上扬子区志留系花瓣海百合动物群的古生物地理学*
2022-11-02毛颖颜李启剑邓小杰史振华
毛颖颜 李启剑 邓小杰 李 越 史振华
(1.中国科学院南京地质古生物研究所现代古生物学与地层学国家重点实验室,中国科学院生物演化与环境卓越创新中心 南京 210008;2.贵州省地质调查院 贵阳 550081)
海百合属于真体腔后口动物,是高等无脊椎动物棘皮动物门中古老的一个纲,也是海洋无脊椎动物中器官复杂、生物多样性极高的纲。海百合化石记录几乎贯穿整个显生宙,迄今全世界已报导逾1 000 余属的海百合化石记录多源于美国和欧洲的数据(Ausich and Kammer,2013;Mao et al.,2017)。海百合是古生代浅海表栖生态群落的重要组成分子,属种多并显示生物地理分区,许多海百合冠部化石层可作为特定区域内划分及对比的标志层;海百合分层滤食以及不同岩相中的生存策略和保存情况都是底质环境的灵敏指示物,具有重要的古生态学意义。中国古生代海百合化石的研究程度偏低,基于冠部器官描述的化石记录仅约90 余属,报导志留纪海百合10 属(穆恩之,1954;田奇㻪等,1957;穆恩之等,1983),其中上扬子区志留纪兰多维列世花瓣海百合动物群以属种级多样性最高、古地理分布最广而最具代表性(Mao et al.,2013,2015,2017)。
花瓣海百合科化石具有极端特化的腕部构造而易于辨识,研究历史长,模式种为产于美国衣阿华州兰多维列统Niagara 灰岩的Petalocrinus mirabilisWeller et Davidson(Weller and Davidson,1896)。Bather(1898)描述瑞典哥特兰岛志留系中的Petalocrinus属7 种,通过对比美国衣阿华州和印第安纳州Hopkinton 白云岩的属种讨论这些种的地层学意义;Norford(1962)描述加拿大卑诗省志留纪Sandpile Group 动物群中的Petalocrinus mirabilisWeller et Davidson。 Moore and Teichert(1978)编 撰 Treatise on Invertebrate Paleontology 中收录美国和瑞典的Petalocrinus化石记录,但未列举已经在中文刊物上发表的华南板块扬子区特有的属SpirocrinusMu et Wu,1974(螺旋海百合)。Witzke et al(.1979)在论述志留纪和泥盆纪棘皮动物的生物古地理时,着重分析花瓣海百合科的层位和分布,认为该科集中产出于兰多维列世晚期到文洛克世中期地层,可作为区域地层划分和对比的标准化石。Prokop(1984)报导花瓣海百合科在捷克泥盆纪早期的孑遗分子VadarocrinusProkop。中国花瓣海百合科化石数量丰富,Mu(1949)最早描述黔北务川志留系石牛栏组灰岩中的花瓣海百合1 属3 种,包括Petalocrinus inferiorBather,P. inflatusMu 和P. chiaiMu。穆恩之等(1974)描述黔东北志留系雷家屯组花瓣海百合科2 属7 种,并建立新属SpirocrinusMu et Wu,该属包含S. typicusMu et Wu 和S. longusMu et Wu 2 个种。Mu et al(.1986)将上扬子区志留系海百合划分出5 个化石组合,其中就包含了黔东北香树园组和雷家屯组的Petalocrinus和Spirocrinus。穆恩之等(1987)描述黔东北石阡地区志留系香树园组和雷家屯组的3 属20 种,包含1 新属SinopetalocrinusMu et Lin(中华花瓣海百合属),并指出扬子区花瓣海百合科化石具有重要的生物地层学对比意义。李罗照(1993)明确花瓣海百合科的最早记录为湖北松滋中奥陶统大湾组的Eopetalocrinus sinensisLi,并推测Eopetalocrinus sinensis和志留纪Spirocrinus的祖—裔演化关系。除扬子区外,属Sibumasu 地块的滇西施甸地区埃隆阶仁和桥组产出全球分布的Petalocrinus mirabilisWeller et Davison(陈重泰等,1993)。Mao et al(.2015)综述全球报导花瓣海百合科的5 属28 种的地点和层位,明确该科化石在兰多维列统(Llandoverian)和文洛克统(Wenlockian)出现频率最高,但仅有Petalocrinus属达到中国、北美、西北欧的全球广泛分布。Mao et al.(2017)记录黔北早志留世Petalocrinus的1 个新种和Spirocrinus的2 新种,依据腕部特化变形、旋转方向等特化方式的成种过程,脉络清晰地复原了上扬子区土著属种的演化谱系,上扬子陆表海区的花瓣海百合动物群属种多样性高居全球之冠,且具有演化过程时间短、分化快以及同域成种之特点。
以特定海区、特定海百合门类的表征地层学特征划分海洋底栖动物生态组合类型,可为解析区域古地理变化、化石分类单元的居群结构、时代延限以及生物—环境的协同演化等理论古生物学关键问题提供直接证据(Halleck,1973;Walker and Diehl,1986;Morris and Felton,1993;Brett et al.,1997)。上扬子陆表海区兰多维列统埃隆阶同期异相的地层单元,即:黔北石牛栏组、黔东北雷家屯组,以及鄂西罗惹坪组均产出花瓣海百合科化石(Mao et al.,2017),各组中化石的属种多样性、丰度以及所赋存岩相差异悬殊,是体现生物—环境协同演化的极好素材。本文聚焦上扬子陆表海区埃隆阶上部赋存花瓣海百合科化石层位的古生物地理学分布特征,选择黔北桐梓水坝塘石牛栏组、黔东北石阡枫香雷家屯组,以及鄂西荆门仙居罗惹坪组为代表剖面(图1),分析不同海底环境中花瓣海百合产出的属种多样性和丰度指标,凸显化石组合在志留纪陆表海不同底质环境和不同生态区间的表现型式,为阐述短暂时限内多元生态因子控制特定底栖群落面貌提供范例。
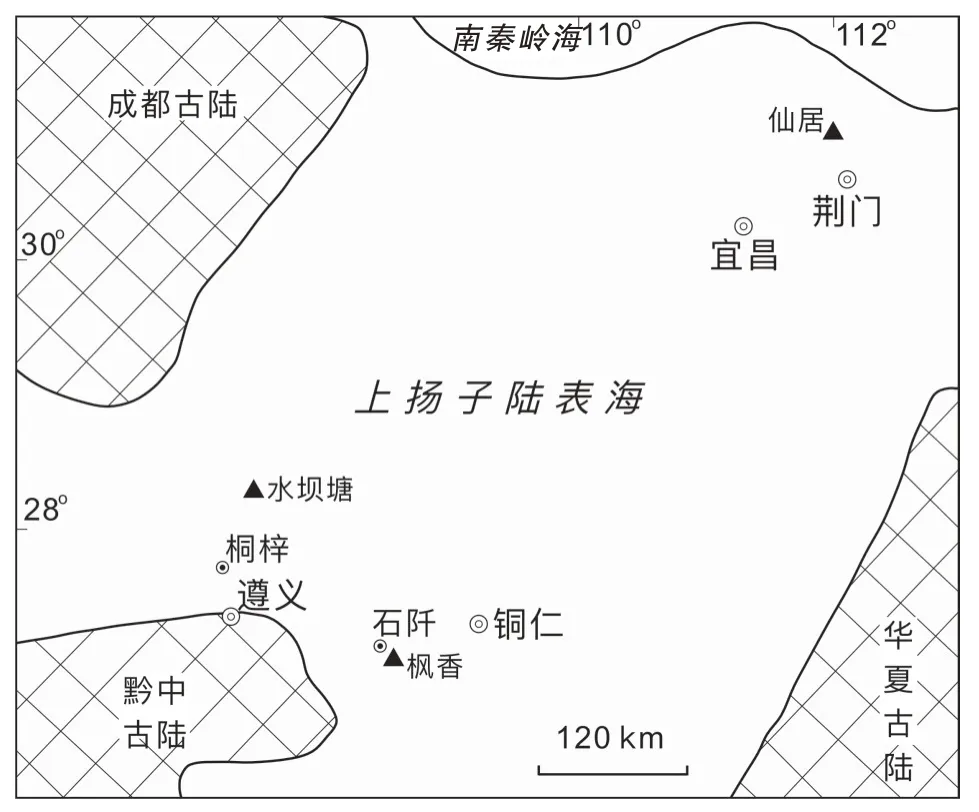
图1 华南板块上扬子陆表海区志留纪兰多维列世埃隆期赋存花瓣海百合化石的3 个剖面位置(底图据Rong et al.,2003)Fig.1 Localities of three Petalocrinidae-bearing sections of Aeronian(Llandovery of Silurian)on the Upper Yangtze epicontinental sea,South China Block(base map after Rong et al.,2003)
1 地质背景和地层
兰多维列世华南板块扬子陆表海区位于低纬度暖水域(Torsvik and Cocks,2013),生物地层学主要基于笔石、牙形石、三叶虫和腕足类化石带进行生物地层学对比(Zhan and Jin,2007)。全球晚埃隆期为高海平面状态(Ross and Ross,1996;Loydell et al.,1998),整个上扬子区陆表海周缘夷平古陆陆源碎屑物供给的减少,引发了碳酸盐岩台地从补丁状拓展为晚埃隆期连片分布,清澈水体有助于生长高生物多样性的浅海群落(陈旭等,1996;Rong et al.,2003;Li et al.,2004);各地层单元生物相—岩相的时空分异明显,碳酸盐岩和碎屑岩含量比例不同,出现多种沉积相区和繁盛的底栖生态单元,底栖壳相动物大化石常见珊瑚、腕足类、层孔虫、头足类、棘皮类等(葛治洲等,1979; 戎 嘉 余 等 , 1984; Li and Kershaw, 2003; Rong et al., 2003; 李 越 , 2004;Wang et al.,2014),石牛栏组的石牛栏段、雷家屯组以及罗惹坪组是埃隆阶上部同期异相的地层单元(戎嘉余等,2019;王怿等,2021)。本文取材花瓣海百合化石在3 个剖面中产出的层位示于图2,各组分述如下。
丁文江(1947)在渝南綦江观音桥定名石牛栏灰岩,其下伏酒店垭页岩,上覆韩家店页岩。尹赞勋等(1947)以含介壳化石的暗灰色页岩作为石牛栏灰岩之底,含海百合茎灰岩和纹层灰岩之上属韩家店黄绿色砂质页岩;现以黄色瘤状泥灰岩出现作为石牛栏群底界(张文堂等,1964;中国科学院南京地质古生物研究所,1974;葛治洲等,1979),可次分石牛栏组为下部薄层瘤状灰岩夹泥岩、粉砂岩的松坎段和上部含丰富壳相化石灰岩夹少量泥岩、粉砂岩的石牛栏段;桐梓县域北部水坝塘镇的石牛栏组厚度122.5 m,石牛栏段主要为后生动物生物礁建造,含丰富的花瓣海百合科化石(图2),顶部清晰的泥裂构造显示桐梓上升期的海退标志(邓小杰等,2012;戎嘉余等,2012)。

图2 上扬子区陆表海3 个埃隆阶剖面的岩性序列以及赋存花瓣海百合化石的生物多样性和丰度Fig.2 Lithological loggings of the three Aeronian sections with diversity and abundance profiles of the Petalocrinidae fossils in the Upper Yangtze epicontinental sea
中国科学院南京地质古生物研究所(1974)在石阡县城北命名志留系香树园组、雷家屯组和马脚冲组序列并分层描述岩性和化石,雷家屯组与下伏香树园组整合接触,顶部可见桐梓上升期的海退泥裂(戎嘉余等,2012)。雷家屯组分布从黔中古陆黔东北的近岸向湘西远岸带延伸,下部以黄绿色页岩间夹薄层或透镜状灰岩为主,上部多灰岩或泥灰岩;腕足类、三叶虫、珊瑚、海百合等壳相化石丰富(葛治洲等,1979)。不同于石阡香树园雷家屯组建组剖面以及石阡白沙剖面的雷家屯组出现丰富的灰岩层,石阡枫香铺子沟剖面的雷家屯组主要是碎屑岩,花瓣海百合化石赋存于该组上部的灰黄色、黄色泥岩、粉砂岩中(图2)。
谢家荣等(1925)在鄂西宜昌分乡大中坝剖面命名志留系罗惹坪层;中国科学院南京地质古生物研究所(1974)将龙马溪群之上含壳相化石丰富的灰岩称罗惹坪组;葛治洲等(1979)、Mu et al(.1986)、汪啸风等(1987)以及戎嘉余等(1990)列述罗惹坪组化石的多样性。罗惹坪组主要分布于鄂西一带,岩性为粉砂岩、泥岩夹少量壳相生物滩灰岩。荆门仙居剖面的罗惹坪组为薄层生屑灰岩夹少量粉砂质泥岩,花瓣海百合化石集中于该组中部的薄层生屑灰岩中(图2)。
2 上扬子区埃隆阶花瓣海百合特化方式和类群划分
花瓣海百合科特殊性在于其冠部的腕呈五幅对称排列,每个腕仅有1 块很小的一级腕板,和一块高度特化的二级骨板,完整保存的二级腕板标本展现清晰的腕部食物沟骨板,高度特化的二级腕部是由多个腕肢骨板胶结而成,二级腕板形态是花瓣海百合科最大的鉴定特征。Ausich et al.(2019)对花瓣海百合科特化发育的腕扇切片研究,确定膨大的单个二级腕扇是由多个腕胶结而成。上扬子陆表海区花瓣海百合化石分布广且研究程度高,在露头上易于识别。含泥灰岩中保存的棘皮动物器官化石在风化松软岩层中易于剥离采集;陆源碎屑粉砂和泥质含量特别低的生物礁或生屑滩相灰岩质地坚硬,尽管其中的棘皮动物骨片含量非常丰富,有时能达到密集排列形成颗粒支撑,但因棘屑具有单晶片式的特殊矿物学结构,在成岩过程中骨片常出现次生加大重结晶现象,化石与碳酸钙亮晶或基质完全固结,在野外露头上不可能对此花瓣海百合化石碎片进行完全剥离的种级鉴定。本文依据在野外露头上能表现花瓣海百合特化成种方式的原则,次分A~F 这6 个大类(图3),其形态学识别标准介绍如下。
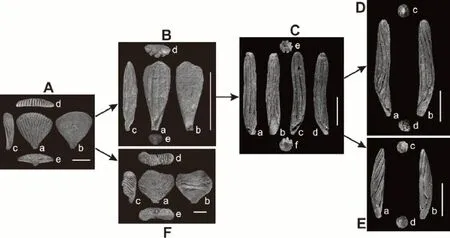
图3 上扬子陆表海埃隆期花瓣海百合科动物群腕部形态学演化单元Fig.3 Morphological evolutionary taxa of the Aeronian(Silurian)Petalocrinidae fauna in the Upper Yangtze epicontinental sea
A 类:这是花瓣海百合科分布最广泛且种数最多的属,包括目前鉴定为Petalocrinus属中排除Petalocrinus stenopetalus的所有种。半圆形的关节面,近三角形胶结的腕和伸展的远端,有多达32 条远端食物沟,相邻的腕并没有旁侧接触,食物沟仅仅在口面;腕部食物沟呈扇形排列。
B 类:Petalocrinus stenopetalus,腕扇形态学特征介于Petalocrinus和Spirocrinus之间,其生长的初期阶段与Petalocrinus相似,腕在长度和宽度上同时扩展并进行分支,约3 次分支后的腕就不再侧向生长,而仅限于两侧平行延展成长条状,与Spirocrinus的形态相似;其中部的食物沟最多只分支一次,侧面两边的食物沟可多达3 级分支,食物沟直,成年期腕扇食物沟总数为6~10 条,与Spirocrinus的食物沟数目基本一致。
C 类:Spirocrinus circularis(直立螺旋海百合),马蹄形关节面,胶结的腕肢圆柱形,两端锥形,食物沟7~11 条,相邻的腕在旁侧不接触,腕扇两侧向背面包卷直至腕扇两侧胶结愈合,背侧中部的背中脊在始端较宽。食物沟多为直的,或有较小的旋转度,左侧视稍向右旋,右侧视稍向左旋。
D 类:所有左旋的Spirocrinus,马蹄形的关节面,胶结的腕肢为圆柱形,两端为锥形,食物沟多达10~12 条,相邻的腕在旁侧不接触,食物沟围绕胶结的腕向左盘旋。
E 类:Spirocrinus dextrosus(右旋螺旋海百合),关节面马蹄形,胶结的腕肢为圆柱形,两端为锥形,食物沟7~10 条,相邻的腕在旁侧不接触,食物沟围绕胶结的腕向右盘旋一周。
F 类:即Sinopetalocrinus,马蹄形的关节面;胶结的腕在宽度上快速地扩展一小段距离,然后两侧平行,最后形成一个凸的远端边缘;多达40 条远端食物沟,相邻的腕在旁侧并不接触,在口面和反口面都有食物沟。
Mao et al.(2017)对这6 大类的形态学演化关系进行了详细的分析讨论,即Petalocrinus独立地向Sinopetalocrinus和Spirocrinus演化。Petalocrinus通过将腕扇末端向反口面包卷胶结从而形成Sinopetalocrinus。而Petalocrinus演变成Spirocrinus则经历了多次形态变化,食物沟先减少变窄加长,演化成Petalocrinus stenopetalus;Petalocrinus stenopetalus食物沟向背侧包卷愈合后,演化成Spirocrinus circularis;Spirocrinus circularis向左或右旋转,演化成所有左旋的Spirocrinus和Spirocrinus dextrosus(右旋),即食物沟从扇形—直立—包卷—螺旋形。
3 上扬子区花瓣海百合动物群地层记录实例
本文选择上扬子陆表海区的3 个埃隆阶同期异相剖面分析其中的花瓣海百合科生物多样性和丰度明显差异(图4),桐梓水坝塘剖面代表黔中古陆之北石牛栏组的生物礁沉积区,石阡枫香剖面代表黔中古陆之东的浅海粉砂质泥岩沉积区,以及荆门仙居剖面代表的外陆棚的生屑滩灰岩沉积区。

图4 上扬子区埃隆阶花瓣海百合科化石的赋存特征Fig.4 Photographs showing profiles of the Aeronian Petalocrinidae fossils in the Upper Yangtze region
3.1 桐梓水坝塘石牛栏组生物礁相的花瓣海百合
桐梓水坝塘镇北侧的羊石山和南侧的虎头城剖面(N 28°44'44",E 107°3'55")均出露石牛栏组石牛栏段灰岩,多为40 余米厚灰岩为中型点礁建造(图2)。松坎段上部瘤状灰岩中的泥质粉砂质成分逐渐减少,从石牛栏段底界向上数米为高清澈浅海造礁环境,可识别出上下两期造礁。第一期点礁生长之前潮道沉积之上的生屑颗粒滩作为礁基,中型规模点礁10余米宽、数米厚,礁核间距百余米;而第二期大型点礁礁核厚度多可超过20 m,直径百余米;两期礁核灰岩均是大量原地生长或基本原地保存的珊瑚和少量层孔海绵格架组成,造礁动物大格架岩之间其它不同粒度的腕足类、苔藓虫以及软体动物生屑丰富,礁核应生长于水动力条件强且富氧的海底高地位置,造礁群落与全球同期的生物礁比较具有很大的相似度,可鉴定床板珊瑚Favosites、Mesofavosites、Paleofavosites、Troedssonites、Quasifletcheriella、Halysites、Cladopora、Coenites和Heliolites; 四射珊瑚Enterophyllum、Parastauria、Aphyllum、Microplasma、Stauria、Ceriaster、Tabularia、Paraceriaster、Mackenziephyllum、Dentilasma、Tryplasma、Ceriaster、Cystiphyllum、Crassilasma和Rhizophyllum;层孔海绵Clathrodictyon、Labechia和Plestylostroma;钙藻Solenopora、Girvanella和Hedstroemia;腹足类Hormotoma;三叶虫Coronocephalus、Latiproetus和Encrinoides(Wang et al.,2014)。
礁核中的造礁生物主要为大个体的Favosites和Halysites形成大型原地格架,枝状和丛状复体四射珊瑚、单体四射珊瑚如Cystophyllum、层孔海绵和苔藓虫形成少量的障积岩或盖覆岩,障积岩或层孔海绵盖覆岩之间花瓣海百合密集,局部可达腕扇颗粒支撑(图4a、图4b)。2 期珊瑚—层孔海绵礁顶部生屑滩灰岩中花瓣海百合碎片可作为核型石核心颗粒(图4c),或密集堆积并达到生屑颗粒支撑程度的棘屑滩,以颗粒粒度分选性好的Sinopetalocrinus最丰富,局部层中达到腕部碎片密度支撑排列(图4d)。该属腕扇偏厚的形态学特征是对高能量水流环境的适应,Sinopetalocrinus伴生大量强水流冲蚀破碎散落后的粗颗粒海百合茎以及珊瑚、苔藓虫细砾共同沉积。花瓣海百合科的6 个形态大类均可在此礁相单元中见及,在野外露头上大致估计其丰度差异显示于图2,花瓣海百合科化石丰富,总采获的标本逾200 枚,其中Petalocrinus占比50%,Spirocrinus占比40%,以左旋的Spirocrinus居多,完整保存的Sinopetalocrinus较少。Mao et al.(2017)报道的Petalocrinus的 1 个新种Petalocrinus stenopetalus和Spirocrinus的两新种Petalocrinus stenopetalus和Spirocrinus dextrosus的标本就是在第二期礁的礁翼相薄层含泥生屑灰岩夹少量泥岩和粉砂岩中获得的,目前未发现于石牛栏组非点礁生态区剖面中,3 个新种栖居的生态空间局限于礁翼相水动力偏低的环境,与腕足类Eospirifer,Nalivkinia以及少量小型珊瑚Favosites、Microplasma伴生,周缘礁核珊瑚—层孔海绵格架岩具有抗浪性,能有效缓冲水流能量,使较细的生屑颗粒和少量泥质粉砂质颗粒在礁翼相沉积,该低能环境中也常见各类花瓣海百合腕部化石。石牛栏段顶部出现核形石滩—泥裂序列(图4e),指示造礁结束的根本原因是桐梓上升的海底暴露。
3.2 黔东北石阡枫香雷家屯组粉砂质泥岩相的花瓣海百合
枫香剖面香树园组沉积时海水深度偏大并制约石阡县城北白沙型生物礁滩的延展区间分布(于深洋等,2017)。与石阡县城北雷家屯组建组标准剖面比较,枫香剖面的少量灰岩层单层和总体厚度明显要小得多,灰岩层灰泥丰富且生屑含量低(毛颖颜等,2015),呈薄层或透镜体夹含于黄色黄绿色粉砂质泥页岩中(图2)。雷家屯组沉积序列中总体化石丰富度很低,无笔石,壳相化石仅见零散的珊瑚、腕足类、三叶虫和海百合茎碎片,其化石密度达不到生屑滩的指标。花瓣海百合腕部仅零星出现于粉砂质泥岩中,每平方米约 2~3 个腕扇(图 4f),分属 3 个大类(图 2),Petalocrinus、Spirocrinus和Sinopetalocrinus属均有产出,但以A 类为主,占比约50%,此外D 和F 也有少量产出。进入雷家屯组沉积期海底深度仍然偏大,无最大浪基面以上或风暴浪基面以上的沉积标志,故而推论这里属于偏深安静海底的泥质粉砂质环境,不利于大量壳相动物栖居。
3.3 鄂西荆门仙居剖面罗惹坪组生屑滩相的花瓣海百合
2013 年笔者曾在宜昌大中坝剖面罗惹坪组含泥薄层壳相生屑灰岩中仔细采集,但仅获3 枚Petalocrinus碎片。荆门仙居剖面出露罗惹坪组地层,下部为灰色薄层灰岩和黄色钙质泥岩,产珊瑚、腕足类、苔藓虫和花瓣海百合科化石;中部为灰色生物碎屑灰岩和灰绿色钙质粉砂质泥岩(图2),产珊瑚、腕足类、遗迹化石、三叶虫和花瓣海百合科化石;上部为黄色粉砂岩、泥岩夹薄层生屑灰岩,粗颗粒的珊瑚苔藓虫以及腕足动物等生屑颗粒丰富,分选性中等,是清澈动荡浅海底的生屑滩。花瓣海百合科化石丰度中等,包含Petalocrinus和Sinopetalocrinus二属,以A 类为主,占比约75%,另外有少量Sinopetalocrinus(图2),与海百合茎以及少量珊瑚、苔藓虫等生屑共同堆积形成生屑滩(图4g),或Petalocrinus与Sinopetalocrinus以及珊瑚等共同堆积形成棘屑滩(图4h)。
4 讨 论
海百合是研究海底表栖群落古生态学的重要指示物(Brett,1978;Brett and Eckert,1982;McGee and Watkins,1994),其底栖生态单元多样性构建方式与海底环境参数的动态变化密切相关。海百合化石栖居方式受海底环境指标控制,古地理和古生态分区明显(Franzen,1974,1977;Guensburg and Sprinkle,1992;Peters and Bork,1998),陆表浅海区多元化生态系统分异由不同比例的广适性和狭适性物种组合而成(Watkins and McGee,1998)。志留纪不同岩相中展示的化石组合型式取决于海底深度和底质类型(Ziegler,1965;Watkins,1981)。Bret(t1991)强调志留纪海百合茎是浅海沉积物中最丰富的生屑颗粒类型之一。
花瓣海百合科特征在于其独有的由5 个大腕扇或者腕束组成特化愈合腕板,形态—功能分析的重要内容是推测花瓣海百合科特化腕扇的摄食方式,目前涉及花瓣海百合形态功能学的文献多涉及腕扇的特化方式讨论。Bathe(r1893)强调花瓣海百合最重要的形态学特征表现为小球形的萼部反口面着生5 个扇形腕,具有独立扇形分支是花瓣海百合科特化的食物沟,该科所有的属种都有强大的食物沟系统;Ausich et al(.2019)认为Petalocrinus通过多个腕胶结而成的腕扇可明显增强滤食效率,该科各属种多形态的腕板表明其有多样性的滤食方式。在形态学优势上,Sinopetalocrinus和Spirocrinus的腕具有更多选择来渠导水流方向;从滤食腕和捕食功能优势看,Sinopetalocrinus和Spirocrinus可选择更多的获食方式。上扬子陆表海区埃隆阶上述3 个区域地层单元中,花瓣海百合科化石赋存面貌,尤其是土著类型分布范围和产出丰度指标,均表现出地层学记录的非均一性。罗惹坪组的花瓣海百合科化石与其它生屑颗粒混合沉积形成滩相灰岩,雷家屯组粉砂质泥岩中丰度最低,单调的花瓣海百合属种与同样低丰度的其它大化石伴生。
黔北桐梓水坝塘距离遵义董公寺附近黔中古陆海岸线近100 km,陆源碎屑影响力度小,温暖高清澈度的浅海环境有利于石牛栏段后生动物格架礁以及附礁栖居的壳相动物群集中生长,这里花瓣海百合科化石密度和多样性最高,多个花瓣海百合类型与其它壳相动物化石密集形成棘屑滩或礁灰岩亚相。较之全球分布且在上扬子区远岸泥岩、近岸礁滩相灰岩和泥岩中最常见的Petalocrinus,上扬子区特有的土著属Spirocrinus和Sinopetalocrinus的部分种地理分布特别狭窄,在此生物礁分布区丰度也最高,说明石牛栏组生物礁灰岩是花瓣海百合动物群最佳生态位,Sinopetalocrinus和Spirocrinus胶结的腕扇重厚,且骨骼密度远大于海水,推测花瓣海百合科动物群更趋于选择腕扇贴近于海底的栖居方式,这不仅有利于提高在海底固着栖居的稳定性,也是对礁滩相频繁出现高能水动力条件的形态学适应。Petalocrinus却成为分布区域最广泛,衍生出种级分类单元最多样性的属,Mao et al(.2017)建立Spirocrinus的2 个新种仅见于黔北桐梓水坝塘石牛栏组近顶部的礁翼亚相,生物礁是其成种生态域,在造礁区成种后具有极端生态狭适性,很难拓展其空间分布,未发生广泛迁移。Mao et al(.2017)复原的志留纪花瓣海百合科形态学演化过程完全在上扬子区陆表海浅水区埃隆中晚期短暂时限内完成的,从Petalocrinus分化出其它3 个属之后,祖先属和后裔属在上扬子区并存,具有成种时间短、分化快、地域局限等特征,诠释了点断平衡的成种模式。埃隆末期桐梓上升导致的海域缩减(戎嘉余等,2012),水坝塘生物礁区的海底暴露导致花瓣海百合动物群迅速消失;特列奇早期大量陆源泥质和粉砂导致高浑浊度的海底(汪啸风等,1987;陈旭等,1996),使花瓣海百合动物群迁出了华南板块。
5 结 论
上扬子陆表海区埃隆阶花瓣海百合科的短暂生存和非均一性分布均受控于古地理模式;灰岩和陆源碎屑岩沉积分异和古地理变迁是重要因素。花瓣海百合化石的多样性和丰度与其它底栖壳相动物化石的同类指标呈正相关。浅水清澈度高的海底沉积生物礁或生屑滩灰岩更有利于花瓣海百合科的底栖生长,浑浊度高的粉砂质泥岩区花瓣海百合稀疏。广适性类型从黔中古陆近岸带延伸到鄂西远岸带,广适性和狭适性类型共同生长于生物礁相带。上扬子陆表海广泛分布的粉砂质泥岩沉积区制约了土著类型在生物礁区成种后的向外迁移扩增,埃隆末期的古地理事件,即扬子上升海退事件,以及特列奇阶全扬子区的陆源碎屑岩广布加速了花瓣海百合生存空间的丧失。
致 谢笔者特别感谢审稿人提出的修改意见。
