水肥耦合效应对核桃属2种幼苗抗寒性的影响
2022-10-28杨梦思罗莎莎王如月叶春秀虎海防
杨梦思, 罗莎莎, 王如月, 叶春秀, 虎海防
(1.新疆农业大学林学与风景园林学院,新疆乌鲁木齐 830052; 2.新疆林业科学院,新疆乌鲁木齐 830063;3.新疆佳木果树学国家长期科研基地,新疆阿克苏 843000)
核桃属植物适应性强,用途多,果实含油率高,为重要的木本油料、干果树种和用材树种。核桃(L.)为核桃属核桃种,是世界上最重要的坚果树种之一,具有极高的经济价值。核桃栽培历史悠久,种植规模巨大,是我国重要的经济林树种。其产地大多分布于干旱半干旱地区,春夏季节干旱少雨,且降水不均匀,水分成为影响核桃苗木生长发育的关键因子。因此在干旱半干旱地区研究灌水对核桃生长发育的影响,对促进水分的高效利用有着重要意义。目前核桃的施肥情况仍是凭经验施肥,盲目施肥、随意施肥的现象比较普遍,制约了核桃产业的健康发展。黑核桃(L.)为核桃属黑核桃组Rhycocaryon,原产于北美洲,具有生长快、抗风沙、耐低温、耐干旱、耐盐碱等特性,是经济价值较高的林果兼用树种。由于黑核桃耐干旱、根系发达、生长快,且与核桃有较强的亲和力,近年来被广泛用作薄皮核桃的优良砧木,以提高核桃的产量和品质。在我国多处已引种栽培黑核桃,产生了显著的经济、生态及社会效益。新疆从1991年起引种栽培黑核桃,但在实际生产中,施肥和灌水依然存在一定的盲目性。在农业生产中,水分和养分在植物生长发育过程中起着重要的相互作用。由于水分和肥料之间存在一定的协同效应和激励机制,即耦合效应,因此在不同水肥条件下,植物生长会略有差异。在实际生产中,要充分利用水肥的耦合效应,建立合理模式。核桃属于喜温树种,抗寒性较差,容易遭受低温和晚霜的危害,这已经成为核桃生产中的重要限制因子。我国核桃砧木培育研究起步较晚,与其他果树砧木相比,在培育、评价和应用等方面还有待加强与完善。新疆丰富的核桃种质资源在育种工作中得到较广泛的应用,利用新疆早实核桃结实早、增产潜力大等优良性状,我国以此作为改良核桃育种的主要亲本取得了丰硕的成果。目前新疆核桃使用的砧木主要是晚实核桃,砧木育种滞后,没有实现砧木良种化。与栽培品种选育工作相比,砧木育种评价难度大、周期长,缺乏广适性强、抗性突出、综合性状优良的砧木品种。
近年来,水肥耦合及抗寒性在核桃上的应用主要集中在核桃树上,而幼苗阶段是核桃树生长的必经阶段。本试验以核桃属2种幼苗为试材,采用田间水肥试验分析核桃幼苗的抗寒指标,进一步选出各品种核桃幼苗最优的水肥组合及抗寒性好的种类,以期为新疆核桃砧木良种化提供技术支撑,并为干旱半干旱地区核桃幼苗水肥耦合效应研究提供理论基础。
1 材料与方法
1.1 试验地概况
试验地为新疆佳木果树学国家长期科研基地。位于新疆阿克苏地区温宿县(80°32′E,41°15′N)境内,海拔1 103.8 m,属暖温带大陆性干旱气候,四季分明,昼夜温差大,多晴少雨,光照充足,空气干燥。年平均气温10.10 ℃,年平均降水量65.4 mm,年平均无霜期185 d。土壤为沙壤土,土层深厚,腐殖质和有机质含量较高,含有机质10.787 g/kg、碱解氮81.329 mg/kg、速效磷18.780 mg/kg、速效钾 83.088 mg/kg,pH值为6.547。
1.2 试验材料
试验材料采用黑核桃品种东部黑核桃和普通核桃品种扎343。于2020年9月10日前从样品树上采集大小均匀饱满的种子各3 000粒。
1.3 试验方法
1.3.1 试验设计 采用大田育苗方式。选择背风向阳、地势平坦、排灌方便等条件良好的育苗地,对苗床进行土壤及水分处理。于2020年11月上旬播种,每100 m播种大约1 000粒种子,株行距为 20 cm×50 cm,各样地之间间隔3 m,方便控水,各样地内播种相同面积的2种核桃。对施肥和灌水进行控制,肥料分别使用尿素(N≥46%)、磷酸二氢钾(PO≥52%,KO≥34%)。施肥设置3个梯度(15、10、5 kg/667 m),2021年5月底、6月底各追施尿素1次,7月底、8月底各施磷钾肥1次;灌水设置3个梯度(10、15、30 d/次),灌水处理从5月底苗木出苗整齐后开始进行,即在6、7、8月进行。9月初停水肥,防止幼苗徒长。水肥因子共9个耦合处理,分别为:1-1,高水高肥,灌水10 d/次,施肥 15 kg/667 m;1-2,高水中肥,灌水10 d/次,施肥10 kg/667 m;1-3,高水低肥,灌水10 d/次,施肥 5 kg/667 m;2-1,中水高肥,灌水15 d/次,施肥 15 kg/667 m;2-2,中水中肥,灌水15 d/次,施肥 10 kg/667 m;2-3,中水低肥,灌水15 d/次,施肥 5 kg/667 m;3-1,低水高肥,灌水30 d/次,施肥 15 kg/667 m;3-2,低水中肥,灌水30 d/次,施肥 10 kg/667 m;3-3,低水低肥,灌水30 d/次,施肥 5 kg/667 m。采取随机区组设计,每个处理重复3次。
1.3.2 指标测定
1.3.2.1 木质化程度指标的测定 于2021年11月下旬在各处理相对一致的部位上采集一年生枝条,采用紫外分光光度法进行木质素及纤维素含量的测定,使用试剂盒(上海酶联生物科技有限公司)根据测定的吸光度计算含量,各指标处理重复3次。
1.3.2.2 生理指标的测定 于2021年11月下旬在各处理相对一致的部位上采集一年生枝条,将枝条放入已贴好标签的自封袋中,带回实验室用自来水清洗干净后,用蒸馏水冲洗3次,用滤纸将枝条上的水分吸干。取自封袋将其分为4 组,置于冰箱中进行模拟低温处理,设置温度为-10、-20、-30 ℃,以 4 ℃ 作为对照,在冰箱中处理12 h。每次取出经各温度处理后的材料,马上测定其相对电导率。将剩余的材料放于冻存管中并标注记号,在液氮中迅速冷冻之后置于-80 ℃的超低温冰箱中保存以测定其余生理生化指标。每个指标处理重复3次。测定的苗木生理指标包括可溶性糖(SS)含量、丙二醛(MDA)含量、脯氨酸(Pro)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性。
相对电导率采用电导率法测定。处理后的枝条去除分节部位,剪去节间部位,将样品剪成3~5 mm的薄片,混合均匀后称取2 g放入锥形瓶,分别置于20 mL去离子水的试管中,盖上试管塞置于室温下浸泡12 h,用雷磁DDS-307A型电导仪测定浸提液电导率(),然后沸水浴加热30 min,冷却至室温,再次测定浸提液的电导率(),计算其相对电导率(REC)。相对电导率=×100%。然后拟合Logistic方程,计算各材料的半致死温度(LT)。
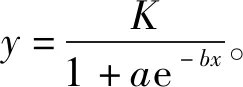
式中:为电导率;为曲线渐近线,即为最大电解质渗出率;、均为方程参数;求曲线的拐点(ln),即为低温半致死温度(LT)。
可溶性糖(SS)含量采用蒽酮比色法测定;丙二醛(MDA)含量采用硫代巴比妥酸法测定;游离脯氨酸(Pro)含量采用酸性茚三酮法测定;过氧化物酶(POD)活性采用愈创木酚法测定;超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法测定。
1.4 数据处理
利用Micrisoft Excel 2010软件进行数据整理及图表制作;指标先通过 Excel进行分析后,利用SPSS 24.0软件进行方差分析、相关性分析、主成分分析,再采用隶属函数值法对其抗寒能力进行综合排序,最后通过聚类分析对抗寒能力进行分类。其中隶属函数法求平均隶属函数值分2种情况:
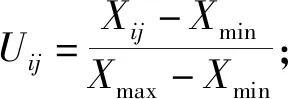
2 结果与分析
2.1 不同水肥处理对核桃幼苗抗寒性的影响
2.1.1 不同水肥处理对核桃幼苗木质化程度指标的影响 如表1所示,水分、施肥及水肥交互作用对2种核桃幼苗木质素、纤维素含量均能产生极显著影响(<0.01)。不同水肥处理下对2种核桃幼苗木质化程度的影响见表2。其中东部黑核桃幼苗木质素含量2-1处理最高,为92.543 ng/L,3-2处理最低,为74.873 ng/L,纤维素含量2-1处理最高,为519.600 μg/mL,3-3 处理最低,为 417.299 μg/mL;扎343幼苗木质素含量2-1处理最高,为98.118 ng/L,3-3处理最低,为 72.243 ng/L,纤维素含量2-1处理最高,为520.776 μg/mL,3-3处理最低,为395.545 μg/mL。
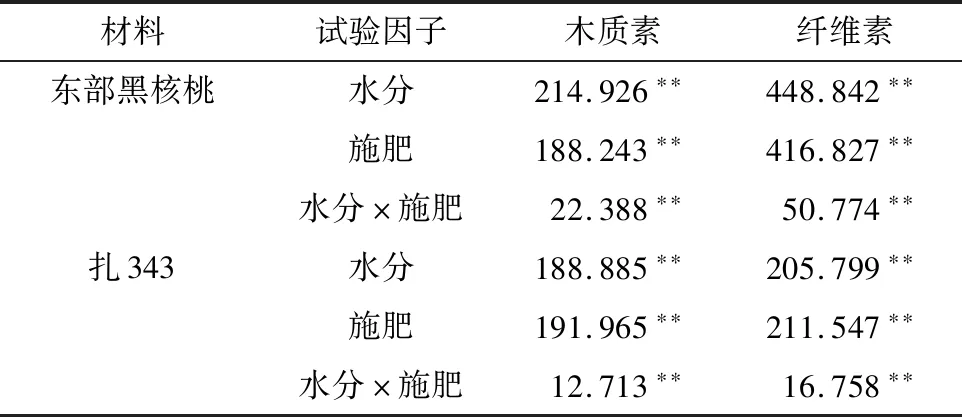
表1 不同水肥处理对核桃幼苗木质素及纤维素含量

表2 不同水肥处理下核桃幼苗木质素及纤维素含量
2.1.2 低温胁迫对不同水肥处理下核桃幼苗相对电导率及半致死温度的影响 如表3所示,水分、施肥及水肥交互作用对2种核桃幼苗相对电导率均能产生极显著影响(<0.01)。由图1可以看出,在低温胁迫下,供试枝条的电解质渗出率随着温度的下降而增加,说明随着温度的降低,细胞膜损害增加,膜透性增大,枝条对低温胁迫的适应能力降缓。其中在-10 ℃~-20 ℃之间电导率上升最快,-20 ℃~-30 ℃之间电导率增长逐渐减小,其变化曲线基本符合“S”型曲线变化趋势。在-30 ℃处,东部黑核桃幼苗(图1-a)所有处理中,1-1处理的相对电导率最小,为50.47%;1-3处理下最大,为60.84%。扎343幼苗(图1-b)所有处理中, 2-1 处理的相对电导率最小,为50.58%;3-2处理最大,为64.22%。
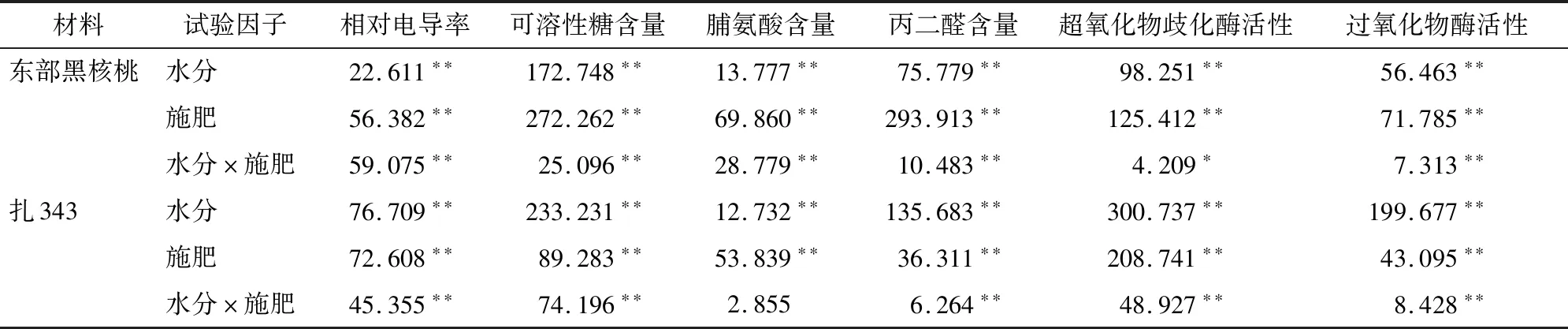
表3 不同水肥处理对核桃幼苗抗寒指标的方差分析(F值)

LT的大小可用作判断植物抗冻能力的依据。利用低温胁迫对不同水肥处理下核桃幼苗相对电导率的变化与Logisitic方程相结合,可以计算出不同种类核桃幼苗在不同水肥处理下的半致死温度(表4)。各方程拟合后的回归系数均在0.9以上,拟合度较好。其中东部黑核桃幼苗半致死温度从大到小为3-3>1-3>3-2>3-1>2-3>1-2>2-2>1-1>2-1;3-3处理下半致死温度为 -18.639 1 ℃,抗寒性最弱;2-1处理下半致死温度为-27.181 4 ℃,抗寒性最强。扎343幼苗半致死温度从大到小为3-2>3-3>2-3>1-3>2-2>3-1>1-2>1-1>2-1;3-2处理下半致死温度为-15.352 4 ℃,抗寒性最弱;2-1处理下半致死温度为-27.770 8 ℃,抗寒性最强。
2.1.3 低温胁迫对不同水肥处理下核桃幼苗可溶性糖含量的影响 当树木遭受低温伤害时,通常通过可溶性糖的积累来维护体内细胞的渗透势从而适应低温环境,提高抗寒性。不同水肥处理对核桃幼苗相对电导率的方差分析如表3所示。水分、施肥及水肥交互作用对2种核桃幼苗可溶性糖含量均产生了极显著影响(<0.01)。由图2可知,随着温度的下降,各处理下2种核桃幼苗的可溶性糖含量基本呈先下降后上升再下降的趋势,并均在 -20 ℃ 出现最高值,说明在-20 ℃受到的伤害程度最大。到达峰值时,东部黑核桃幼苗(图2-a)SS含量从高到低为2-1>1-1>2-3>1-2>3-1>2-2>3-3>1-3>3-2;3-2处理的SS含量为108.085 mg/g,抗寒性最弱; 2-1处理的SS含量为168.204 mg/g,抗寒性最强。扎343幼苗(图2-b)SS含量从高到低为2-1>1-2>2-2>1-1>2-3>1-3>3-1>3-3>3-2;3-2处理的SS含量为101.937 mg/g,抗寒性最弱;2-1处理的SS含量为152.848 mg/g,抗寒性最强。
2.1.4 低温胁迫对不同水肥处理下核桃幼苗脯氨酸含量的影响 游离脯氨酸作为渗透调节物质,参与细胞水分调节,诱导植物抗逆性,在低温胁迫下能够主动积累,提高植物的抗寒能力。不同水肥处理对核桃幼苗脯氨酸含量的方差分析如表3所示。除了水肥交互作用对扎343幼苗脯氨酸含量未产生显著影响(>0.05)之外,水分、施肥及水肥交互作用对2种核桃幼苗脯氨酸含量均产生极显著影响(<0.01)。由图3可知,随着温度的下降,2种核桃幼苗在各处理下脯氨酸含量均呈先上升后下降的趋势,并均在-20 ℃出现最高值,说明在 -20 ℃ 受到的伤害程度最大。到达峰值时,东部黑核桃幼苗(图3-a)Pro含量从高到低为2-1>1-2>3-1>1-1>2-3>2-2>3-3>1-3>3-2;3-2处理下Pro含量为0.262 μg/g,抗寒性最弱;2-1处理下Pro含量为0.394 μg/g,抗寒性最强。扎343幼苗(图3-b)Pro含量从高到低为2-1>1-1>3-1>1-2>1-3>2-2>2-3>3-2>3-3;3-3处理下Pro含量为0.182 μg/g,抗寒性最弱,2-1处理下Pro含量为0.330 μg/g,抗寒性最强。

表4 低温胁迫下不同水肥处理核桃幼苗的半致死温度

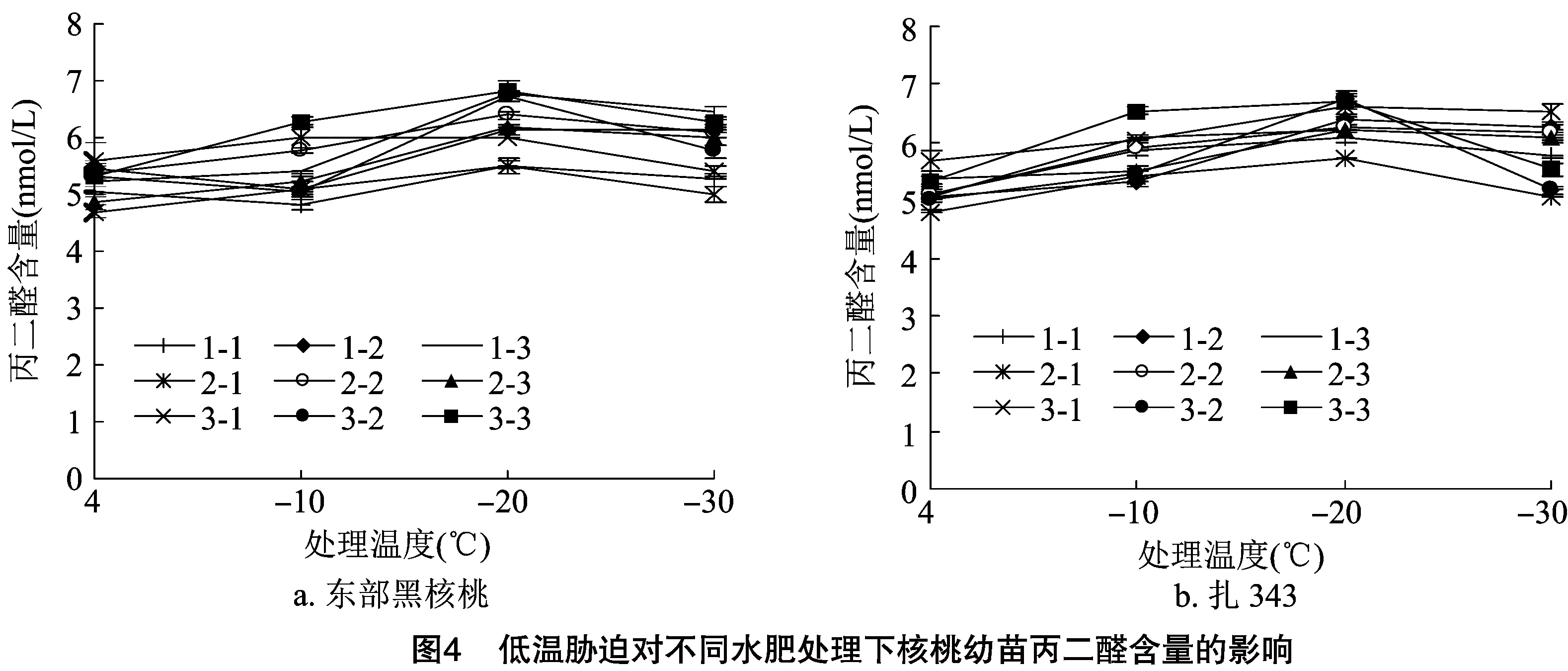
2.1.5 低温胁迫对不同水肥处理下核桃幼苗丙二醛(MDA)含量的影响 植物在遭受低温伤害时会发生膜脂过氧化,在这个过程会产生丙二醛,对细胞质膜有毒害作用,丙二醛的含量能够体现出植物受到不良环境伤害时的严重程度。不同水肥处理对核桃幼苗丙二醛含量的方差分析如表3所示。水分、施肥及水肥交互作用对2种核桃幼苗丙二醛含量均能产生极显著影响(<0.01)。如图4所示,随着温度的下降,2种核桃幼苗在各处理下的丙二醛含量均呈先上升后下降的趋势,前期上升趋势十分缓慢,说明受到的伤害胁迫较小;随着温度的降低,MDA含量逐渐升高,到达-20 ℃时出现最高值,说明在-20 ℃受到的伤害程度最大。到达峰值时,东部黑核桃幼苗(图4-a)MDA含量从高到低为3-3>1-3>3-2>2-2>2-3>1-2>3-1>2-1>1-1;3-3处理下MDA含量为6.812 nmol/L,抗寒性最弱;1-1处理下MDA含量为5.501 nmol/L,抗寒性最强。扎343幼苗(图4-b)MDA含量从高到低为3-2>3-3>3-1>1-2>2-2>1-3>2-3>2-1;3-2处理下MDA含量为 6.770 nmol/L,抗寒性最弱;2-1处理下MDA含量为5.728 nmol/L,抗寒性最强。

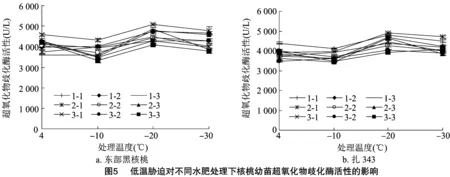
2.1.6 低温胁迫对不同水肥处理下核桃幼苗超氧化物歧化酶活性的影响 在低温胁迫下,植物会产生大量的活性氧,会破坏植物细胞膜,对植物造成伤害。此时,植株会通过增强超氧化物歧化酶的活性来清除活性氧的伤害。不同水肥处理对核桃幼苗SOD活性的方差分析如表3所示。除了水肥交互作用对东部黑核桃幼苗SOD活性产生显著影响(<0.05)之外,水分、施肥及水肥交互作用对2种核桃幼苗SOD活性均产生极显著影响(<0.01)。如图5所示,随着温度的下降,2种核桃幼苗在各处理条件下SOD活性均呈先下降后上升再下降的趋势,并均在-20 ℃出现最高值,说明在 -20 ℃ 受到的伤害程度最大。当温度持续下降时,细胞受到的胁迫超出自身耐受范围,SOD活性下降。到达峰值时,东部黑核桃幼苗(图5-a)SOD活性从高到低为2-1>1-1>2-3>1-2>3-1>2-2>3-2>1-3>3-3;3-3处理下SOD活性为4 087.125 U/L,抗寒性最弱;2-1处理下SOD活性为5 087.125 U/L,抗寒性最强。扎343幼苗(图5-b)SOD活性从高到低为2-1>1-1>1-2>2-3>3-1>1-3>2-2>3-2>3-3;3-3处理下SOD活性为3 920.758 U/L,抗寒性最弱;2-1 处理下SOD活性为4901.906 U/L,抗寒性最强。
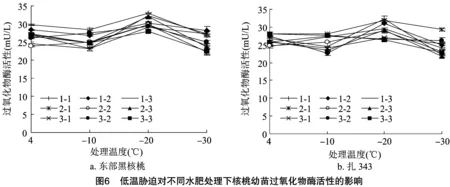
2.1.7 低温胁迫对不同水肥处理下核桃幼苗过氧化物酶活性的影响 过氧化物酶是植物体内普遍存在的一种保护酶,在低温引起膜脂过氧化时能通过酶促系统的响应来增强植物的抗寒能力。不同水肥处理对核桃幼苗POD活性的方差分析如表3所示。水分、施肥及水肥交互作用对2种核桃幼苗POD活性均能产生极显著影响(<0.01)。如图6所示,随着温度的下降,2种核桃幼苗在各处理下POD活性基本呈先下降后上升再下降的趋势,并均在-20 ℃出现最高值,说明在 -20 ℃ 受到的伤害程度最大,当温度持续下降时,细胞受到的胁迫超出自身耐受范围,POD活性进一步下降。到达峰值时,东部黑核桃幼苗(图6-a)POD活性从高到低为1-1>2-1>2-3>1-2>2-2>3-1>3-2>1-3>3-3;3-3处理下POD活性为27.923 mU/L,抗寒性最弱;1-1处理下POD活性为32.950 mU/L,抗寒性最强。扎343幼苗(图6-b)POD活性从高到低为1-1>2-1>1-2>1-3>2-3>2-2>3-1>3-2>3-3;3-3处理下POD活性为 26.354 mU/L,抗寒性最弱;1-1处理下POD活性为32.239 mU/L,抗寒性最强。
2.1.8 各抗寒指标间的相关性分析 将测定的9个抗寒指标进行相关性分析(表5)。半致死温度与丙二醛含量存在显著正相关,相关系数为0.335;与可溶性糖、脯氨酸、木质素、纤维素含量存在极显著负相关关系,相关系数分别为-0.756、-0.692、-0.872、-0.910;与超氧化物歧化酶活性存在显著负相关,相关系数为-0.343。以半致死温度作为抗寒评价标准,可以反映出各指标与抗寒性之间的关系,即可溶性糖含量、脯氨酸含量、超氧化物歧化酶活性、过氧化物酶活性、木质素含量、纤维素含量与抗寒性呈正相关,指标越高,抗寒性越强;丙二醛含量、相对电导率、半致死温度与抗寒性呈负相关,指标越高,抗寒性越弱。
2.1.9 不同水肥处理下核桃幼苗抗寒能力综合评价分析 抗寒性是多项指标综合作用的结果,用单一指标评价植物的抗寒性不具有代表性。本研究为了较全面地反映核桃幼苗的抗寒性,分别运用主成分分析、隶属函数值分析及聚类分析进行综合分析。
2.1.9.1 指标主成分分析 利用SPSS 24.0进行主成分分析。由表6可知,东部黑核桃幼苗第1主成分的特征根大于1,且对综合抗寒性的贡献率达到87.988%,可以反映大部分的信息,所以提取第1主成分对东部黑核桃幼苗抗寒性作综合评价;扎343幼苗第1主成分的特征根大于1,且对综合抗寒性的贡献率达到84.147%,可以反映大部分的信息,所以提取第1主成分对扎343幼苗抗寒性作综合评价。抗寒指标的权重通过第1主成分中各指标负荷量与贡献值计算各抗寒性指标对抗寒性的作用大小,确定权重。由表6可知,对东部黑核桃幼苗抗寒性影响由强到弱的指标依次是SOD活性、半致死温度、纤维素含量、可溶性糖含量、丙二醛含量、木质素含量、相对电导率、POD活性、脯氨酸含量;对扎343幼苗抗寒性影响由强到弱的指标依次是相对电导率、半致死温度、纤维素含量、木质素含量、SOD活性、POD活性、丙二醛含量、可溶性糖含量、脯氨酸含量。
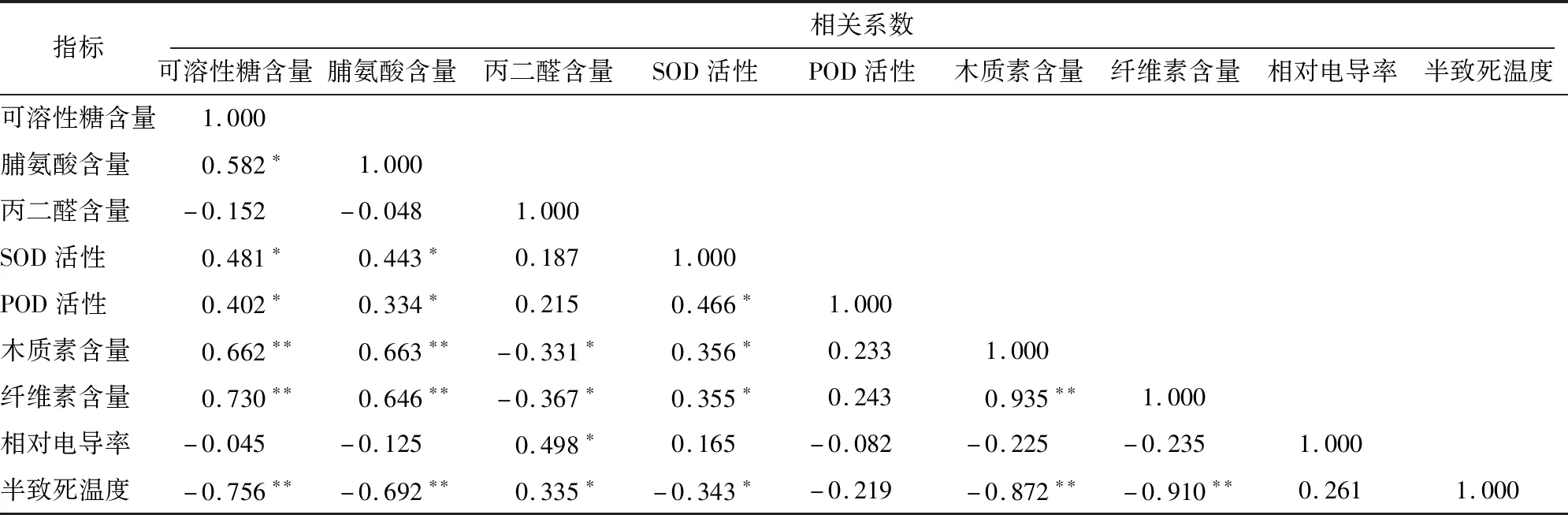
表5 核桃幼苗抗寒指标的相关性分析

表6 不同水肥处理下核桃幼苗抗寒指标的特征根、贡献率及负荷量、权重
通过综合得分模型计算2种不同处理核桃幼苗的综合得分,并进行排序得到表7。东部黑核桃幼苗不同处理抗寒能力从强到弱排序为2-1>1-1>1-2>2-3>3-1>2-2>1-3>3-2>3-3;扎343幼苗不同处理抗寒能力从强到弱排序为2-1>1-1>1-2>2-2>2-3>3-1>1-3>3-2>3-3。
2.1.9.2 指标隶属函数值分析 运用隶属函数法,用隶属度对不同水肥处理的2种核桃幼苗的抗寒能力进行综合评价。通过对所测指标进行相关性分析可知,可溶性糖含量、脯氨酸含量、SOD活性、POD活性、木质素含量、纤维素含量采用升型函数公式,丙二醛含量、相对电导率、半致死温度采用降型函数公式。根据各项指标隶属度值与权重,通过加成法则=×,为各指标权重,为各指标隶属度值,对各项指标进行合成,得到抗寒性综合指数。由表8可知,根据抗寒性综合指数的大小,东部黑核桃幼苗不同处理抗寒能力从强到弱排序为2-1>1-1>2-2>1-2>3-1>2-3>1-3>3-2>3-3;扎343幼苗不同处理抗寒能力从强到弱排序为2-1>1-1>1-2>2-2>2-3>3-1>1-3>3-2>3-3。

表7 不同水肥处理下核桃幼苗抗寒指标主成分综合排序
综合表6、表7,主成分分析法所得结论与隶属函数值法所得结论基本一致,说明用这2种方法中的任何一种,都可以作为核桃幼苗抗寒能力的评价方法。
2.1.9.3 聚类分析 利用SPSS 24.0进行聚类系统分析。东部黑核桃幼苗(图7-a)9种处理根据其抗寒能力的强弱及抗寒指标所占比重,在遗传距离为10时可以分为3组:第1组为2-1、1-1,抗寒能力最强; 第2组为1-2、2-2、3-1、2-3,抗寒能力中等;第3组为3-3、3-2、1-3,抗寒能力最弱。
扎343幼苗(图7-b)9种处理根据其抗寒能力的强弱及抗寒指标所占比重,在遗传距离为10时可以分为3组:第1组为2-1、1-1,抗寒能力最强;第2组为1-2、2-3、3-1、2-2、1-3,抗寒能力中等;第3组为3-3、3-2,抗寒能力最弱。
2.2 2种核桃幼苗抗寒性的差异
2.2.1 2种核桃幼苗木质化程度的差异 由表9可知,2种核桃幼苗木质素和纤维素含量存在显著差异(<0.05)。东部黑核桃、扎343幼苗的木质素含量分别为84.90、82.96 ng/L,东部黑核桃>扎343;东部黑核桃、扎343幼苗纤维素含量分别为462.635、450.615 μg/mL,东部黑核桃>扎343。
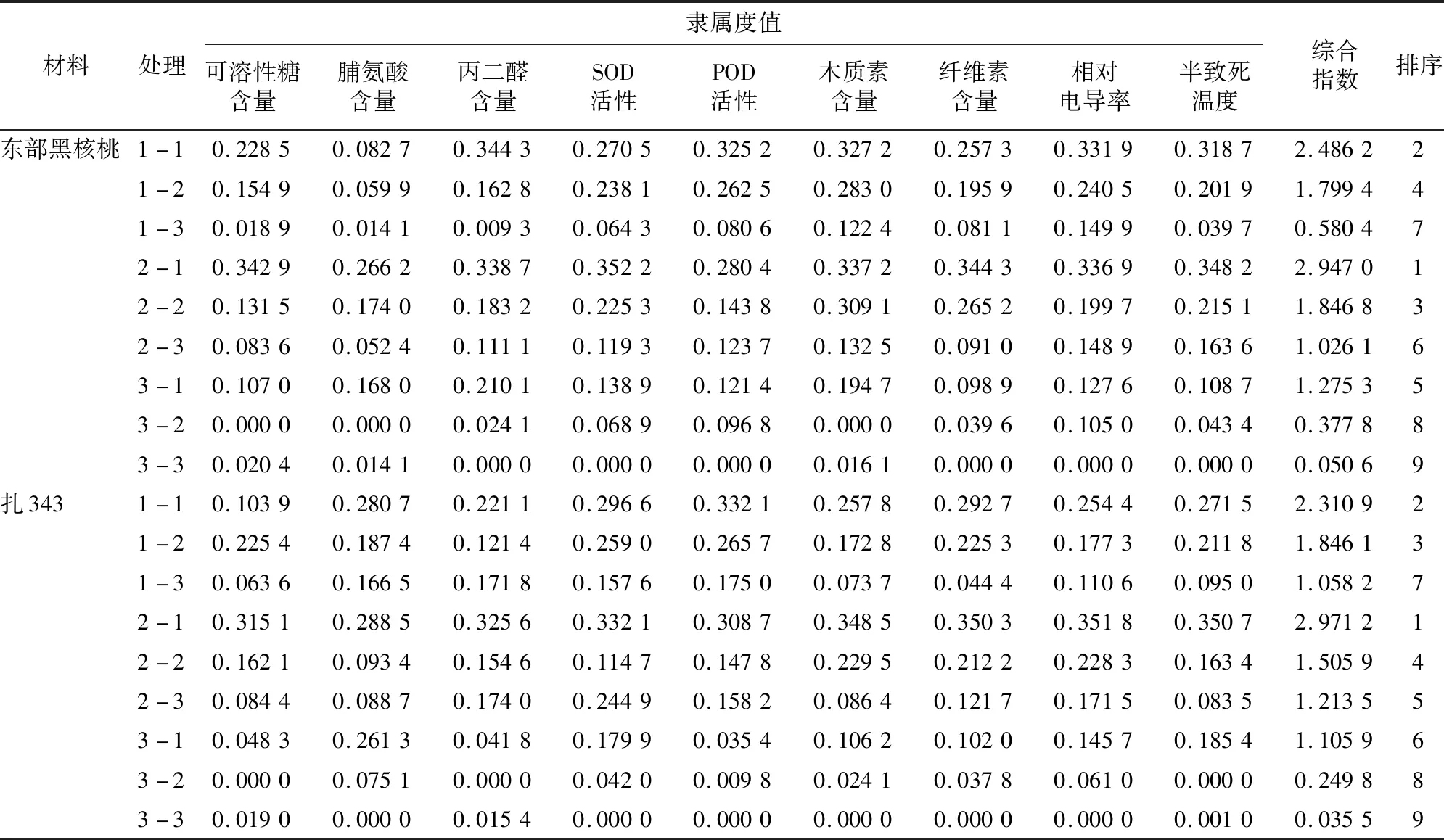
表8 不同水肥处理下核桃幼苗抗寒指标隶属函数值综合排序

表9 2种核桃幼苗木质素及纤维素含量
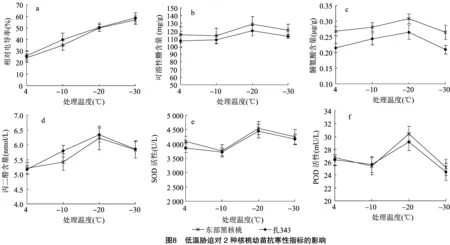
2.2.2 低温胁迫对2种核桃幼苗相对电导率及半致死温度的影响 由图8-a可以看出,在低温胁迫下,供试枝条的电解质渗出率随着温度的下降而增加,说明随着温度的降低,细胞膜损害增加,膜透性增大,枝条对低温胁迫的适应能力降低。在-30 ℃处,东部黑核桃幼苗相对电导率最大,为56.74%;扎343幼苗相对电导率最大,为58.63%。LT的大小可用作判断植物抗冻能力的依据。利用低温胁迫对2种核桃幼苗相对电导率的变化与Logisitic方程相结合,可以计算出2种核桃幼苗的半致死温度(表10)。各方程拟合后的回归系数均在0.9以上,拟合度较好。扎343的半致死温度高于东部黑核桃,扎343幼苗半致死温度为 -20.526 4 ℃,抗寒性较弱;东部黑核桃幼苗半致死温度为-22.632 2 ℃,抗寒性较强。

表10 低温胁迫下2种核桃幼苗的半致死温度
2.2.3 低温胁迫对2种核桃幼苗可溶性糖含量的影响 由图8-b可以看出,随着温度的下降,2种核桃幼苗可溶性糖含量呈先下降后上升再下降的趋势,并均在-20 ℃出现最高值,说明在-20 ℃受到的伤害程度最大。到达峰值时,SS含量为东部黑核桃>扎343,扎343幼苗SS含量为120.283 mg/g,抗寒性较弱;东部黑核桃幼苗SS含量为129.271 mg/g,抗寒性较强。
2.2.4 低温胁迫对2种核桃幼苗脯氨酸含量的影响 由图8-c可以看出,随着温度的下降,2种核桃幼苗的脯氨酸含量呈先上升后下降的趋势,在 -20 ℃ 出现最高值,说明在-20 ℃受到的伤害程度最大。到达峰值时,Pro含量为东部黑核桃>扎343,扎343幼苗 Pro含量为0.264 μg/g,抗寒性较弱;东部黑核桃幼苗Pro含量为0.308 μg/g,抗寒性较强。
2.2.5 低温胁迫对2种核桃幼苗丙二醛含量的影响 由图8-d可以看出,随着温度的下降,2种核桃幼苗的丙二醛含量呈先上升后下降的趋势,并在-20 ℃出现最高值,说明在-20 ℃受到的伤害程度最大。到达峰值时,MDA含量为扎343>东部黑核桃,扎343幼苗MDA含量为6.334 nmol/L,抗寒性较弱;东部黑核桃幼苗MDA含量为 6.227 nmol/L,抗寒性较强。
2.2.6 低温胁迫对2种核桃幼苗超氧化物歧化酶活性的影响 由图8-e可以看出,随着温度的下降,2种核桃幼苗的SOD活性呈先下降后上升再下降的趋势,并均在-20 ℃出现最高值,说明在 -20 ℃ 受到的伤害程度最大。到达峰值时,SOD活性为东部黑核桃>扎343,扎343幼苗SOD活性为4 454.766 U/L,抗寒性较弱;东部黑核桃幼苗SOD活性为4 553.271 U/L,抗寒性较强。
2.2.7 低温胁迫对2种核桃幼苗过氧化物酶活性的影响 由图8-f可以看出,随着温度的下降,2种核桃幼苗POD活性呈先下降后上升再下降的趋势,并均在-20 ℃出现最高值,说明在 -20 ℃ 受到的伤害程度最大。到达峰值时,POD活性为东部黑核桃>扎343,扎343幼苗 POD活性为29.175 mU/L,抗寒性较弱;东部黑核桃幼苗POD活性为 30.387 mU/L,抗寒性较强。
2.2.8 2种核桃幼苗抗寒能力综合评价分析
2.2.8.1 指标主成分分析 利用SPSS 24.0进行主成分分析。由表11所示,第1主成分与第2主成分的特征根均大于1,且对综合抗寒性的累计贡献率达到95.141%,可以反映大部分的信息,所以提取第1主成分及第2主成分对2种核桃幼苗抗寒性作综合评价。抗寒指标的权重通过第1主成分及第2主成分中各指标负荷量与贡献值计算各抗寒性指标对抗寒性的作用大小,确定权重。由表11可知,对核桃幼苗抗寒性影响由强到弱的指标依次是脯氨酸含量、丙二醛含量、木质素含量、可溶性糖含量、POD活性、SOD活性、纤维素含量、半致死温度、相对电导率。通过综合得分模型计算2种核桃幼苗综合得分,并进行排序得到表12。2种核桃幼苗抗寒能力为东部黑核桃>扎343。
2.2.8.2 指标隶属函数值分析 运用隶属函数法,综合各项相关指标用隶属度对2种核桃幼苗的抗寒能力进行综合评价(表13)。根据抗寒性综合指数的大小,2种核桃幼苗抗寒能力为东部黑核桃>扎343。
3 讨论
在低温胁迫下, 植物中的酚类化合物的合成速度加快,随后这些酚类物质就会进入细胞壁间,最终形成木质素。国外研究者认为,木质化与软木质沉积物可以作为细胞壁之间的防水屏障,可以提高植物的抗寒能力。在逆境中,植物可以通过调控体内一系列的生理生化过程来增强抵御逆境的能力。苗阳通过对野杏抗寒能力的研究,发现LT与大多数生理指标之间都存在着显著正或负相关,因此,LT可以作为鉴定植物抗寒能力简单快捷的单因素指标。利用相对电导率结合Logistic曲线方程可推导半致死温度,植物半致死温度作为评价植物抗寒性强弱的指标, 能准确反映植物所耐受的低温极限。本研究采用Logistic方程对低温处理后2种核桃幼苗不同水肥处理核桃枝条相对电导率与温度关系曲线进行拟合,所有处理的拟合度值均达显著水平,所求半致死温度可靠。2种核桃幼苗在9个水肥处理中,LT由低到高顺序有所差别,但最低均为2-1处理,说明在2-1处理下抗寒能力最强;2种核桃幼苗枝条的LT中,东部黑核桃半致死温度低于扎343,黑核桃抗寒能力要强于普通核桃,与前人研究结果一致。

表11 2种核桃幼苗抗寒指标的特征根、贡献率及负荷量、权重

表12 2种核桃幼苗抗寒指标主成分综合排序

表13 2种核桃幼苗抗寒指标隶属函数值综合排序
本研究中,LT与木质化程度指标及生理指标均存在显著相关性,可以用来评价抗寒能力。相关性分析表明,LT与MDA含量呈显著正相关;与SS、Pro、木质素、纤维素含量存在极显著负相关关系;与SOD活性存在显著负相关关系;POD活性和相对电导率虽然没有达到显著水平,但也有一定的规律性。这些抗寒指标与LT之间没达到显著水平,其原因可能是由于测定的指标本身就涉及到复杂的生理过程,受到多种因素的影响。最后各指标与抗寒性的关系为可溶性糖含量、脯氨酸含量、SOD活性、POD活性、木质素含量、纤维素含量与抗寒性呈正相关,指标越高,抗寒性越强;丙二醛含量、相对电导率、半致死温度与抗寒性呈负相关,指标越高,抗寒性越弱。
植物抗寒性是一个复杂的生理生化变化过程,用单一指标很难反映其抗寒本质。为克服单个抗寒指标的不足,隶属函数法、主成分分析法及聚类分析法等已经广泛应用于植物抗寒性的综合评价。本研究通过比较主成分分析法与隶属函数法发现,两者所得结论基本一致,说明这2种方法都可以作为核桃幼苗抗寒能力的评价方法。9种处理下2种核桃幼苗排序略有区别,但均为2-1处理排序第一;2种核桃幼苗抗寒能力为东部黑核桃>扎343。利用聚类分析法对不同水肥处理抗寒能力的强弱进行归类分析,最终可将2-1、1-1分为一组,抗寒能力最强,2-1 处理(中水高肥)为最强抗寒能力水肥组合。抗寒能力表现为黑核桃>普通核桃。
4 结论
本研究通过比较核桃属2种幼苗在不同水肥处理下的抗寒性,采用主成分分析、隶属函数值分析、聚类分析等方法对不同水肥处理下及不同种类核桃幼苗进行综合评价。研究表明,不同的水肥处理显著影响核桃幼苗的抗寒性,合理的水肥管理有利于促进核桃幼苗抗寒能力的提高。中水高肥下核桃幼苗抗寒能力最强,黑核桃抗寒能力强于普通核桃。期望本研究结果可以为新疆核桃砧木良种化提供技术支撑,并为干旱半干旱地区核桃幼苗水肥耦合效应研究提供理论基础,同时为鉴定、筛选和培育优良核桃种质资源提供科学依据。本研究仅对核桃砧木实生苗进行了评价,下一步工作应考虑实际生产中砧木与接穗间的互作效应,进而为核桃砧穗的正确选择、抗逆栽培及优良抗寒砧木品种的选育提供参考。
