运动神经网络CPG的研究进展
2022-10-28张鹏
张 鹏
前言
动物能正常并准确无误地协调机体各个组织进行诸如行走、呼吸、吞咽等基础生命活动与CPG神经网络有着非常紧密的联系。CPG的“半中心”网络结构使其能保证运动信号及其相位的稳定,也能使动物在不同的运动模式之间进行平稳地切换。CPG在动物的生长发育及生存中发挥着举足轻重的作用,同时在当下人工智能与运动控制等前沿研究中扮演着奠基石的角色,因此对这个领域的研究进行总结与普及尤为重要。
1911年Brown在实验中发现剔除大脑的猫仍然能产生基本的踏步动作[1]。这说明在没有大脑的控制下,简单的重复运动仍然能够由脊髓产生,既脊髓中存在某个网络能控制或维持节律运动。1967年Jankowska在猫的脊髓实验首次为CPG的存在提供了的证据[2]。时至今日,人类对CPG已经有了非常深入的了解,研究范围也从运动扩展到游泳、飞行、呼吸等多个领域。本文主要总结了CPG在运动方面的作用与贡献,同时也归纳出一些这个领域仍不清楚的问题为后续研究提供参考。
1 CPG与运动的简介
CPG是可以在没有信号输入的情况下产生节律输出的生物神经环路[3-5]。它由一对或多对神经元振荡器与反馈回路组成地复杂的分布式神经网络。CPG的输出并不固定[4, 5],这是因为CPG神经元膜特性和突触连接网络会根据输入或环境条件的改变灵活地变化,比如呼吸节律会根据运动强度改变、鸟的飞行模式会因条件变化。一个网络被称为CPG要满足两个条件:1、有两个或多个相互作用的过程,此消彼长;2、在相互作用的影响下,网络能反复回到初始状态[4]。CPG特殊的结构使得动物能产生稳定的节律性运动,如步行、游泳、呼吸或咀嚼。
运动通常分为反射运动、随意运动和节律运动三大类。反射运动重复简单不需要大脑控制,如膝跳反射;随意运动灵活多变且需要大脑控制,如打篮球、踢足球;节律运动介于这两类运动之间,既具备一定的灵活性又具备一定的机械性,如边走路边玩手机。节律运动不需要大脑的持续控制且启动后能自发重复进行,这是节律运动的关键特征。一般来说,我们把能产生节律运动的神经网络环路称为CPG。现在,CPG已在无脊椎动物[6]和几乎所有研究的脊椎动物物种中发现[7, 8],包括人类[9, 10]。
2 CPG的生理学特性
2.1 CPG神经元的特性
CPG的节律产生取决于神经元的内在膜特性及其突触连接。和其他神经网络一样,CPG的基本组成单元是神经元。构成CPG的神经元具有不同的内在膜特性[11]。一类神经元始终能产生动作电位的节律性放电,无论是在内源性还是加入神经调节物质条件下(图1)。另一类神经元具有双稳态且能产生平台电位,这种电位可以由去极化电流脉冲触发,由超极化电流脉冲中止(图1b)。前者主要构成起搏器-跟随器连接,后者主要构成相互抑制连接,关于连接方式本文会在下一小节详细介绍。此外,许多CPG神经元在抑制中释放后会放电(图1c),在恒定去极化(尖峰频率适应)期间放电频率会降低(图1d)。这些膜特性对CPG节律活动中不同状态的转换有着极大的贡献。
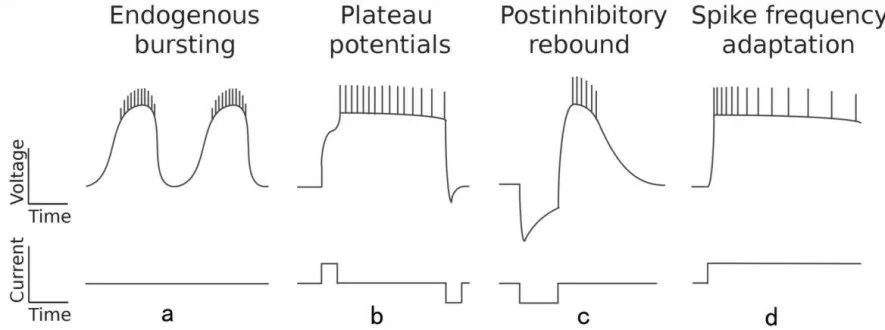
图1 CPG神经元的内在特性[11]
2.2 CPG神经元的连接机制
节律的产生一般有两种连接机制:起搏器-跟随器和相互抑制[11]。在起搏器驱动的连接中(图2),由一个或多个神经元充当核心振荡器,驱动其他神经元产生节律信号。这种网络存在于甲壳类动物胃神经元的幽门节律[12]和脊椎动物的呼吸节律[13]。在相互抑制驱动的连接中(图2b),两组神经元相互抑制,无论何时只有一半处于激活状态所以称为半中心振荡网络。单个神经元无法产生节律信号,但通过抑制性连接耦合时,它们可以产生交替重复的节律信号。相互抑制连接是许多CPG的核心特征,比如与运动相关的CPG[14-17]。

图2 CPG节律产生的连接机制[11]
CPG的网络具有广泛的反复突触连接,包括相互激活或抑制两类。这些连接对节律活动的激活、终止起着重要的作用[18, 19]。此外,结合前文所述,激活神经元的尖峰频率适应可能会使其他神经元从抑制中释放出来从而达到状态转换的目的[20]。突触连接方式会受习惯性活动修改[21],比如不同的跑步姿势的CPG突触连接不同。刻意练习某一动作会建立相应的CPG网络,这也是运动训练会强化相应身体机能的重要因素。
2.3 CPG神经元的位置
不同功能的CPG位置有所不同,参与运动控制的CPG神经元位于脊椎动物的下胸椎和腰椎区域[22]以及无脊椎动物腹神经索区域[23];参与吞咽的CPG神经元位于脑干中,尤其是髓质内的舌下核[24, 25]。虽然CPG的大概位置能被推断出来,但由于CPG神经元最近才被发现且CPG又具备灵活重组的特性,所以CPG准确的位置很难被锁定。
3 CPG的网络结构模型
由于生理实验很难既能深入解剖又能长时间保持实验样本活性,加上运动神经网络庞大且连接众多因素,节律运动准确的CPG结构仍不清楚,因此很多相关的研究是以理论模型的方式展开。行进运动的CPG包含CPG绝大多数的特征,为此以行进运动为切入点了解CPG是很多科研人员的不二之选。行进运动是节律运动中比较复杂的运动,它的特点是简单重复并且产生位移,像人的行走、鸟的飞翔,马的奔跑等。
3.1 单侧肢体的CPG结构模型
从1914年布朗提出半中心模型至今[14],CPG已经有非常完善的发展。最初的单层半中心模型无法描述真实的运动过程。动物的运动过程会根据环境或条件发生变化,这就需要有反馈回路。再者,运动的节律产生(RhythmGenerator,RG)和模式发生(PatternFormation,PF)也是会发生改变的,所以CPG的结构应该有调节运动节律和模式的部分。最后,产生的节律信号会通过运动神经元(Motoneurons)作用于骨骼肌并使其交替地屈伸从而产生形变[26]。图3是综合以上条件得到的单侧肢体的3层CPG网络结构模型,其包含了反馈回路、运动节律和模式调节结构及运动神经元。

图3 三层CPG结构[26]
3.2 CPG之间的协调
单侧肢体的屈伸变化还不足以产生运动。一个简单的运动过程需要多个CPG的参与和协调。Shevtsova在2015年的模型研究中通过联合中间神经元(Commissural interneurons,CINs)构建左右肢体CPG的网络结构[27]。随后Danner在前者基础之上用模型仿真出脊椎动物四肢交替的CPG结构[28]。每个肢体都由一个CPG控制;前后肢则通过一组抑制性神经元构成一个更大的CPG网络;左右侧肢通过CNS控制;整个网络都受脊髓调控(图4)。这种结构不仅能够很好地模拟老鼠的行走、小跑、疾驰、跳越等节律信号,也能很好地拟合之前生理实验的结果。

图4 四肢交替的CPG结构[28]
4 总结与展望
本文通过简短的篇幅总结了CPG的特征,神经元膜特性和连接方式,运动CPG网络模型的发展。这些研究均表明,CPG在节律运动中发挥着无可替代的作用。这些研究为揭示运动的本质做出了建设性贡献,同时也为人工智能时代构建高智能化的机器人提供了理论支撑。然而,目前这个领域仍有许多问题尚待解决,诸如控制节律运动产生的准确CPG结构仍然不确定、生理实验技术的局限性、模型仿真研究很难与生理研究有效结合等。解决这些问题,还需要有足够的生理与模型实验的数据作为支撑;需要相关技术的进步与提高;需要研究人员在这个领域持续不断的耕耘。
