十字花科栽培作物根肿病抗性研究进展
2022-10-27苏贺楠杨双娟魏小春赵艳艳王志勇尹喜恩张晓伟原玉香
苏贺楠,杨双娟,魏小春,赵艳艳,王志勇,尹喜恩,张晓伟,原玉香
(1.河南省农业科学院园艺研究所 郑州 450002; 2.鄢陵县甘罗养生养老有限公司 河南 鄢陵 461200)
十字花科根肿病是由芸薹根肿菌(Woron.)侵染引起的世界范围内普遍发生的一种重要土传病害,在全世界多个国家均有分布。黄齐望等首次报道了十字花科作物根肿病在我国江西发生,近年来蔓延速度迅猛、发生程度加重、分布范围极广、经济损失严重。据统计,十字花科作物根肿病目前在我国20 多个省(自治区、直辖市)均有不同程度地发生,尤其在西南部和中部发病地区危害最严重。该病专性寄生在十字花科作物的根部,包括白菜、菜心、油菜、甘蓝、花椰菜、青花菜、芥蓝、萝卜、芥菜和芜菁等栽培作物,通过侵染根毛导致根部薄壁细胞增生而形成肿瘤,对大面积十字花科栽培作物生产影响巨大,一旦发病,蔬菜作物产量急剧下降,经济效益严重下滑。
1 根肿病病害特点、症状及分类
1.1 根肿菌的生物学特征
根肿病病原菌为芸薹根肿菌,同时具有真菌和原生动物特性,目前普遍认为该菌属于原生动物界、根肿菌门、根肿菌纲、根肿菌目、根肿菌属。根肿菌在土壤中的主要存活状态是形态和大小不一的休眠孢子。王靖等总结出油菜根肿菌休眠孢子直径为1.7~4.3µm,白菜根肿菌休眠孢子直径为2.2~4.2µm,结球甘蓝根肿菌休眠孢子直径为1.6~4.0µm,根肿菌休眠孢子呈近球形。
当遇到适宜的环境条件时,土壤中的休眠孢子萌发释放初级游动孢子,初级游动孢子形成变形体,靠近并接触寄主根毛表皮,产生一种管状结构侵入细胞壁,完成根毛侵染阶段,此阶段的感病植株没有明显的表型病征。初级游动孢子在侵染的根毛内形成原生质团,原生质团经过大量的核分裂后形成多个游动的孢子囊,已经成熟的游动孢子囊会释放出次级游动孢子,随后2 个次级游动孢子融合形成双核的次级游动孢子,融合后的次级游动孢子侵入植株根部皮层细胞,完成皮层侵染阶段。皮层侵染阶段会导致植物根部细胞异常增生,引发根部肿大甚至畸形,形成不同形状的肿瘤。
1.2 根肿菌的发病症状
根肿菌专性寄生在十字花科作物根部,从寄主植株的幼苗期到成株期均可发病。在发病初期,受侵染的植株发病症状并不明显,只在侧根处出现一些小瘤子,肿根表面通常比较光滑,对植物的地上部分生长基本没有影响。但是伴随着根肿菌侵染时间的延长,植株主根也出现肿瘤,根部的肿瘤逐渐膨大,肿根表面变得粗糙且发生龟裂,使其他的杂菌容易入侵根部,造成植株吸收养分受阻,从而导致发病植株地上部长势较弱、叶片逐渐萎蔫,且越到侵染后期发病越严重,最终引起整个植株枯萎死亡。
根肿菌侵染不同的十字花科栽培作物,根部肿瘤的存在部位、大小以及形状也会产生差异(图1)。当根肿菌侵染的寄主是白菜、油菜、甘蓝和芥菜类蔬菜作物时,主根的肿瘤体积大且数量少,而在侧根的肿瘤体积小且数量多,根瘤的形态特征表现为球形、手指状或者短棒状;当萝卜、芜菁等蔬菜作物受到根肿菌侵染时,主根一般无肿瘤或者肿瘤存在于主根的末端,根上有较大缢缩黑疤,须根上有较多的肿瘤,侵染后期根上黑疤凹陷形成大的黑洞,且中空腐烂。

图1 大白菜、甘蓝和萝卜根肿病田间发病情况统计标准[12]
1.3 根肿病生理小种鉴定
Honig于1931 年首先提出根肿菌生理小种存在变异分化,随着研究的深入,Ayers、Lamrnerink、Williams、Johnston、Toxopeus和Buczack等多位学者已经建立了多种鉴定根肿菌小种分化的系统,目前国内外学者公认的根肿菌生理小种的鉴别系统为Williams 建立的Williams 系统和Buczacki 等建立的European Clubroot Differential(ECD)系统。Williams 系统的鉴别寄主分别由2个芜菁甘蓝品种和2 个结球甘蓝品种共4 个十字花科栽培作物品种组成,Williams 通过统计这4 个栽培作物品种对根肿菌不同的抗性反应,从而制定了Williams 系统,该体系可以把从16 个不同的国家和地区收集而来的124 个菌株划分为9 个不同的根肿菌生理小种。去除Williams 鉴别系统中1个芜菁甘蓝品种,增加12 个不同的十字花科鉴别寄主品种,利用这些鉴别寄主对不同生理小种的根肿病抗性来鉴定生理小种,命名为欧洲鉴别寄主体系的鉴别系统(ECD)。
根肿菌活体寄生的特殊性导致自然环境中并不存在单孢生理小种,侵染时都是以混合生理小种的状态存在,因此通过鉴别系统鉴别出的小种仅为该菌株的优势生理小种。目前我国大部分地区的主要优势小种为4 号生理小种,还有一些其他生理小种也被相继报道。原玉香等通过Williams 鉴别系统明确了河南省新野大白菜根肿菌是4 号生理小种,通过ECD 系统明确其生理小种为ECD16/15/31 和ECD21/31/31,说明经Williams 鉴别系统鉴定的该地区4 号生理小种实际存在致病性差异;黄蓉等通过Williams 鉴别系统对江西省油菜根肿病发病区域进行鉴定,发现致病菌为4 号和9 号生理小种;张晶等利用单孢分离技术和Williams生理小种鉴定方法对中国湖北利川、内蒙古通辽、陕西太白、山东青岛、北京、河南南阳、辽宁大连、辽宁沈阳以及云南昆明等9 个大白菜主产区的根肿菌进行分离及鉴定,发现4 号生理小种为中国大白菜主产区的根肿菌优势小种,同时还包含除5 号以外的所有生理小种。
2 根肿病抗病基因与分子标记的研究进展
迄今,伴随着科技的不断发展,根肿病的相关研究已经从遗传学的理论水平发展到分子生物学水平,越来越多的十字花科栽培作物的抗根肿病基因被定位出来,通过杂交等常规育种途径和克隆、转基因等分子生物学手段导入感病的优良品种中,加快了十字花科作物抗根肿病的育种步伐。因此,研究根肿病的抗病机制和抗病性的遗传规律,不仅可以为十字花科栽培作物抗病育种提供相应的理论依据,另外,对于挖掘抗病基因、加快抗病育种的进程以及改良育种技术都具有十分重要的意义。
2.1 白菜抗根肿病基因定位与克隆研究
大白菜(ssp.,AA=20)属于芸薹属芸薹种,是起源于我国的一种重要蔬菜。其根肿病抗性研究工作进行较早,白菜中的根肿病抗性基因来源于不同亚种的欧洲饲用芜菁(ssp.)。在1998 年,Matsumoto等利用RFLP 标记HC352b 和HC181 将根肿病抗性基因定位于A03 连锁群上,遗传距离分别为3 cM 和12 cM。2003 年,Suwabe 等利用4号生理小种接种抗性来源于Siloga 芜菁的抗病大白菜F分离群体,定位到了2 个与根肿病抗性相关的位点,分别命名为(位于A08)和(位于A01),并筛选出2 个紧密连锁的SSR 标记BRMS-088 和BRMS-096。2004 年,Hirai 等利用Ano-01 生理小种对大白菜抗病分离群体接种,经遗传分析确定一个由1 对主效基因控制的新的抗根肿病基因(位于A03)。2006 年,Suwabe 等利用抗感根肿病大白菜材料G004 和A9709 构建的F群体,绘制遗传连锁图谱并进行共线性分析,发现了位于A06 染色体上的。Piao 等对抗性来源于ECD01 芜菁的大白菜分离群体接种4 号生理小种,发现具有抗性的位于A03 连锁群,显性SCAR 标记TCR09 与基因紧密连锁,遗传距离为0.78 cM,2 个侧翼标记TCR01 和TCR09 与之间的遗传距离为2.9 cM。Zhang 等认为和是两个紧密相连的抗性位点。Sakamoto等通过接种2 号生理小种和K04 菌株,定位了来源于Debra 芜菁的抗病基因,分别命名为(位于A03)和( 位 于 A02)。 Laila 等利 用ddRAD-seq 技术鉴定到1 个新的抗性位点(位于A08),不同于以往报道的位于A08 染色体上的。Chen 等定位了来源于ECD04 芜菁的抗根肿病基因.、.和.,这3 个位点分别抗2、10 和7 号生理小种。Kato 等对抗病性来源于Gelria 芜菁的抗病大白菜材料进行定位研究,定位到抗病基因CRb。Pang 等对大白菜抗感根肿病的F分离群体进行定位研究,发现了位于A03 染色体60 kb 区域内的,且在的上游区域。Chu 等通过构建连锁遗传图谱及转录组学测序手段将()精细定位在A03 连锁群上,两侧分子标记MS7-9 和sN8591,遗传距离分别为0.06 cM 和0.19 cM。Huang 等通过对大白菜抗感根肿病材料进行RNA-seq 研究,结果发现了位于A03 染色体近端区的。 Huang 等利 用Bulked Segregant RNA Sequencing(BSR-Seq)技 术 在A03 染 色 体23 339 019~23 465 030 bp 之间定位到1 个新的抗病基因,命名为。Chang 等对大白菜抗感根肿病池进行BSR-Seq 和KASP 分析,结果定位到了1 个新的抗性基因,位于A08 染色体14.8~15.4 Mb 之间,命名为。Yu 等在大白菜根肿病抗性研究中定位了3 个抗性基因(位于A03)、(位于A02)和(位 于A08)。Karim 等利 用BSR-Seq 技术在大白菜A08 染色体上定位了2 个根肿病抗性位点和Rcr9。Choi 等对大白菜抗根肿病研究发现1 个位于和位点附近区域的抗病位点PbBrA08,通过标记验证、基因的结构变异分析等手段确定了该位点是不同于上述抗病基因的新位点。Wei 等对大白菜抗根肿病研究发现1 个位于A03 染色体80 kb 区域内的,通过RACE 克隆技术克隆出的全长序列,包含3 个外显子和2 个内含子,与已报道的(3 个外显子和3 个内含子)CDS 序列一致。
目前芸薹属A 基因组已定位的抗根肿病基因或位点有20 余个(表1),、、CRb和基因已经被克隆,Hatakeyama 等通过构建区间的高密度分子遗传图谱和BAC 文库,以紧密连锁的分子标记为探针对BAC 文库进行筛选,随后利用PCR 技术进行cDNA 末端快速克隆,最后分别在抗病和感病材料中克隆出全长序列。Ueno 等利用拟南芥及大白菜参考基因组序列信息,通过cDNA 末端快速克隆技术最终克隆出,另外,利用基因的突变体及多个不同根肿病抗性的白菜材料分析了基因内部可能与抗病相关的结构域。Hatakeyama 等精细定位了位于A03 染色体140 kb 区域的,随后利用新构建的群体把区间缩小至64 kb,结合拟南芥同源序列及参考基因组信息,克隆出序列信息,与已报道的序列一致,最终确认和是同一个基因。付鹏宇精细定位了位于染色体A03上60 kb 区域的,在抗根肿病和感病白菜材料中利用PCR 扩增出序列全长,另外,把基因转化到拟南芥中,对获得的阳性植株进行抗病性鉴定及根部组织细胞学形态观察,最终结果验证了抗4 号生理小种。伴随着大白菜全基因组数据库的公布,以及更多A 基因组亚种重测序以及从头测序工作的进行,抗病基因的定位与克隆工作将在白菜类蔬菜作物中加速开展起来,基因的功能研究工作也将得到进一步的提升。
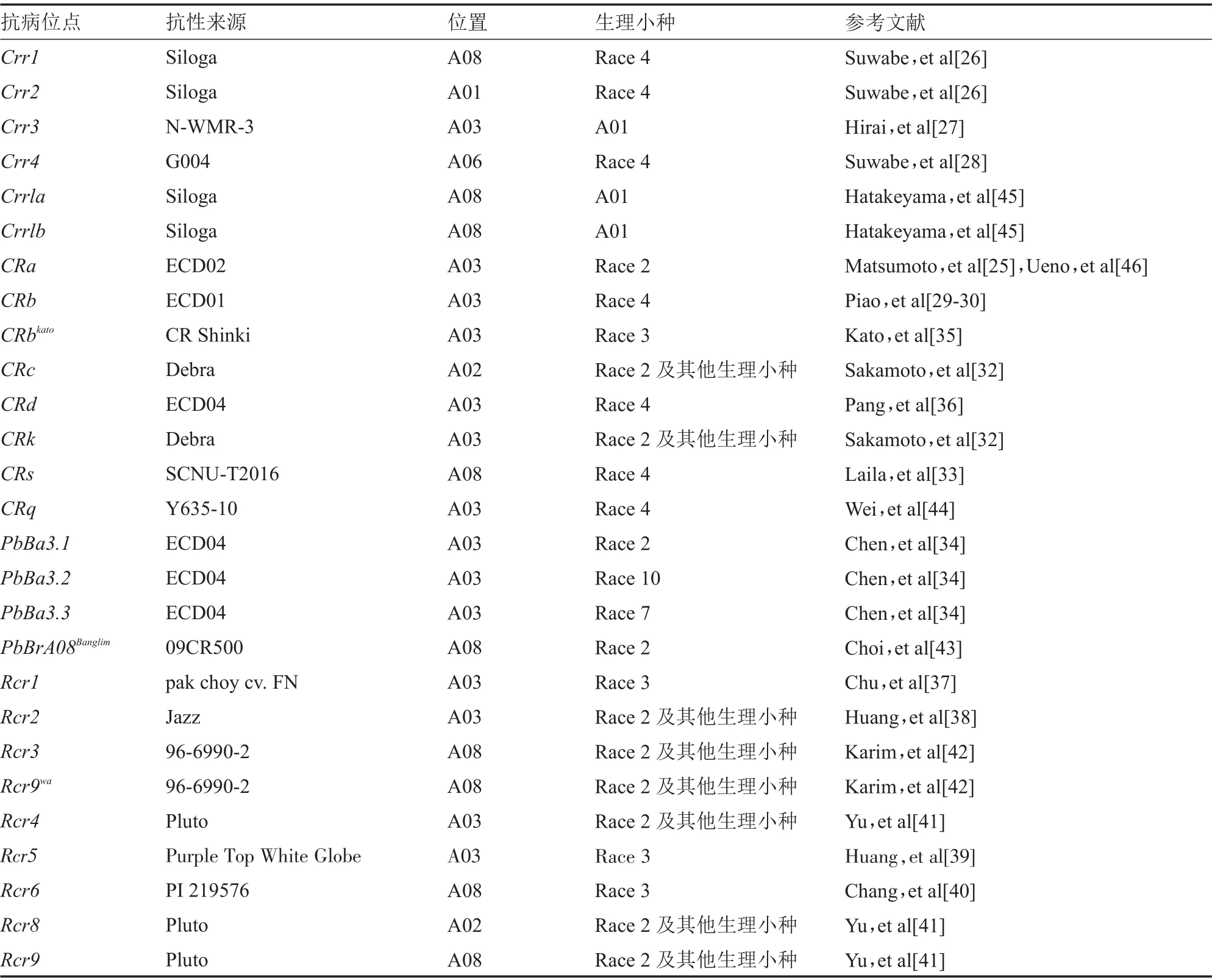
表1 白菜类抗根肿病基因位点及标记信息
2.2 甘蓝抗根肿病QTL定位研究
结球甘蓝(L. var.,CC=18)简称甘蓝,近年来研究人员对甘蓝品种进行广泛的根肿病抗病性鉴定,结果发现感病的甘蓝品种占大部分,而根肿病抗性资源很少。由于接菌的生理小种及研究的抗源材料的不同,甘蓝抗根肿病遗传规律的研究结果也不同,目前大多数研究认为甘蓝中的根肿病抗性是由几个主效位点和微效位点控制的数量性状,司军等认为甘蓝根肿病抗性受到3 对以上基因控制,为不完全隐性。
迄今,甘蓝中关于抗根肿病位点精细定位的报道较少,目前也没有克隆出抗病基因。Nagaoka等利用不同根肿病抗性的甘蓝DH 系构建分离群体进行QTL 定位,结果定位到了5 个QTL 位点,其中1 个主效QTL 位点的LOD 值为13.7,能够解释47%的表型变异。Lee 等利用Genotyping By Sequenceing(GBS)技术,通过构建F群体进行甘蓝抗根肿病QTL 定位,一共获得了抗2 号生理小种的1 个主效QTL 位点CRQTL-YC 和抗9 号生理小种的 2 个 主 效 QTL 位 点 CRQTL- GN_1 和CRQTL-GN_2,其中抗2 号生理小种的CRQTL-YC位点能够解释47.1%的表型变异。孙超等通过比较基因组学手段,在甘蓝中同源克隆出白菜的和基因,鉴定出1 个可能与抗根肿病相关的位点。张小丽筛选出1 个抗根肿菌4 号生理小种的甘蓝近缘野生种B2013,通过与感病品种杂交,后代抗病鉴定结果表明其抗病性状是多基因控制的数量性状,且存在显性的主效基因,并通过QTL-seq将抗病QTL 定位在甘蓝C09 染色体560 kb 区域。Peng 等利用SNP 基因芯片在甘蓝中检测到23 个与抗根肿病相关的QTL 位点,可解释6.1%~17.8%的表型变异。Dakouri 等通过BSR-seq 技术将甘蓝抗根肿病3 号生理小种的主效基因定位到C07 染色体41~44 Mb 区间内,通过比较区间内基因组的注释结果,进一步将TIR-NBS-LRR 类型基因和确定为候选基因。
2.3 甘蓝型油菜根肿病抗性研究
甘蓝型油菜(L.AACC=38)是通过白菜与甘蓝种间杂交,随后加倍形成的异源四倍体,种内遗传变异非常少,对根肿病抗性较差,整体上缺乏高抗或抗病品种。传统遗传学研究认为,甘蓝型油菜的根肿病抗性是由1~2 个独立的显性基因控制,这些显性基因在大量抗性材料中都存在。Chiang 等对甘蓝型油菜抗根肿病研究发现,1 号和3 号生理小种的抗性均受单一显性基因控制,而5 号生理小种的抗性是通过2 个隐性基因遗传的。余青青等以人工合成甘蓝型油菜为材料,对根肿病抗性进行遗传规律研究,发现该油菜的抗性受到一对显性基因控制。Rahman 等发现甘蓝型油菜抗根肿病基因受1~2 个显性基因控制。Diederichsen 等利用ECD04 和DH 群体发现1 个控制甘蓝型油菜根肿病抗性的主效基因和至少2 个隐性基因。Werner 等应用7 种不同的根肿病生理小种,定位到位于8 条染色体上的19 个QTL,这些QTL 对于不同的根肿病菌生理小种具有特异性。目前研究人员已经在甘蓝型油菜上发现一些抗病基因或QTL 位点,这些抗病位点大多与白菜和甘蓝存在一定的同源关系,控制根肿病抗性的少数几个起关键性作用的显性基因很有可能来源于白菜的A 染色体组。
2.4 萝卜根肿病抗性研究
与白菜、甘蓝和甘蓝型油菜相比,萝卜(L. RR=18)的根肿病抗病性整体上更强。Diederichsen 等研究发现大部分萝卜品种对根肿菌的抗性很有可能是由单个显性位点控制的。Akaba 等以日本萝卜为材料,接种根肿病2号生理小种,利用9 种类型的染色单体附加系进行研究,结果表明,根肿病抗性基因存在于萝卜的1条染色体上,不排除存在微效基因的影响。Kamei等用日本萝卜和中国萝卜杂交的95 个F群体进行萝卜根肿病抗病位点的QTL 分析,使用AFLP 和SSR 标记构建连锁图谱,结果表明该萝卜的抗性由或与其紧密连锁的基因控制,该基因与拟南芥第3 条染色体顶端区域的基因同源。Gan 等利用抗病萝卜品种BJJ 和感病品种XNQ 杂交获得的F群体,通过RAD-seq 技术定位到了分别位于8 号和9 号染色体上的5 个与根肿病抗病相关的QTL 位点,分别命名为、、、和。
3 十字花科栽培作物抗根肿病机制研究进展
3.1 抗病R基因在植物抗病中的作用
当病原体入侵时,植物自身会产生有效的机制来识别和应对,植物抵抗病原体的模式有2 种:由起初的病原体相关分子模式(PAMP)触发的免疫(PAMP-triggered immunity,PTI)和随后的效应因子触发的免疫(effector-triggered immunity,ETI)。植物抗病基因(disease resistance gene,)编码的R蛋白可以识别相应的PAMP 和效应因子,基因通常含有核酸结合位点(nucleotide binding site,NBS)、亮氨酸重复(leucine rich repeat,LRR)结构,可以直接或间接地与病原菌的效应因子结合,从而激发ETI 免疫反应,这些基因使植物能够识别外源病原体从而激活诱导防御。
迄今,根据基因和蛋白的序列和结构特点可将基因分为分为5 大类型:NBS-LRR(nucleotide binding site plus leucine richrepeat )、LRR-TM(leucine-rich repeat plus transmembrane receptor)、STK(serine-threonine kinase)、RLK(recepetor-Iike kinase)和SA-CC(signal anchorplus coiled-coil),其中NBS-LRR 类型基因在植物抗病基因中占据着重要的地位,根据N 端结构特征,可以将其分为2 个亚家族(TIR-NBS-LRR 和non-TIR-NBS-LRR)。目前,已经在白菜上克隆出来的4 个根肿病抗性基因、、和都具有核苷酸结合位点(nucleotide binding site,NBS)、亮 氨 酸 富 集(leucine-rich repeat,LRR)和TIR(drosophila toll or animal interleukin-1 receptors)结构域。在白菜中基因是特异的、单一的、显性的,而甘蓝中存在的基因则具有更连续的抗性,包括主基因和次级基因的各种组合。
3.2 植物激素在根肿病反应中的作用研究
植物激素在与病原菌互作过程中起着重要的作用,其中水杨酸(salicylic acid,SA)、茉莉酸(jasmonic acid,JA)和乙烯(ethylene,ET)是植物抵御外界胁迫信号转导途径中的重要调控因子,能帮助植物抵御病原菌的侵染;而生长素(indole-3-acetic acid,IAA)、细胞分裂素(cytokinin,CTK)和脱落酸(abscisic acid,ABA)等可能在某些病原菌侵染寄主时起到促进作用。
在拟南芥中至少存在2 种应答病原菌侵染的信号转导途径:一种是抵抗活体营养型病原菌侵染的水杨酸信号转导途径,另外一种是茉莉酸和乙烯依赖的途径,主要是抵抗腐生型病原菌的侵染。水杨酸和茉莉酸/乙烯信号转导途径在十字花科作物抵抗根肿菌侵染的过程中起着重要的作用,并且外源喷施SA 和JA 可以适当减轻植株根肿病发病症状。Lovelock 等对花椰菜根肿病研究发现,与对照相比,在受侵染的根部喷施SA 可以促进一些病程相关基因-及-等上调表达,减少植株根部肿瘤的形成,同时喷施SA 和JA 时,能显著降低感病植株的发病症状。Ning 等在研究根肿病侵染结球甘蓝的过程中发现,在抗感材料都受到根肿菌侵染后,与SA 和JA 信号途径相关的基因在抗病材料中上调表达,而在感病寄主中下调表达。Galindo 等对不同根肿病抗性甘蓝型油菜在根肿病侵染后的7、14、21 d 进行转录组测序,结果发现在抗病材料中与免疫功能相关的许多基因都与SA介导的反应有关。Ji 等对根肿菌侵染后的大白菜进行差异表达基因的研究发现,在16 个候选基因中有2 个是与SA 信号通路相关,同时还发现在SA处理后抗病寄主中SA 水平上升以及这2 个SA 信号通路相关的候选基因表达上调,说明SA 信号路径参与了白菜与根肿菌的互作过程。
多数研究认为植物激素包括IAA 和CTK 促进根肿病菌侵染寄主,在侵染过程中,IAA 和CTK 信号途径均表现活跃,引起受侵染的植株根部IAA 和CTK 含量显著上升,IAA 水平的上升可导致植株的细胞壁松弛、细胞伸长,最终可造成植株根部形成肿瘤。Xu 等对接种根肿菌后的甘蓝型油菜研究发现,接菌后3 d 和7 d 植株根部游离的IAA 水平提高,同时与IAA 合成相关的一些基因在根肿菌侵染早期被诱导表达,另外,向接种后的植株喷施外源IAA 可显著提高根毛的侵染率,而阻碍IAA传导途径可以有效减轻植株发病症状。
3.3 其他抗性机制在根肿病反应中的研究
近年来,伴随着一些高通量测序技术的发展与应用,为研究十字花科作物根肿菌侵染过程中涉及防御反应相关的代谢和信号网络应答机制提供了有效措施。谭翀通过对大白菜根部接种前后差异基因的表达分析发现,根肿菌侵染后的根部与胼胝质合成相关的基因均下调表达,与胼胝质降解相关的基因上调表达,说明胼胝质可能参与大白菜响应根肿菌的侵染过程。最近,Chen 等对大白菜近等基因系进行转录组分析,发现PAMPs、钙离子泵入、激素信号、PR、转录因子及细胞壁重塑等相关基因在不同抗感材料之间呈现差异表达,说明这些基因在根肿菌侵染初级阶段的寄主-病原菌互作过程中起作用。Yuan 等对根肿菌侵染不同时期的大白菜进行比较转录组学的研究,结果发现与防御酶、生长素、病程相关蛋白、抗病蛋白、细胞壁组织与生物合成、和转录因子相关的基因与根肿病抗性有关。Zhang 等对抗病甘蓝野生种及感病青花菜进行了转录组测序,结果显示,寄主对根肿菌的防御反应在早期被诱导且相关途径在侵染14 d 时受到抑制,与细胞壁成分、硫甙葡萄糖苷合成及植物信号转导相关的基因均在侵染早期表达,且大多数基因在抗病材料中上调表达。Verma 等利用miRNA 测序手段对甘蓝型油菜根肿病的发生过程进行了研究,结果发现差异表达miRNA 的靶基因主要涉及转录因子、激素相关基因及植物防御反应相关基因等。
4 展 望
十字花科作物作为我国栽培总量最大的蔬菜和油料作物,高发的根肿病严重影响着产量。因此,选育抗病品种及探索抗病机制显得尤为重要。中国现阶段白菜类蔬菜、甘蓝类蔬菜、甘蓝型油菜、萝卜育种中的基础研究环节相对薄弱,除了在大白菜中根肿病抗性基因报道的较多,在甘蓝和油菜中抗根肿病基因研究目前仍然还是处于QTL 初级定位阶段,还未有明确的基因被报道,根肿病抗性的分子机制尚不完全清楚。随着生物技术的发展,利用生物技术手段选育抗病品种及解析抗病机制已经成为今后的研究趋势,为在基因水平上认识植物与根肿菌的互作机制创造了有利条件,目前已有很多研究人员通过组学等测序手段研究预测了根肿菌侵染后植物体内基因或蛋白间可能的调控机制,但是关于这些机制研究的内容还是处于初级阶段,关于功能的研究较薄弱。因此,需进一步加大对基础研究的投入力度,建立更准确的十字花科作物根肿病生理小种的鉴别体系,明确国内病原生理小种的分布情况。同时结合高通量测序技术、数量性状定位及生物信息学分析手段,加快对抗性基因的克隆及功能研究。
