢酶检测不到催化活性。
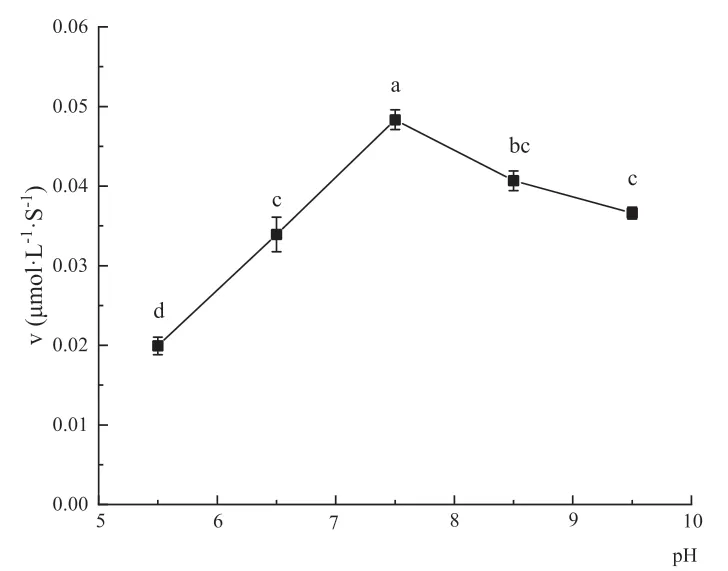
图5 不同pH值对醛脱氢酶反应速度的影响Fig. 5 Effects of pH values on aldehyde dehydrogenase reaction rates
2.6 不同温度对醛脱氢酶反应速度的影响
温度对醛脱氢酶催化活力影响较大,当测试温度范围为15℃-50℃,由图6可知重组醛脱氢酶的最适温度为25℃,当反应温度达到50℃时,已检测不到催化活性。
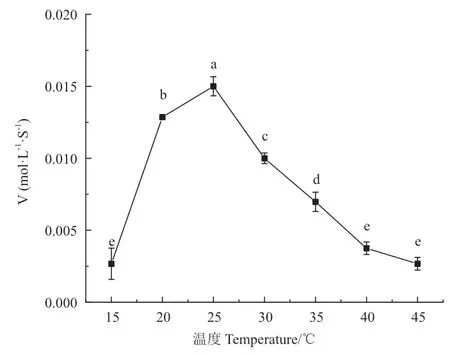
图6 不同温度对醛脱氢酶活性的影响Fig. 6 Effects of temperatures on aldehyde dehydrogenase activity
2.7 金属离子对醛脱氢酶反应速度的影响
向测试体系分别添加终浓度为3 mmol/L的不同的金属离子,与完全不添加金属离子相比,Fe3+和Ni2+能够提高酶的活力,Mn2+、Na+、K+和Ca2+对重组醛脱氢酶的活性均有抑制作用(图7)。
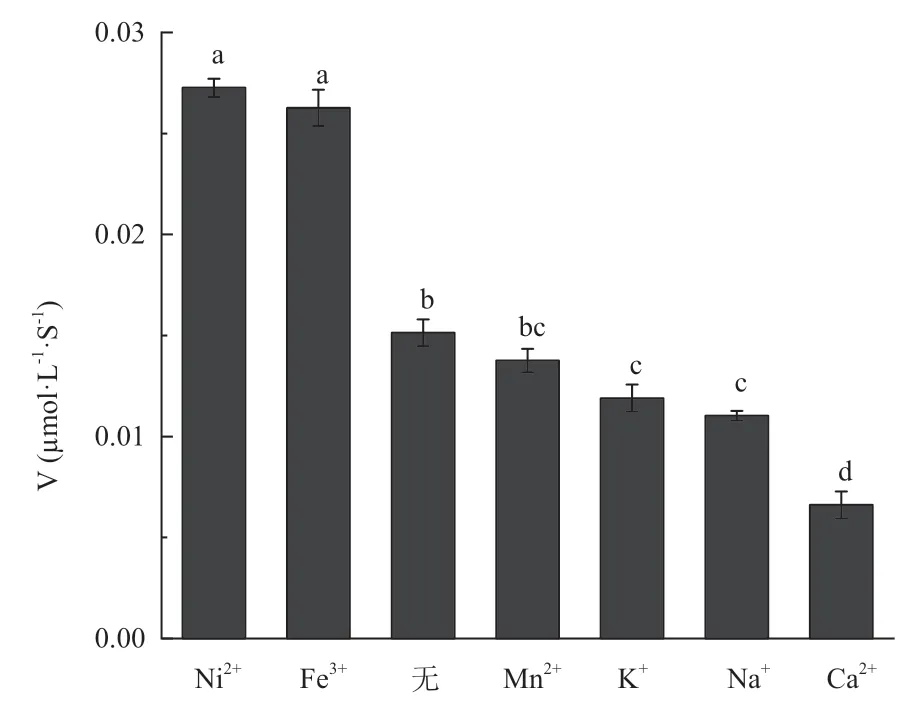
图7 不同金属离子对醛脱氢酶反应速度的影响Fig. 7 Effects of metal ions on aldehyde dehydrogenase reaction rates
2.8 醛脱氢酶对不同醛类的催化能力
在供试的5种醛类中,重组醛脱氢酶对香草醛显示了较高的催化活力(图8),酶活力为(4.06±0.20)U,对乙醛的催化活性最低,酶活仅为(0.866±0.12)U。对结构复杂的芳香族醛类4-(二甲氨基)-苯甲醛的没有检测到催化活性。
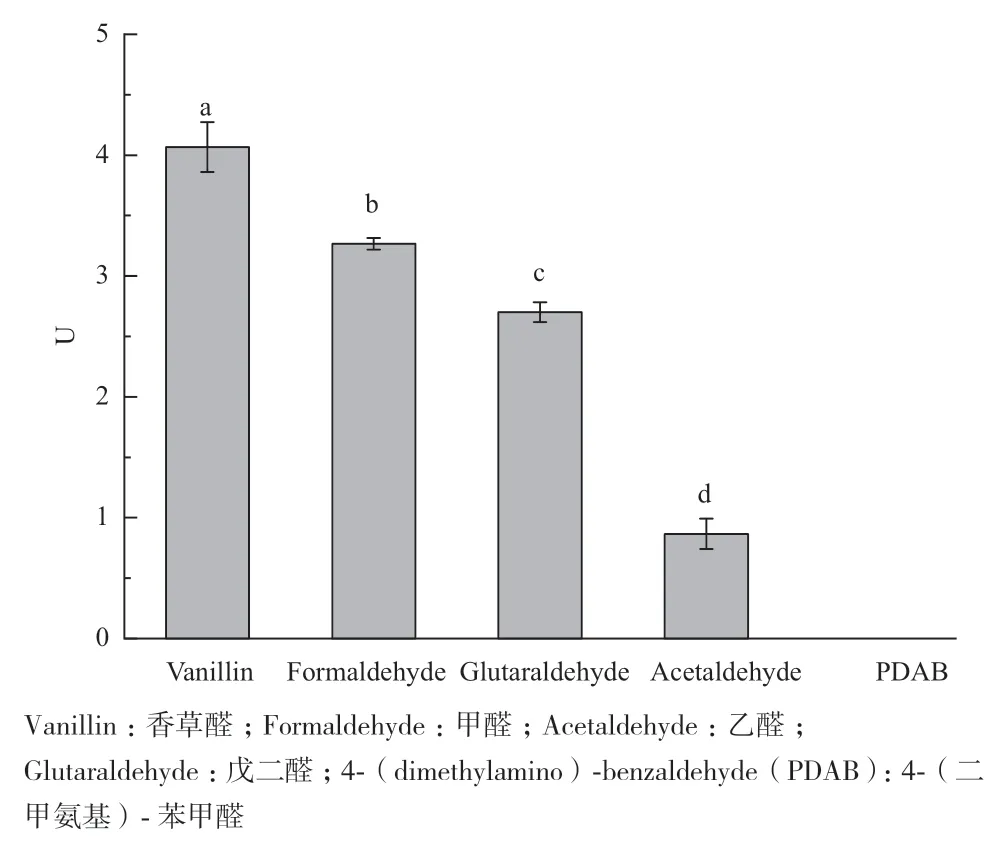
图8 醛脱氢酶对不同醛类的催化活性Fig. 8 Catalyzed activities of aldehyde dehydrogenase on different aldehydes
3 讨论
松树对松材线虫的抗性与其体内次生代谢物有关。马尾松种源的抗性与α-松油醇、长叶烯、雪松烯和金合欢烯等成分呈正相关[21]。马尾松富含松醇,其含量也与马尾松的抗逆性密切相关[22]。松材线虫侵染黑松后,宿主产生的挥发性成分中检测到乙醇和萜类化合物[23]。已有研究表明,松树产生的挥发物反式-2-己烯醛有较好的杀线活性[24],松树正常的脂类代谢也会产生丙二醛。松材线虫在入侵宿主的过程中,需要克服宿主产生的这些醇类、醛类等次生代谢产物,才能成功定殖并致病。前期我们研究发现,松材线虫的醇脱氢酶能够催化多种醇类化合物的脱氢反应生成醛类化合物[15],但无论宿主体内还是松材线虫代谢产生的醛类物质仍然具有较大毒性,需要进一步氧化为酸类成分才能进一步解毒和利用,推测松材线虫体内的醛脱氢酶就有此种功能,因此,松材线虫醛脱氢酶可能成为松萎蔫病防治的一个潜在靶点,但目前尚未有松材线虫醛脱氢酶的研究报道。
通过生物信息学方法,我们在GenBank中发现了13个与醛脱氢酶的编码基因高度同源的序列(未公开结果),本文通过RT-PCR的方法从松材线虫体内成功克隆出一个编码醛脱氢酶的cDNA,并成功构建了表达载体。转入大肠杆菌BL21(DE3)中构建了工程菌,IPTG诱导该基因的高效表达,并利用亲和层析的方法纯化获得该基因编码的醛脱氢酶。活性分析显示,以甲醛为底物,以氧化型NAD为辅酶,通过双倒数作图法求得醛脱氢酶的Km值为27.87 mmol/L,与其他使用NAD作为辅因子的醛脱氢酶相比,来自太平洋火色杆菌(Flammeovirga pacifica)的一种醛脱氢酶,其Km值为(421±23.5)μmol/L;大肠杆菌的甘油醛-3-磷酸脱氢酶(C192A)的Km值为(0.074±0.004)mmol/L[25-26],松材线虫的醛脱氢酶具有中等的Km值。在对不同底物的催化活性的实验中,松材线虫的醛脱氢酶表现出了对香草醛较好的催化活性,这可能是松材线虫在松树体内抵御香草醛的重要手段,而香草醛存在于感病樟子松木质部和赤松树皮部位[27-28]。
已有很多研究报道金属离子显著影响醛脱氢酶的活性,嗜热脱氮土壤芽孢杆菌(Geobacillus thermodenitrificans)NG802 的醛脱氢酶可被 Mg2+、Ca2+和Mn2+的激活,但受到Co2+、Cu2+和Ni2+的抑制[29]。本研究发现Fe3+和Ni2+可提高松材线虫醛脱氢酶的活性,而Ca2+、Mn2+、Na+和K+可降低酶活性,这为醛脱氢酶的开发利用打下基础。此外,对该酶的一级结构比对分析发现,该醛脱氢酶内位于催化中心的保守氨基酸残基并不是半胱氨酸(Cys),而是亮氨酸(Leu),但其C-端紧邻Leu残基的是组氨酸残基(His),而不是常见的非极性氨基酸残基(图9),推测His参与了醛类化合物的催化作用。因此,对松材线虫醛脱氢酶的深入研究有助于我们了解它们的理化性质并为松萎蔫病药物的研发提供更多的线索。

图9 多序列对比不同来源的醛脱氢酶的一级结构Fig.9 Multiple sequences alignments of primary structures of aldehyde dehydrogenases from different sources
4 结论
本研究克隆了松材线虫体内一个编码醛脱氢酶的cDNA,获得了重组醛脱氢酶,研究了该醛脱氢酶的生化性质,证实以甲醛为底物的醛脱氢酶Km值为27.87 mmol/L,最适pH值为7.5,最适温度为25℃,Fe3+和Ni2+可提高其酶活性,Ca2+、Mn2+、Na+和K+可降低该酶活性。重组醛脱氢酶对5种测试醛类化合物显示出不同的催化活性,香草醛是其最佳底物。
