东祁连山高寒草甸土壤理化性质对海拔和坡向的响应及其与植被特征的关系
2022-10-25何国兴杨冬阳张德罡韩天虎潘冬荣柳小妮
李 强, 何国兴, 文 铜, 杨冬阳, 张德罡, 韩天虎,潘冬荣, 柳小妮
(1.甘肃农业大学草业学院/草业生态系统教育部重点实验室,甘肃 兰州 730070;2.甘肃省草原技术推广总站,甘肃 兰州 730000)
作为草地生态系统的重要组成部分,土壤是植物生长发育的基质,直接影响植被群落的结构和生产力[1]。土壤养分是草地植物养分的主要来源,其含量差异和平衡关系及土壤物理结构是调控植物生长和分布的关键因子[2]。土壤生态化学计量比表征了生物个体、种群的养分限制乃至生态系统的结构和功能[3-4],如碳氮比(C/N)和碳磷比(C/P)反映土壤氮、磷供应能力和分解速率,而氮磷比(N/P)反映了土壤氮、磷元素供应的限制[5-6]。贺金生等[3]系统的论述了生态化学计量比从个体到生态系统的统一理论及其养分限制理论,而陈云等[5]和孙小妹等[6]对我国典型生态脆弱区和天然草原生态化学计量方面的研究成果进行了梳理。研究发现,地形因素通过影响土壤与环境之间物质与能量的交换,且与其他成土因素共同影响土壤养分的分布与积累,进而影响着土壤生态化学计量特征的变化[1]。海拔和坡向是山地生态系统主要的地形因子,通过影响山地生态系统的水热环境进而影响区域内土壤养分分布格局和化学计量特征比[7]。因此,探索草地土壤理化性质和生态化学计量特征,揭示植物与土壤养分之间的关系,进而更好地认识养分元素的循环和平衡机制[8],了解地形因子变化过程中土壤养分含量和生态化学计量特征的变化规律,对评估土壤质量和合理管理草地具有重要的生态学意义。
祁连山是我国西北重要的生态安全屏障[9-11],而高寒草甸是祁连山的主要草地类型之一,对祁连山地区的生态系统稳定性起着举足轻重的作用[12]。目前,针对祁连山不同海拔、植被类型的土壤养分和生态化学计量特征开展了大量研究发现[13-16],如马剑等[13]和高海宁等[14]研究发现,祁连山北麓土壤C/N、C/P 随海拔升高总体呈下降趋势,且均在中高海拔处最高,植被类型、海拔、坡向和盖度是影响土壤碳(C)、氮(N)、磷(P)含量和化学计量比的主要因素,但是针对高寒草甸系统的对海拔和坡向变化过程中的土壤理化性质和生态化学计量比的变化特征的研究尚未见报道,海拔和坡向地形因子的互作对土壤理化性质和生态化学计量比的影响有助于揭示区域高寒草甸的养分匮缺和养分限制,对合理的管理高寒草地具有重要的意义。
因此,本研究以东祁连山高寒草甸为研究对象,通过分析不同海拔和坡向下土壤理化性质含量和生态化学计量比变化规律及其与植被的关系,以期为实现高寒草甸的合理管理和生态保护提供科学的数据参考。
1 材料与方法
1.1 研究区概况
研究区(102°26′31″~102°55′01″E,37°07′23″~37°17′53″N)位于甘肃省天祝藏族自治县抓喜秀龙乡的东祁连山金强河上游,海拔2600~4300 m。该区域属于高寒湿润气候,全年≥0 ℃的积温平均为1360 ℃,年平均气温-0.1~0.6 ℃,年均蒸发量1483~1614 mm,年平均降水量446 mm,气候寒冷潮湿,平均相对湿度55%[16]。研究区主要的草地类型是高寒草甸,土壤类型以亚高山草甸土、山地黑钙土、山地草甸土为主,且pH为6.94~8.17。
1.2 样地设置与样品采集
2020 年7 月30 日—8 月2 日,在东祁连山金强河流域选取了7 个不同海拔(2800 m、3000 m、3200 m、3400 m、3600 m、3800 m 和4000 m),同一海拔设置2个不同坡向(阳坡、阴坡)样地,采样区域均为禁牧区,每个样地随机各选取3 个10 m×10 m 的采样区域(表1)。
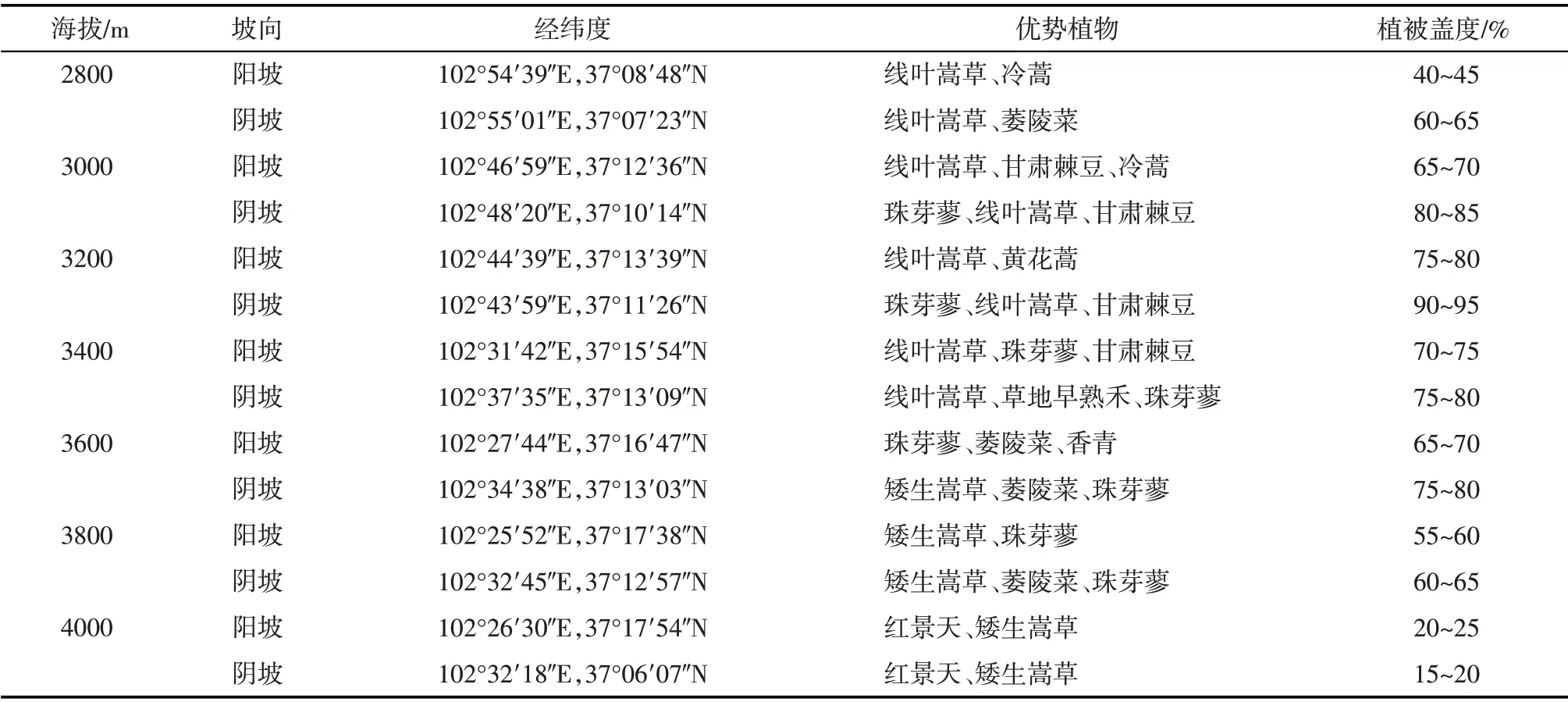
表1 样地基本信息Tab.1 Basic information of sample points
每个采样区域按对角线设3 个50 cm×50 cm 的样方,即3 个重复。采卷尺和针刺法用测定每个样方中植被的草层高度和总盖度及不同种植物的高度和分盖度,然后分物种采集地上生物量(齐地面刈割,放入样品袋中),称鲜重后,带回实验室于105 ℃烘箱中杀青2 h 后,60 ℃下恒重,称重。草样刈割后,采用直径5 cm 土钻采集表层0~30 cm 土样用于测定土壤养分含量,同时,用环刀取土壤样品测定土壤容重和含水量。
1.3 土壤理化性质测定
土壤容重和含水量分别采用环刀法和烘干法测定;土壤持水性参照由中国科学院南京土壤研究所物理研究室出版的《土壤物理性质测定方法》[17]。
土壤pH 和电导率测定:取部分风干土,过100目筛子,称取10 g,加入50 mL 去离子水,在全自动摇床上150 rpm振荡1 h,静至10 min,采用酸度计和电导率仪测定土壤pH和电导率。
全效养分测定:取部分风干土壤样品,过80 目筛,然后进行研磨,使粒径小于0.15 mm,待测。土壤有机碳(SOC)测定采用全自动碳分析仪(Multi N/C 2100S/1,Analytik Jena AG,Germany),土壤全氮测定采用凯氏定氮法、全磷测定采用钼锑抗比色法、全钾测定采用氢氧化钠熔融-火焰光度计法[17]。
速效养分的测定:将部分阴干土过100目筛子,将土壤样品进行研磨,使粒径小于0.25 mm,取土壤样品测定土壤速效氮(AN)、速效磷(AP)和速效钾(AK)。土壤速效氮采用扩散吸收法测定[17],土壤速效磷采用钼锑抗比色法测定(由于研究区土壤有机质含量较高,加入活性炭脱色测定效果不佳,故采用高锰酸钾氧化-葡萄糖还原法的方法进行脱色处理,详细脱色处理步骤参照张德罡等[18]的草原速效磷测定方法),土壤速效钾采用醋酸铵-火焰光度计法测定[17]。
1.4 数据处理与分析
使用Excel 2010软件进行数据整理和绘图。使用SPSS 21.0 软件进行方差分析和T 检验。使用SPSS 21.0和Canoco 5.0进行聚类和冗余分析(RDA)研究高寒草甸不同海拔和坡向土壤理化性质和生态化学计量比的异质性及其与植被的关系。
2 结果与分析
2.1 不同海拔和坡向下高寒草甸植被特征
由表2可知,随着海拔升高,植被盖度先升高后降低,阳坡和阴坡均在海拔3200 m 处达最大值(80.00%和96.67%);同一海拔,除了海拔4000 m 之外,阴坡植被盖度均高于阳坡,且海拔2800 m、3000 m、3200 m和3400 m的阴坡植被总盖度显著高于阳坡。高寒草甸草层高度和地上生物量变化规律与植被盖度基本类似。
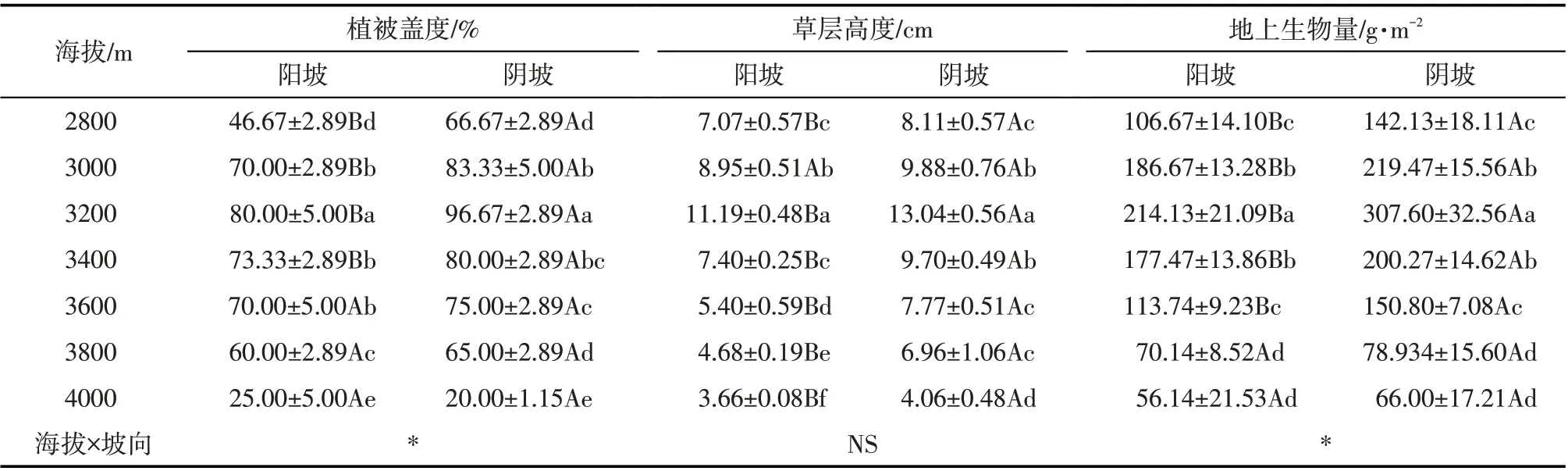
表2 不同海拔和坡向下高寒草甸植被特征Tab.2 Vegetation characteristics of alpine grasslands at different altitudes and aspect
2.2 不同海拔和坡向下高寒草甸土壤容重、含水量、pH和电导率
海拔和坡向的交互效应对土壤容重、含水量、pH和电导率有显著影响(P<0.05)(图1)。随着海拔的升高,土壤容重呈先降低后升高,阳坡和阴坡均在海拔3200 m处达最小值(0.68 g·cm-3和0.66 g·cm-3)(图1a);同一海拔梯度上,阴坡土壤容重均低于阳坡。随着海拔的升高,土壤含水量呈先升高后降低,阳坡和阴坡分别在海拔3600 m(52.18%)和3200 m(64.36%)处达最大值(图1b);同一海拔梯度上,阴坡土壤含水量均高于阳坡,除海拔3600 m外,其他海拔梯度阴坡与阳坡之间差异显著(P<0.05)。随着海拔的升高,阳坡土壤pH 呈先升高后降低再升高,阴坡呈先降低后升高的变化趋势,阳坡的最大值和最小值分布在海拔3000 m(7.80)和3800 m(6.63)处,而阴坡在海拔3800 m 处达最小值(6.67)(图1c);同一海拔梯度上,除海拔2800 m、3800 m和4000 m 外,阳坡土壤pH 均高于阴坡,且海拔3000m、3200 m、3600 m 和4000 m 阴坡与阳坡的土壤pH之间差异显著(P<0.05)。随着海拔的升高,除海拔4000 m外,土壤电导率在阳坡和阴坡呈先升高后降低(图1d),阳坡和阴坡土壤电导率分别在海拔3200 m(249.00 μS·cm-1)和海拔3000 m(195.83 μS·cm-1)处达最大值;同一海拔梯度上,除海拔2800 m和3000 m 外的其他海拔的阳坡土壤电导率均高于阴坡,且差异显著(P<0.05)。
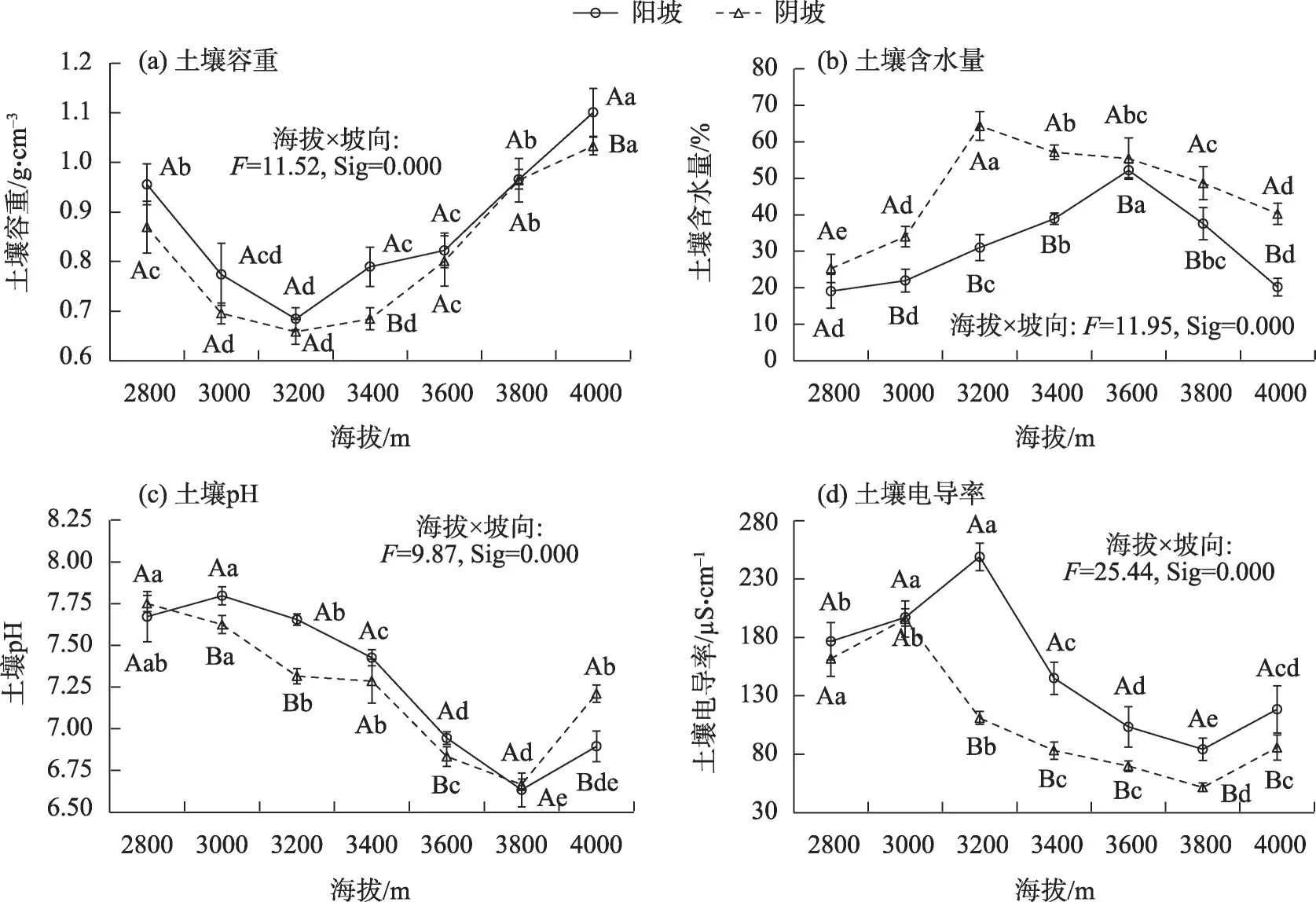
图1 不同海拔和坡向下高寒草甸土壤容重、含水量、pH和电导率Fig.1 Soil bulk density,water content,pH,and conductivity of alpine grasslands at different altitudes and aspect
2.3 不同海拔和坡向下高寒草甸土壤养分含量变化特征
海拔和坡向的交互效应对土壤有机碳、全氮、全磷和全钾含量有显著影响(P<0.05)(图2)。随着海拔的升高,土壤有机碳在阳坡和阴坡呈先升高后降低的变化趋势(图2a),阳坡和阴坡土壤有机碳在海拔3200 m 处达最大值(85.66 g·kg-1和99.26 g·kg-1);同一海拔梯度上,海拔3200 m 以下阴坡土壤有机碳均高于阳坡,且海拔2800 m和3000 m阴坡与阳坡之间差异显著(P<0.05)。土壤全氮随海拔和坡向的变化规律与土壤有机碳类似(图2b)。随着海拔的升高,土壤全磷在阳坡和阴坡呈先降低后升高的变化趋势(图2c),阳坡和阴坡土壤全磷在海拔3800 m处达最小值(0.77 g·kg-1和0.54 g·kg-1);同一海拔梯度上,除海拔4000 m 外,其他海拔梯度阳坡土壤全磷均高于阴坡。随着海拔的升高,土壤全钾在阳坡和阴坡呈先升高后降低的变化趋势(图2c),阳坡和阴坡土壤全钾分别在海拔3600 m(12.24 g·kg-1)和3800 m(14.45 g·kg-1)处达最大值;同一海拔梯度上,海拔3200 m、3400 m 和3600 m 梯度阳坡土壤全钾含量高于阴坡,其他海拔梯度阳坡土壤全钾低于阴坡。
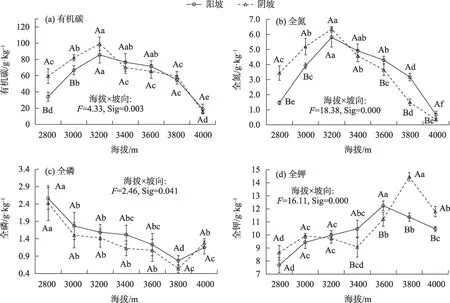
图2 不同海拔和坡向下高寒草甸土壤有机碳、全氮、全磷、全钾变化特征Fig.2 Changes of soil organic carbon,total nitrogen,total phosphorus and total potassium of alpine grasslands at different altitudes and aspect
土壤碱解氮随海拔和坡向的变化规律与土壤有机碳类似(图3a)。随着海拔的升高,土壤速效磷在阳坡和阴坡呈先升高后降低的变化趋势(图3b),均在海拔3200 m 处达最大值(24.62 mg·kg-1和28.75 mg·kg-1);同一海拔梯度上,阴坡土壤速效磷高于阳坡,除海拔4000 m 外,其他海拔均差异不显著(P>0.05)。随着海拔的升高,土壤速效钾在阳坡和阴坡呈先升高后降低的变化趋势(图3c),均在海拔3200 m处达最大值(286.61 mg·kg-1和221.06 mg·kg-1);同一海拔梯度上,除海拔3600 m和4000 m外,阴坡土壤速效钾低于阳坡。
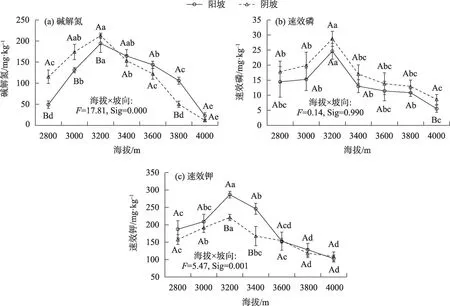
图3 不同海拔和坡向下高寒草甸土壤碱解氮、速效磷、速效钾变化特征Fig.3 Changes of soil alkali-hydrolyzable nitrogen,available phosphorus and available potassium of alpine grasslands at different altitudes and aspect
2.4 不同海拔和坡向下高寒草甸土壤化学计量比
不同海拔和坡向的高寒草甸土壤C/N 处于14.55~38.13之间(图4a),随着海拔的升高,土壤C/N在阳坡和阴坡均呈先降低后升高的变化趋势,阳坡土壤C/N在海拔3200 m处达最小值(14.55),阴坡土壤C/N在海拔3400 m处达最小值(15.20);同一海拔梯度上,海拔2800 m、3000 m 和3400 m 阳坡的C/N高于阴坡,其他海拔阳坡的C/N 低于阴坡,且海拔2800 m 和4000 m 的阴坡与阳坡土壤C/N 差异显著(P<0.05)。不同海拔和坡向的高寒草甸土壤C/P处于12.61~87.94 之间,随着海拔的升高,阳坡和阴坡土壤C/P 均呈先升高后降低的变化趋势,阳坡和阴坡土壤C/P 在海拔3800 m 处达最大值(75.24 和87.91);同一海拔梯度上,除海拔4000 m 外,阳坡土壤C/P均低于阴坡(图4b)。不同海拔和坡向的高寒草甸土壤N/P 处于0.58~5.01 之间,土壤N/P 的变化规律与C/P类似(图4c)。
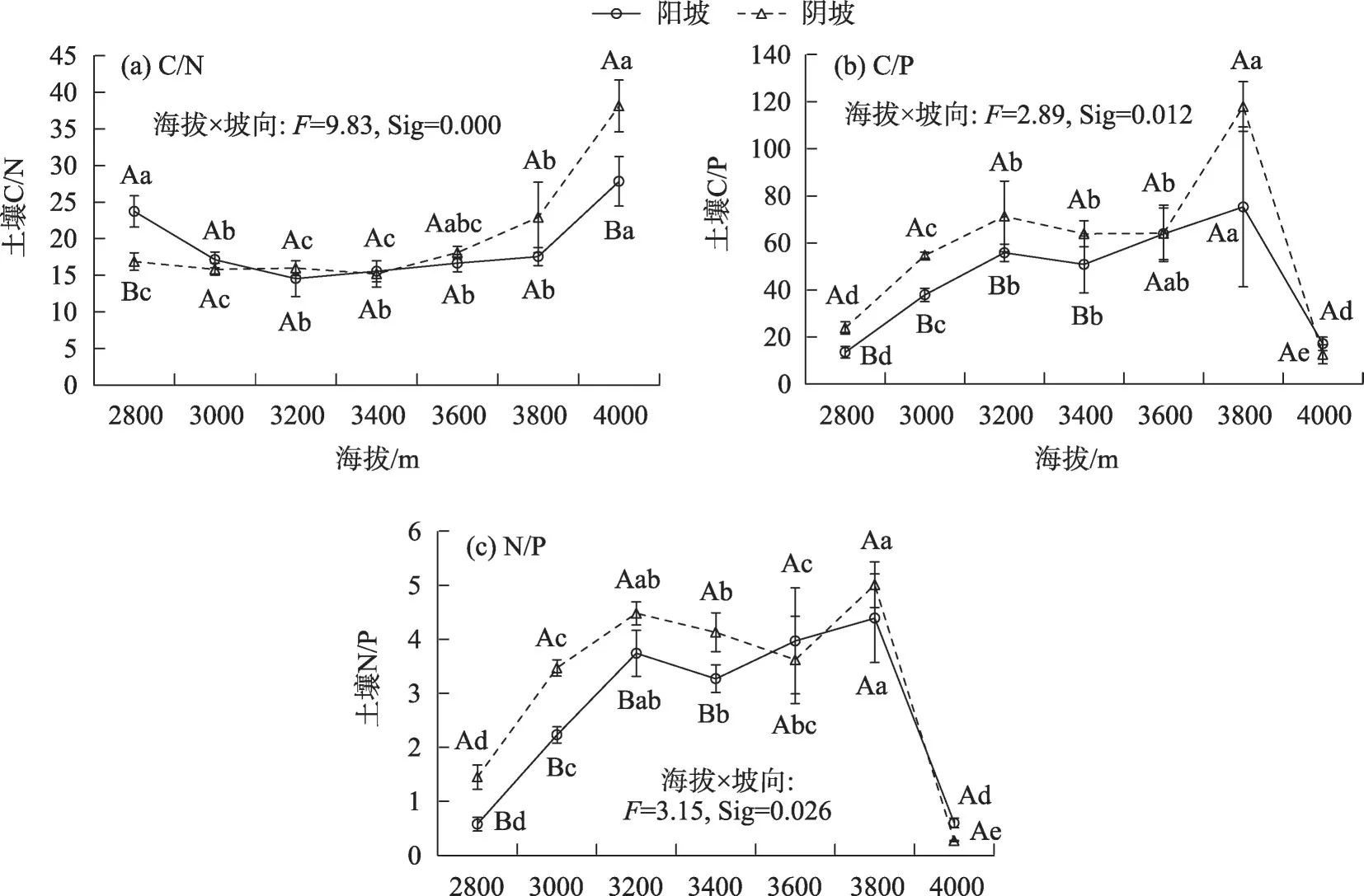
图4 不同海拔和坡向下高寒草甸土壤化学计量比变化特征Fig.4 Changes of soil stoichiometric of alpine grasslands at different altitudes and aspect
2.5 土壤因子与植被特征关系及基于植被和土壤因子的聚类分析
将土壤因子与植被特征进行RDA分析发现(图5a),第一轴和第二轴的总贡献率为95.18%,具有生物学统计意义。即前两轴可以比较完全的反映出高寒草甸土壤与植被的信息,以第一轴为主。由图5a 可知,高寒草甸植被特征受土壤容重、全氮和速效磷的影响较大,且土壤容重对植被特征的影响呈负相关,全氮和速效磷的影响呈正相关。
基于本文植被特征、土壤理化性质和生态化学计量比指标,对不同海拔和坡向处理进行聚类分析发现(图5b),高海拔(3600 m、3800 m 和4000 m)和低海拔区域(2800 m)的坡向之间类似性高于中海拔区域(3000 m、3200 m和3400 m)。
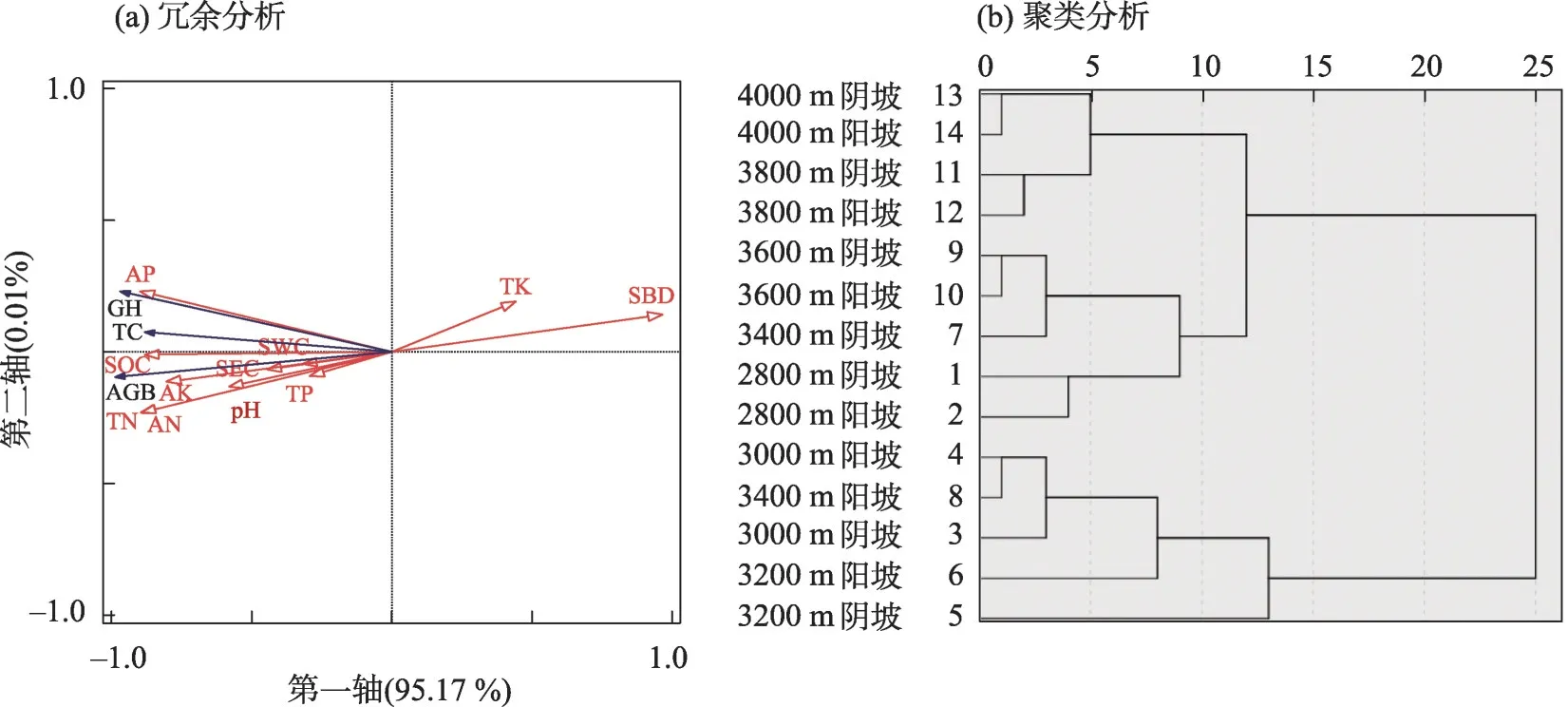
图5 土壤因子与植被特征冗余分析及基于植被和土壤因子的聚类分析Fig.5 Redundancy analysis of soil factors and vegetation characteristics and cluster analysis based on vegetation and soil factors
3 讨论
土壤是植物生长发育的基础载体,是各类生态环境因子与生物群落经过长期相互作用的产物[19],而土壤理化性质不仅仅是衡量土壤肥力的重要指标[20],还是土壤质量评价的基础[19]。海拔和坡向作为重要的地形因素,能够通过改变环境因子(气温、年降水量、年积温、植被群落组成)影响着土壤理化性质[19-20]。
3.1 土壤物理性质
土壤容重和含水量是反映土壤物理性质的重要指标;土壤pH 不仅影响着土壤养分循环和酶活性[21],而且还参与了土壤中的许多化学反应和过程[22];土壤电导率是衡量土壤养分含量的重要参数,能够准确地反映土壤质量。刘西刚等[20]研究发现,土壤含水量随海拔升高先升高后降低,而土壤容重随海拔升高逐渐降低。吕宸等[23]研究发现,半阴坡土壤容重大于半阳坡,土壤含水量阴坡大于阳坡。冯挺等[24]研究发现,土壤含水量、电导率随着海拔的升高呈先增加后降低趋势,阴坡土壤电导率小于阳坡。本研究发现,随着海拔的升高,土壤容重和pH 先降低后升高,土壤含水量和电导率先升高后降低;同一海拔,大部分海拔阳坡土壤容重、pH和电导率高于阴坡,但是阳坡土壤含水量低于阴坡,这与车明轩等[25]等研究结果类似。由于研究区高寒草草甸主要分布在2800~4000 m,在3200 m 以下,随着海拔的升高,草地植被盖度增加,同时气温下降,潜在蒸发量减少,冰雪融化进一步促使土壤含水量上升,草地植被根系活性增强,土壤容重和pH降低;当海拔超过3400 m后,低气温不利于植物的生长,冰雪融化水较少,土壤含水量逐渐下降,导致植被盖度下降和植物根系活力减缓[22]。同一海拔,阳坡接收到的太阳辐射显著高于阴坡,蒸发量差异导致土壤容重、含水量、电导率和pH 的坡向差异。
3.2 土壤养分
土壤有机碳是土壤肥力的直接体现,其含量与分布特征对区域土壤碳、氮、磷、钾的分布具有显著的影响[26]。土壤氮和磷是植被生长发育以及繁殖过程中最重要的养分元素[27]。金章利等[28]经过3 a定位试验发现,高寒草甸土壤碳、氮、磷均随着海拔的升高先升高后降低。本试验中,土壤有机碳、全氮、全钾、碱解氮、速效磷和速效钾含量随海拔的升高呈先升高后降低,全磷呈先降低后升高;同一海拔,阳坡的土壤全磷(除海拔4000 m)和速效钾(除海拔3600 m 和4000 m)高于阴坡,海拔3200 m以下阳坡的土壤有机碳、全氮、碱解氮和速效磷低于阴坡,这与金章利等[28]等研究结论存在差异,这可能是全磷分布的地域性差异造成的。随着海拔升高,气温下降,促使植物凋落物和土壤有机质增加,进一步加速了土壤微生物呼吸和微生物对土壤养分的利用效率,即一定范围内海拔的升高,提高了土壤有机碳和氮的含量[29]。海拔相同时,低海拔区域坡向对水热的二次分派明显,高海拔区域,气温过低,坡向对水热的分派减弱[30]。在高海拔,受气温较低的限制,减缓了土壤有机质的利用和矿化率,导致土壤有机碳和氮含量降低[31]。
3.3 土壤生态化学计量比
土壤生态化学计量可以反映土壤内部养分循环状况,对陆地生态系统的固碳过程具有很强的调控作用[32],具有重要的生态指示作用[33]。由于环境因子差异,不同海拔和坡向土壤碳、氮、磷的总量变化很大,从而使土壤C/N、C/P 和N/P 存在较大的空间变异性[34]。本研究发现,不同海拔和坡向的高寒草甸土壤C/N处于14.55~38.13之间,除海拔4000 m外,其他样地的C/N 均低于25,即有利于碳转化;随海拔的升高,土壤C/N 先降低后升高,同一海拔,海拔2800 m、3000 m 和3400 m 阳坡的C/N 高于阴坡,其他海拔阳坡的C/N 低于阴坡,这与高海宁等[14]研究结果相同,而与陈涵兮等[1]研究结果存在差异,主要的原因是陈涵兮等[1]研究区域属于低海拔区域。本研究发现,不同海拔和坡向的高寒草甸土壤C/P处于12.61~87.94之间,低于我国陆地生态系统平均值(105),也就是说研究区土壤碳、磷有效性较高。研究表明,当N/P<10时,植被生产力受氮素限制,当N/P>20 时,植被生产力受磷素限制[29,35]。本研究不同海拔和坡向的高寒草甸土壤N/P 处于0.27~5.01之间,随着海拔的升高,N/P 先升高后降低,也就是说高寒草甸生产力主要受土壤氮限制,低海拔和高海拔区域更为严重;同一海拔,大部分海拔阳坡土壤N/P低于阴坡。
3.4 影响高寒草甸植被特征的关键土壤因子
植被和土壤是高寒草地生态系统的重要组成部分,且相互影响相互制约[36]。相关研究发现,草地植被变化引起的土壤理化性质的改变将反作用于植物本身,尤其是土壤营养元素(氮、磷、钾等)、土壤水分等对植物性状、凋落物分解等均有显著影响,从而进一步影响植被的可持续性[37]。本研究中,基于RDA分析发现,土壤容重、全氮和速效磷是影响高寒草甸植被的关键土壤因子,这与李海云[35]在祁连山高寒草甸植被与土壤互作关系的研究结果类似。聚类分析将不同海拔和坡向的高寒草甸划分为两类,其中一类为海拔3000 m 的阴坡和阳坡、3200 m的阴坡和阳坡、3400 m的阴坡和阳坡,另一类为2800 m 阴坡和阳坡、3600 m 阴坡和阳坡、3800 m阴坡和阳坡以及4000 m阴坡和阳坡,也就是说,海拔3000 m和3400 m是该区域草地植被和土壤特征发生变化的临界线。
海拔和坡向等地形因子的变化,显著的改变了高寒草甸植被特征和土壤物理结构、养分含量和生态化学计量比,进一步形成了高寒草地山地生态系统的复杂性和异质性。因此,在高寒草地管理过程中,应该充分考虑地形因素(海拔和坡向)对草地的影响,制定合理的管理制度和措施,促进高寒草地生态系统的可持续发展。
4 结论
基于不同海拔和坡向东祁连山高寒草甸土壤理化性质及与植被特征关系研究结果表明:
(1)东祁连山高寒草甸土壤含水量、电导率、有机碳、全氮、全钾、碱解氮、速效磷、速效钾含量、C/P和N/P 随海拔的升高呈先升高后降低的变化趋势,土壤容重、全磷和C/N呈先降低后升高的变化趋势。
(2)同一海拔,大部分海拔梯度阳坡的土壤土壤容重、速效钾、电导率和全磷高于阴坡,阳坡的土壤含水量、速效磷、C/P和N/P低于阴坡,海拔3200 m梯度以下阳坡的土壤有机碳、全氮、碱解氮和C/N低于阴坡。
(3)基于对N/P 比的研究发现,高寒草甸类草原的初级生产力主要受土壤氮限制,且低海拔和高海拔区域尤显突出。
(4)结合RDA 和聚类分析发现,土壤容重、全氮和速效磷是影响高寒草甸植被生长的关键土壤因子,且海拔3000 m和3400 m是该区域草地植被和土壤特征发生变化的临界线。建议在高寒草甸类草原的管理过程中,应充分考虑海拔和坡向的分异性特征。
