PKR基因表达高效阻断Marc-145细胞系建立及其对PRRSV增殖的影响
2022-10-21肖跃强王文秀孙培姣沈志强
肖跃强,杨 慧,于 雪,王文秀,孙培姣,魏 凤,唐 娜,沈志强
(1.山东省滨州畜牧兽医研究院,山东 滨州 256600;2.山东绿都生物科技有限公司,山东 滨州 256600)
干扰素诱导的双链 RNA活化蛋白激酶(interferon-induced,double-stranded(ds)RNA-activated kinase,PKR)是一种丝氨酸-苏氨酸激酶,因组织类型和分化程度不同,PKR 在非活化细胞基础水平存在差异[1]。PKR在宿主病毒防御中起着关键作用,病毒感染后复制产生的双链RNA(dsRNA)能够活化PKR形成二聚体并磷酸化,之后催化其底物-真核生物翻译起始因子2α(alpha subunit of the eukaryotic initiation factore,eIF2α)的磷酸化[2-3],导致病毒蛋白的合成受到抑制,从而产生抗病毒活性。
猪繁殖与呼吸障碍综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸障碍综合征病毒(PRRS virus,PRRSV)引起一种以母猪繁殖障碍和各日龄猪呼吸障碍为特征的接触性传染病。猪肺泡巨噬细胞(porcine alveolar macrophages,PAMs)是PRRSV的天然宿主靶细胞,本课题组前期研究表明,PRRSV或灭活病毒粒子体外感染PAMs早期能够抑制PKR的活化,以促进病毒的增殖[4],因此,在早期感染阶段PKR激活下调应该是PRRSV避免抗病毒反应启动复制的必要条件。
PRRSV体外培养主要依赖于MA-104细胞及其衍生的Marc-145细胞[5],其中,Marc-145细胞是PRRSV体外培养应用最重要的细胞系,广泛用于科研与疫苗生产,但该细胞PKR基因与PRRSV相互作用机制尚不明确。本研究将在本实验室前期克隆获得Marc-145细胞PKR基因序列、构建融合EGFP标签真核表达载体基础上设计shRNA[6],并筛选能够下调表达PKR的shRNA,进而构建该shRNA稳定表达的Marc-145细胞系,研究PKR在下调状态下对PRRSV增殖的影响,为阐明PRRSV与PKR相互作用机制奠定基础。
1 材料与方法
1.1 细胞与病毒Marc-145、HEK293与Vero细胞、新城疫病毒(Newcastle disease virus,NDV)Mukteswar毒株、表达EGFP的重组病毒PRRSV XZ06a-EGFP[6],均由山东省滨州畜牧兽医研究院重点实验室保存;pSilencerTM2.1-U6 hygro载体由军事兽医研究所扈荣良研究员惠赠。
1.2 主要试剂Lipo2000、PageRulerTM预染蛋白Marker、GeneJET Plasmid小量制备试剂盒、GeneJET Gel Extraction试剂盒、GFP Tag mAb、HRP标记Goat anti-Mouse IgG等购自Thermo Scientific公司;Rabbit anti-PKR mAb(YE350)购自Abcam公司;Goat anti-Rabbit IgG购自苏州睿瀛生物技术有限公司;DMEM培养基、胎牛血清购自Gibco公司;限制性内切酶、T4DNA Ligase购自NEB公司;潮霉素B购自Amresco公司;PRRSV M蛋白mAb由哈尔滨兽医研究所蔡雪辉研究员惠赠。
1.3 shRNA设计与合成根据本实验室前期已经克隆获得的Marc-145细胞PKR基因序列设计shRNA,同时设已公布的阳性shRNA对照(Sigma-Aldrich、TRCN0000196400)、pSilencerTM2.1-U6 hygro载体为阴性对照。PKR shRNA与阳性shRNA对应的DNA由生工生物工程(上海)股份有限公司(上海生工)合成,序列见表1。

表1 PKR基因shRNA序列
1.4 shRNA表达载体的构建以BamHⅠ+HindⅢ双酶切pSilencerTM2.1-U6 hygro,根据试剂盒说明书纯化回收,与合成的shRNA经T4 DNA连接酶连接,转化大肠杆菌感受态细胞DH5α,涂布Amp抗性平板过夜培养筛选,随机挑取单菌落过夜培养后小量提取质粒,送上海生工测序验证。
1.5 PKR下调表达shRNA筛选将PKR shRNA表达质粒与本实验室前期构建的Marc-145细胞PKR基因融合EGFP标签的重组质粒pC1-PKR分别组合[6],共转染培置于12孔细胞培养板平铺面积约80% HEK293细胞,转染剂量为0.5 μg + 0.5 μg,同时设pC1-PKR(0.5 μg)、pC1-PKR+ pSilencerTM、pEGFP-C1+ pSilencerTM转染对照,具体转染方法根据Lipo2000转染试剂说明书进行。24 h 后弃去细胞培养基,无菌PBS溶液轻柔漂洗2次,加入200 μL SDS-PAGE上样缓冲液裂解细胞并收获,煮沸10 min,然后进行SDS-PAGE电泳并转印硝酸纤维素膜,以EGFP标签为靶标开展Western blot检测,筛选能抑制PKR表达的shRNA。
1.6 shRNA稳定表达Marc-145细胞系构建确定Marc-145细胞抗性筛选的最佳潮霉素B工作质量浓度,设100,200,300,400,500 mg/L共5个质量浓度梯度进行筛选(均2个重复),7~10 d后发现300 mg/L 及以上质量浓度全部致死细胞,因此选择300 mg/L 工作质量浓度筛选表达shRNA的Marc-145细胞系。选择对PKR表达有阻抑作用的shRNA表达质粒转染Marc-145细胞,同时设pSilencerTM空载体对照(NC),转染剂量为1.0 μg,抗性筛选并进行克隆纯化,构建PKR下调表达的Marc-145细胞系。
1.7 细胞系PKR表达检测及PRRSV感染待克隆纯化的细胞系F3代细胞、Marc-145细胞铺满12孔细胞培养板时弃去培养基上清,无菌PBS溶液轻柔漂洗2次,加入200 μL SDS-PAGE上样缓冲液裂解细胞,煮沸10 min,置于-20℃保存备用。将克隆纯化的细胞系F3代细胞、Marc-145细胞置于12孔细胞培养板培养,待细胞面积约80%时感染重组病毒PRRSV XZ06-∧EGFP,感染剂量为0.1 MOI,在感染48 h时弃去感染病毒后的培养基上清液,经无菌PBS漂洗、细胞裂解、煮沸获得样品,与未感染病毒的细胞样品同时进行SDS-PAGE并转印硝酸纤维素膜,Western blot检测细胞PKR、重组病毒M蛋白与EGFP的表达。同时,持续观察病毒对各细胞系细胞的致病变效应(cytopathic effect,CPE)。
1.8 NDV感染试验有研究表明当PKR基因表达被siRNA抑制后能促进NDV的增殖[7],因此,以中等毒力NDV Mukteswar毒株感染Marc-145细胞对照、克隆纯化的细胞系F3代细胞,感染剂量为1 MOI。同时,将细胞系细胞培养上清液处理Vero细胞,接种同样剂量NDV,以探究所构建细胞系shRNA是否激活干扰素通路,从而产生抗病毒活性。
2 结果
2.1 使PKR下调表达的shRNA的筛选经测序验证,成功构建了Marc-145细胞PKR基因shRNA、及阳性对照shRNA的表达载体,分别命名为pShRNA-143/295/1058/1296/1870S1C1。将构建的pC1-PKR与上述shRNA重组质粒共转染HEK293细胞,Western blot检测EGFP表达,检测结果见图1。与转染pC1-PKR、pC1-PKR+ pSilencerTM对照相比,PKR143、PKR295、PKR 1870S1C1均能够下调PKR的表达,以PKR 1870S1C1下调效率最高,而PKR1058却能够上调PKR表达,其原因不得而知。因此,本研究选择pShRNA-143/295/1870S1C1构建稳定表达细胞系。

M.PageRulerTM预染蛋白Marker;1.细胞对照;2.pC1-PKR转染;3~6.pC1-PKR+pSiRNA-143/1058/295/1870S1C1共转染;7.pC1-PKR+pSilenceerTM共转染;8.pEGFP-C1+pSilenceerTM共转染
2.2 稳定表达shRNA Marc-145细胞系构建经潮霉素B筛选,转染7 d后,细胞大部分死亡,随着时间延长,局部出现单个细胞克隆缓慢生长。待细胞克隆形成细胞簇后消化传代,进一步克隆纯化,并持续用潮霉素B筛选培养,最后获得与Marc-145细胞形态相似、生长速度相同的细胞系,分别命名为Marc-145∧143/295/1870S1C1/NC,构建的细胞系见图2。
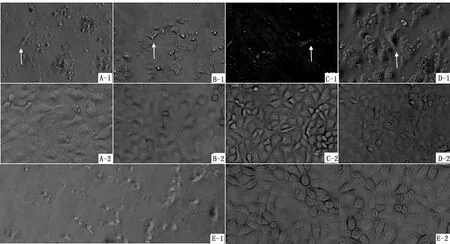
A.Marc-145∧143细胞系;B.Marc-145∧295细胞系;C.Marc-145∧1870S1C1细胞系;D.Marc-145∧NC细胞系;E.Marc-145细胞(1代表潮霉素B筛选存活细胞;2代表筛选后的细胞系或者正常Marc-145细胞)
2.3 细胞系PKR表达及PRRSV感染检测Western blot检测各细胞系PKR、重组病毒M蛋白与EGFP的表达结果如图3所示。可见,Marc-145∧143细胞系PKR的表达大幅度下调,下调比例在90%以上,Marc-145∧295、Marc-145∧1870S1C1细胞系PKR表达下调不显著,虽然SIGMA-ALDRICH公司对PKR-1870S1C1产品的说明显示其对A549细胞PKR基因表达的抑制效率在90%以上,载体阴性对照细胞系Marc-145∧NC PKR的表达未见显著变化;而重组病毒感染48 h后,接种Marc-145、Marc-145∧NC、Marc-145∧295后M蛋白与EGFP均获得高效表达,说明不影响PRRSV XZ06a-EGFP的增殖,而接种Marc-145∧143、Marc-145∧1870S1C1细胞后M蛋白与EGFP表达效率均大幅度降低,尤其是Marc-145∧143细胞。以上结果表明,所构建的细胞系Marc-145∧143、Marc-145∧1870S1C1可有效抑制病毒的增殖,而Marc-145∧1870S1C1细胞PKR表达效率未见明显下降,其原因有待阐明。

图3 shRNA稳定表达细胞系PKR表达与PRRSV XZ06a-EGFP感染后M蛋白与EGFP的表达
细胞系细胞感染重组病毒 PRRSV XZ06a-EGFP72~96 h,Marc-145∧143、Marc-145∧1870S1C1细胞均未呈现明显的CPE,而Marc-145、阴性对照Marc-145∧NC、Marc-145∧295细胞均呈现典型的CPE,进一步说明所构建的细胞系Marc-145∧143、Marc-145∧1870S1C1可有效地抑制病毒的增殖。
2.4 NDV感染试验NDV接种构建的细胞系培养上清处理的Vero细胞,出现典型病变后收获测定HA效价,与未进行任何处理的Vero细胞相比,病毒HA效价无任何差异,均为4 log2,表明构建的细胞系细胞培养上清对NDV在Vero增殖无显著性影响,说明细胞系未产生干扰素,抑制PRRSV的增殖。NDV感染Marc-145细胞、Marc-145∧NC/143/295/1870S1C1,24 h后Marc-145、Marc-145∧NC/295细胞全部出现细胞融合病变,Marc-145∧143/1870S1C1未出现典型的CPE,但48 h 后全部病变。分别收集感染24 h细胞培养上清与细胞并冻融破碎细胞(培养基同体积无菌PBS),分别测定HA效价,NDV感染细胞上清液HA效价无差异,均为1 log2,感染的Marc-145∧143细胞破碎后HA效价为5 log2,其他4个细胞系,虽然Marc-145∧1870S1C1在病变时间上与Marc-145∧143类似,但细胞破碎后HA效价无差别,为3 log2,说明Marc-145∧143细胞系促进了NDV的增殖,佐证了该细胞系PKR基因表达受到显著抑制。
3 讨论
shRNA在基因功能学研究领域是一种极其重要的工具,本研究通过构建U6启动子转录的shRNA表达载体,转染后经抗性筛选、细胞传代、克隆纯化获得了稳定表达shRNA并下调PKR表达的细胞系Marc-145∧143,PKR表达下调90%以上,PRRSV XZ06a-EGFP增殖受到强烈抑制,也说明PKR对PRRSV复制具有促进作用。同时,开展本研究时,根据Marc-145细胞PKR基因序列,选择序列可完全匹配的在售shRNA产品(sigma-Aldrich,TRCN0000196400)核心序列作为阳性对照,该产品在A549细胞对PKR下调效率高达97%,但在Marc-145细胞,以该核心序列shRNA构建的细胞系Marc-145∧1870S1C1对PKR的下调表达并不显著,但却可以抑制PRRSV重组病毒的增殖,M蛋白与EGFP表达均受到抑制,其中EGFP抑制程度与Marc-145∧143细胞相差无几,而M蛋白受抑制程度较Marc-145∧143细胞低很多。PRRSV感染后释放基因组RNA,在转录调控序列(transcription regulatory sequence,TRS)和RNA依赖的RNA聚合酶的作用下以病毒基因组为模板,采用不连续转录的方式转录出一系列共5′末端和 3′末端的负链亚基因组RNA,然后再以此为模板合成病毒亚基因组mRNA,最终翻译成病毒的结构蛋白[8-9]。PRRSV XZ06a-EGFP重组病毒EGFP基因是通过在病毒基因组ORF1b与ORF2之间引入ORF6(M蛋白)TRS及酶切位点后插入的,即EGFP与M蛋白基因是受同一个TRS序列调控。有研究表明[10],直接转染siRNA沉默Marc-145细胞PKR表达并不会影响PRRSV的感染,并且揭示PKR主要通过调节病毒基因转录促进病毒的复制,而不是病毒蛋白的合成。而本研究在重组病毒感染Marc-145∧143、Marc-145∧1870S1C1M后M蛋白与EGFP在受同一TRS调控且PKR表达受到抑制差异较大情况下,M蛋白与EGFP的表达受到抑制程度却不同,说明PKR 也可能通过调控PRRSV结构蛋白的表达促进病毒的增殖,这一推测也需要进行病毒非结构蛋白及其他结构蛋白的检测进一步加以论证。同时,另外一个细胞系Marc-145∧295与Marc-145∧NC对照相比,均不能下调PKR的表达,究其原因,可能是该shRNA容易发生脱靶效应,同时,重组病毒M蛋白与EGFP表达较Marc-145细胞差异不明显,进一步说明PKR对于PRRSV在Marc-145细胞增殖起到重要调控作用。
有研究表明,在干扰素-γ(interferon gamma,IFN-γ)处理PAMs与Marc-145细胞后加入PKR抑制剂2-氨基嘌呤(2-aminopurine,2-AP),PRRSV核酸复制得以恢复,接近病毒在正常细胞的复制,并且加入2-AP后PKR重新自细胞浆进入细胞核,且这种转移是因为PRRSV感染复制或恢复复制所致[11];HR等[12]以2-AP处理猪IFN-β作用的PAMs与Marc-145细胞,可以恢复病毒在Marc-145细胞的复制,但在PAMs无明显作用,说明猪IFN-β在2种细胞采用不同的抗病毒信号通路与PRRSV相互作用。本课题组前期研究显示,在PAMs细胞PRRSV通过抑制PKR活性促进自身增殖[4],而本研究通过shRNA下调Marc-145细胞PKR的表达能够抑制PRRSV增殖。同时,2-AP并不是一种PKR特异抑制药物,而是一种广谱的丝氨酸/苏氨酸激酶抑制剂[13],细胞内多种细胞激酶和磷酸酶参与病毒复制、组装、出膜等[14],因此,推测PKR下调表达也可能会导致其他激酶活化或抑制,并在一定程度参与了PRRSV的复制,但这一推测需要后续研究阐明。
为了排除构建的细胞系shRNA在转录过程中是否激活IFN通路而对PRRSV增殖产生影响,采用文献[15]方法,即将细胞系培养基上清液处理Vero细胞,然后接种NDV,通过观察CPE与测定病毒产量,发现细胞系培养基上清液对NDV感染复制无影响,排除了shRNA诱导细胞系产生干扰素的可能。也进一步佐证PRRSV在细胞系复制受到抑制与PKR下调表达相关。在HeLa细胞,PKR/eIF2α信号通路级联活化后NDV的复制受到抑制,当PKR基因表达被siRNA抑制后则能促进NDV的增殖[7],本研究构建的PKR下调表达的Marc-145细胞系同样促进了NDV的增殖,病毒培养时,与正常细胞相比,病变时间延后1倍,可能是因PKR表达水平的降低,病毒复制产生的dsRNA活化PKR/eIF2α信号通路级联作用强度有限所致。
综上,本研究成功构建可高效下调PKR表达的细胞系Marc-145∧143,能够抑制PRRSV的增殖,进一步揭示PKR在调控PRRSV的增殖中发挥重要作用,这种作用除通过调节病毒基因转录促进病毒增殖以外[10],还可能通过调控病毒蛋白的合成发挥促病毒增殖的作用。
