取食不同猎物对黄带犀猎蝽生长发育和繁殖的影响
2022-10-13陈苏怡李永青石安宪查世钰彩万志王孟卿
陈苏怡, 李永青, 石安宪, 查世钰, 彩万志, 王孟卿, 李 虎*
(1. 中国农业大学植物保护学院昆虫学系, 北京 100193; 2. 云南省昭通市植保植检站, 昭通 657100;3. 中国农业科学院植物保护研究所, 北京 100193)
我国天敌昆虫资源十分丰富,其中猎蝽科Reduviidae是半翅目Hemiptera昆虫中最大的捕食性类群。在已知的7 500余种猎蝽中,肉食性种类占87%以上,是重要的天敌昆虫资源[1]。目前环斑猛猎蝽Sphedanolestesimpressicollis、角轮刺猎蝽Scipiniasubula、红彩瑞猎蝽Rhynocorisfuscipes[2]、黄带犀猎蝽Sycanuscroceovittatus和大红犀猎蝽S.falleni等多种猎蝽正逐渐被开发和利用[3-7]。食物是天敌昆虫规模化饲养过程中的一个关键因子,亦是限制天敌捕食效果的因素之一,同时还决定天敌的饲养成本。因此,对可选猎物种类的评价对于衡量食物质量至关重要[8-9]。
黄带犀猎蝽,又名中黄猎蝽,属猎蝽科真猎蝽亚科Harpactorinae犀猎蝽属Sycanus,广泛分布于我国广西、云南、贵州、香港等南方省区,以及马来西亚、缅甸和印度等国家[10]。据文献记载,黄带犀猎蝽可作为农林害虫天敌,防治桉树、芒果树和降香黄檀树等林木上的鳞翅目、鞘翅目、膜翅目及直翅目等害虫[11-13]。近年有报道,在云南省江城县的玉米田中发现其捕食草地贪夜蛾Spodopterafrugiperda[6]。此外,本研究团队在广西壮族自治区那坡县玉米田也发现黄带犀猎蝽,且在云南省师宗县和巧家县玉米田进行人工释放后可见其捕食草地贪夜蛾和斜纹夜蛾Spodopteralitura幼虫。黄带犀猎蝽具有较广的取食范围和较强的存活能力,对农林业害虫有一定的制约能力,可成为农林害虫综合防治中的一种优势天敌。
猎物质量直接影响捕食者的生长发育[9]。研究可替代猎物种类是了解未来捕食者在室内规模化饲养潜力的重要一步,对今后天敌的应用有一定的参考价值。本研究以黄粉虫Tenebriomolitor幼虫,果蝇Drosophilasp.成虫、斜纹夜蛾幼虫和棉蚜Aphisgossypii若虫作为猎物,单头饲养黄带犀猎蝽初孵若虫至成虫(图1)。比较取食不同猎物的黄带犀猎蝽若虫生长发育历期、体重、成虫寿命和繁殖情况,评价不同猎物作为黄带犀猎蝽食物的适宜性,为推进其在生产中的应用奠定基础。

图1 黄带犀猎蝽若虫取食4种不同猎物Fig.1 The nymphs of Sycanus croceovittatus consume four different preys
1 材料与方法
1.1 供试虫源
黄带犀猎蝽起始种群为采自广西壮族自治区百色市那坡县百省乡百省村玉米田的成虫,在昆明云南恒草生物科技有限公司的人工气候室内配对后置于300 mL透明太空杯(上底直径7.5 cm,下底直径5.5 cm,高9.6 cm,厦门市永诚鑫塑胶有限公司)中,放入剪裁后的单格蛋托盘(纸浆模塑,约5 cm×5 cm×5 cm)供其产卵。
黄粉虫幼虫和残翅果蝇成虫从网上购得,分别用麦麸和卖家提供的饲料培养基饲养于人工气候室内;棉蚜若虫、斜纹夜蛾幼虫和斜纹夜蛾人工饲料均由云南恒草生物科技有限公司提供,其中斜纹夜蛾已在人工气候室内饲养15代以上。
除棉蚜外所有供试昆虫均饲养于人工气候室内,温度(25±1)℃,相对湿度为(60±5)%,光周期为L∥D=14 h∥10 h(下同);棉蚜饲养于云南昆明室外大棚内,寄主植物为南瓜Cucurbitamoschata苗,温度(22.5±5.5)℃,相对湿度(65±10)%,光照为自然光,中午高温时需使用遮阳网。
1.2 黄带犀猎蝽饲养方法
1.2.1卵的孵育
将24 h内产于蛋托盘上的卵置于塑料盒(长22.3 cm,宽15.3 cm,高9.0 cm,盒盖中央切割一块12.0 cm×6.0 cm长方形孔,罩上120目尼龙纱网)中,放置一块浸湿脱脂棉增加湿度,待其孵化,记录卵的孵化时间。
1.2.2若虫饲养
从3块卵期相同的卵孵化的健康初孵若虫中各取80头,用毛笔挑单头置于太空杯中,用纱网和橡皮筋封口,纱网上放置一块浸湿的脱脂棉提供水分。若虫按饲喂的猎物平均分为4组处理,即黄粉虫组、果蝇组、斜纹夜蛾组和棉蚜组。饲喂1~2龄若虫时需选取黄粉虫幼虫2 cm左右个体、果蝇成虫健康个体、斜纹夜蛾1~2龄幼虫和带棉蚜饱满个体的南瓜苗叶片,捕食者和猎物比例分别约为1∶2、1∶10、1∶5和1∶50;饲喂3~5龄若虫时选取黄粉虫幼虫3 cm左右个体、果蝇成虫健康个体、斜纹夜蛾3~4龄幼虫和带棉蚜饱满个体的南瓜苗叶片,捕食者和猎物比例分别约为1∶3、1∶20、1∶5和1∶200。每48 h更换1次猎物,保证各组猎物充足。每组处理20头黄带犀猎蝽若虫,重复3次。每 24 h观察记录黄带犀猎蝽若虫的发育进度,若蜕皮前死亡计入该发育阶段死亡数,在蜕皮24 h内用恒平FA2104电子分析天平称量(感量为0.1 g,上海舜宇恒平科学仪器有限公司)新蜕皮黄带犀猎蝽的体重,直到羽化为成虫。羽化失败或羽化24 h内死亡均记为羽化期死亡。
1.2.3成虫饲养
新羽化的成虫称重、记录性别后进行组内配对,单对转移至太空杯中,饲喂同种猎物(捕食者和猎物比例同上述高龄若虫),放入剪裁后的蛋托盘供其产卵,产卵后24 h内收集新产的卵,并放入新蛋托盘供其继续产卵,将卵转移至新的太空杯中,用纱网和橡皮筋封口,纱网上放置一块浸湿的脱脂棉增加湿度,待其孵化。在试验阶段,配对成虫中若雄虫在雌虫开始产卵前死亡,则从同一组种群内选择羽化日期相近的雄性个体补充配对。每24 h观察记录各项生物学指标,包括雌虫产卵前期、产卵日期、孵化日期、产卵量等,直至所有黄带犀猎蝽成虫死亡,记录成虫寿命。
1.3 试验设计与数据统计分析
不同猎物喂养试验同时进行。所有试验均在1.1所述人工气候室内进行,将每组处理的3次重复共60头个体放置在同一片区域,方便更换饲料和系统观察,4组处理平行排列于人工气候室内架子上。各生物参数计算方法如下:
各发育阶段死亡率=各发育阶段死亡数/20×100%;
成虫获得率=100%-∑各发育阶段死亡率;
单雌产卵量=∑单次产卵量;
卵孵化率=初孵若虫数/单次产卵量×100%;
各发育阶段死亡率服从二项分布,需将小于30%或大于70%范围内数据进行反正弦转换,以矫正其正态性和方差的均匀性。死亡率、发育历期和体重等种群生命参数在取食不同猎物组处理间的差异显著性采用单因素方差分析(F检验),平均数比较采用Tukey’s检验法(P<0.05)。下文2.4中成虫繁殖率等参数使用t测验比较成虫获得率高的两组处理。统计计算过程由Excel和GraphPad Prism 6.01软件实现。
2 结果与分析
2.1 不同猎物对黄带犀猎蝽各发育阶段死亡率的影响
单独取食黄粉虫、果蝇、斜纹夜蛾和棉蚜,黄带犀猎蝽均能完成世代发育(表1)。取食不同猎物的黄带犀猎蝽1龄若虫的死亡率没有显著差异(df=3, 8;F=0.808 6;P=0.523 8),2龄、3龄、4龄和5龄若虫不同处理间死亡率存在显著差异(2龄:df=3, 8;F=6.665;P=0.014 4;3龄:df=3, 8;F=49.66;P<0.000 1;4龄:df=3, 8;F=7.199;P=0.011 6;5龄:df=3, 8;F=31.56;P<0.000 1),其中2~4龄以棉蚜组死亡率最高,分别为(30.1±7.6)%、(30.0±0.0)%和(26.8±4.4)%;5龄若虫则为果蝇组死亡率显著高于取食其他猎物组,达到(58.3±9.3)%;羽化期死亡率各处理间无显著差异(df=3, 8;F=1.697;P=0.244 3)。取食不同猎物组的成虫获得率存在显著差异(df=3, 8;F=32.00;P<0.000 1),其中斜纹夜蛾组和黄粉虫组显著高于棉蚜组和果蝇组,其成虫获得率分别为(69.9±7.6)%、(50.0±7.6)%、(8.3±1.7)%和(5.0±2.9)%;取食黄粉虫组黄带犀猎蝽各龄期若虫死亡率、羽化期成虫死亡率和成虫获得率与取食斜纹夜蛾组无显著差异。
2.2 不同猎物对黄带犀猎蝽若虫发育历期的影响
取食不同猎物组的黄带犀猎蝽各龄若虫的发育历期存在显著差异(1龄:df=3, 217;F=29.88;P<0.000 1;2龄:df=3, 178;F=60.96;P<0.000 1;3龄:df=3, 153;F=60.15;P<0.000 1;4龄:df=3, 123;F=63.49;P<0.000 1;5龄:df=3, 81;F=24.82;P<0.000 1)。若虫发育历期:1龄期取食果蝇组和斜纹夜蛾组显著长于黄粉虫组和棉蚜组(表2);2龄和3龄期取食棉蚜组显著长于其他3组,取食斜纹夜蛾组最短,取食黄粉虫组与果蝇组间差异不显著;4龄期以取食棉蚜组最长,取食黄粉虫组和斜纹夜蛾组最短;5龄期取食果蝇组和棉蚜组显著长于取食黄粉虫组和斜纹夜蛾组;若虫总历期在取食各猎物之间均差异显著(df=3, 81;F=159.3;P<0.000 1),由长到短分别为棉蚜组(138.2±3.4)d、果蝇组(102.9±6.9)d、黄粉虫组(81.9±1.2)d和斜纹夜蛾组(70.3±0.6)d。
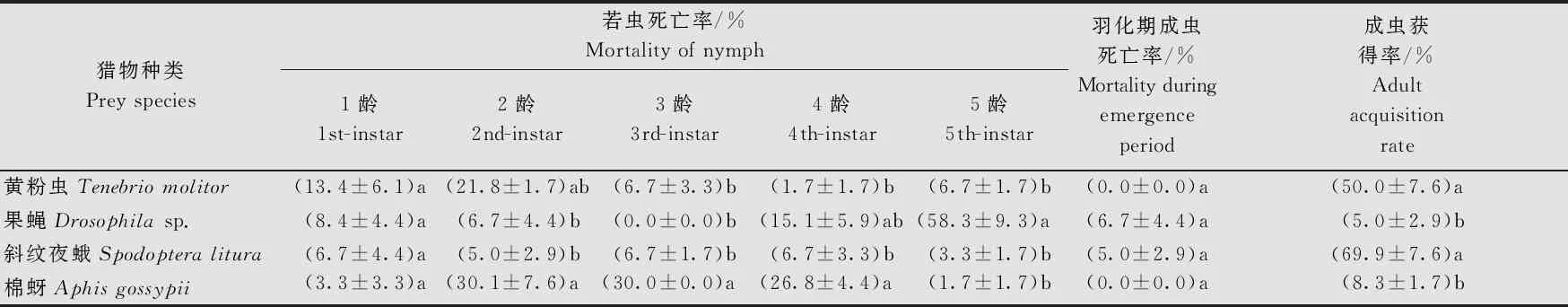
表1 不同猎物饲养条件下黄带犀猎蝽的各发育阶段死亡率和成虫获得率1)

表2 不同猎物饲养条件下黄带犀猎蝽若虫发育历期1)
2.3 不同猎物对各发育阶段黄带犀猎蝽体重的影响
取食不同猎物的黄带犀猎蝽2~5龄若虫和雌、雄成虫体重存在显著差异(2龄:df=3, 168;F=101.7;P<0.000 1;3龄:df=3, 158;F=211.0;P<0.000 1;4龄:df=3, 145;F=57.18;P<0.000 1;5龄:df=3, 117;F=29.82;P<0.000 1;雌成虫:df=3, 33;F=28.75;P<0.000 1;雄成虫:df=3, 35;F=7.194;P=0.000 7),均以斜纹夜蛾组体重最高,分别为(3.1±0.1)、(11.2±0.2)、(18.2±0.5)、(54.4±1.3)、(195.4±5.8)mg和(141.8±3.9)mg;除2龄若虫以果蝇组体重最低外,其他虫态均为棉蚜组最低(表3)。除此之外,取食同种猎物的黄带犀猎蝽雌成虫体重均高于雄成虫,其中果蝇组和棉蚜组体重差异较小,而黄粉虫组和斜纹夜蛾组差异较大。

表3 不同猎物饲养条件下黄带犀猎蝽各发育阶段的体重1)
2.4 不同猎物对黄带犀猎蝽成虫寿命、繁殖力和卵的影响
由于取食果蝇组和棉蚜组成虫获得率低于10%,样本量小,此部分仅比较样本量相对较多的黄粉虫组和斜纹夜蛾组。以黄粉虫和斜纹夜蛾喂养的黄带犀猎蝽,其成虫繁殖相关系数存在显著差异(雌成虫寿命:df=14, 16;F=2.463;t=4.709;P<0.000 1;雄成虫寿命:df=11, 22;F=1.329;t=1.590;P=0.121 3;产卵前期:df=15, 14;F=5.217;t=2.402;P=0.023 0;单雌产卵量:df=15, 14;F=5.664;t=1.050;P=0.302 4;卵期:df=29, 28;F=6.821;t=1.332;P=0.188 0;卵孵化率:df=9, 14;F=3.718;t=3.879;P=0.000 8)。取食斜纹夜蛾组雌、雄成虫寿命和产卵前期均短于取食黄粉虫组,分别为(31.6±3.0)、(39.2±2.7)d和(21.5±0.5)d,卵期较取食黄粉虫组长,为(20.0±0.1)d;取食黄粉虫组单雌产卵量为(274.8±52.4)粒,高于斜纹夜蛾组,但其卵孵化率仅(45.1±8.4)%,显著低于斜纹夜蛾组(76.5±3.6)%(表4)。相关记录图片见图2。
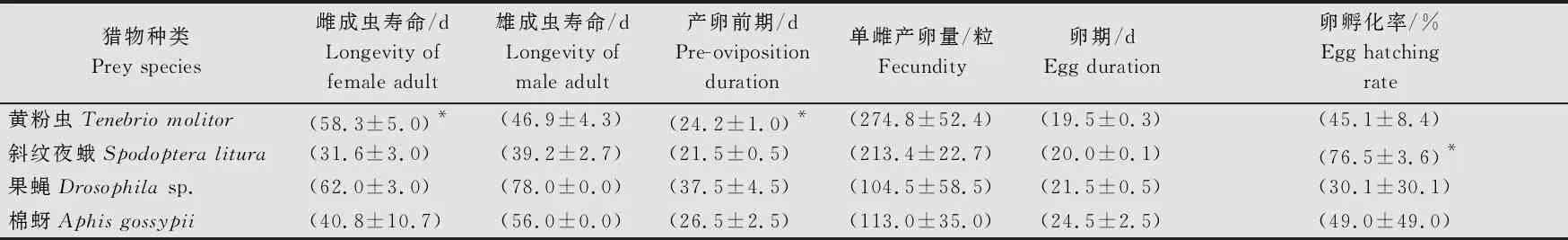
表4 不同猎物饲养条件下黄带犀猎蝽实验种群生命表参数比较1)

图2 黄带犀猎蝽成虫繁殖过程Fig.2 Reproduction process of the assassin bug Sycanus croceovittatus adults
3 结论与讨论
猎物类型不同其营养价值可能不同,不同发育阶段的捕食者对不同类型食物的适生性也是不同的,Morales-Ramos等认为不同种类的节肢动物含有的营养成分不同,特别是碳水化合物、脂肪和蛋白质的比例不同,这会影响捕食者的发育[14]。本研究结果表明,黄带犀猎蝽取食黄粉虫、果蝇、斜纹夜蛾和棉蚜均可完成世代发育,且可以继续繁殖,但猎物种类对其生存、生长发育和繁殖能力等有显著影响。取食果蝇和棉蚜的黄带犀猎蝽成虫获得率不到10%,显著低于取食黄粉虫和斜纹夜蛾。棉蚜仅适合黄带犀猎蝽1龄若虫取食。取食棉蚜的1龄若虫死亡率低且发育历期短,自2龄若虫开始死亡率显著上升。取食果蝇的黄带犀猎蝽1龄若虫死亡率较高且发育历期长,自2龄若虫开始死亡率降低,发育历期变短,但体重依旧偏小,至4龄若虫开始死亡率显著增加,发育历期延长,体重也比黄粉虫组和斜纹夜蛾组低。昆虫在遇到逆境时,例如猎物营养不足时会发生拒食现象,而取食量的减少会导致发育历期延长、不能正常产卵或产卵量降低等现象[15]。作者每次更换饲料时棉蚜和果蝇均有剩余活虫,不存在猎物不足问题,推测可能是由于随着黄带犀猎蝽龄期的增长,棉蚜和果蝇无法满足其生长发育对营养质量的需求。此外,作者还发现残翅果蝇爬行能力较黄带犀猎蝽1龄若虫强,这可能导致1龄若虫消耗过多能量用于追赶猎物。
与其他猎物相比,使用黄粉虫喂养黄带犀猎蝽时其低龄若虫死亡率较高,这可能是由于低龄若虫口器较小,而黄粉虫若虫体型较大且体壁较厚,低龄若虫捕食较为困难所致。用黄粉虫喂养二叉犀猎蝽Sycanusdichotomus,其1龄和2龄若虫发育历期显著长于用米蛾Corcyracephalonica喂养,用黄粉虫和米蛾混合喂养时,1龄和2龄若虫更倾向于食用米蛾幼虫[9]。在本研究中,使用黄粉虫喂养的黄带犀猎蝽高龄若虫和成虫具有死亡率低、若虫历期和卵期较短、体重以及单雌产卵量较大等特点,这与取食黄粉虫的二叉犀猎蝽高龄若虫体重和腿节长度大于取食米蛾的结果类似[9]。取食黄粉虫组黄带犀猎蝽单雌产卵量高达(274.8±52.4)粒,较同样室内饲养的环角犀猎蝽Sycanusannulicornis和大红犀猎蝽单雌产卵量更多[15-16],规模化扩繁潜力更强。
取食斜纹夜蛾的黄带犀猎蝽若虫死亡率较低,发育历期较短,体重最重,成虫获得率最高。且成虫产卵前期较短,卵孵化率较高。以菜青虫、斜纹夜蛾、小菜蛾和米蛾喂养的大红犀猎蝽的若虫期存活率分别为76.7%、69.3%、67.8%和57.3%[17],与取食斜纹夜蛾的黄带犀猎蝽的成虫获得率相近。以斜纹夜蛾幼虫为猎物较以桃蚜Myzuspersicae为猎物的红彩瑞猎蝽的种群生命表各项参数占优[18],且在桃蚜若蚜、斜纹夜蛾和烟青虫Helicoverpaassulta幼虫3种猎物同时存在的情况下,红彩瑞猎蝽成虫更喜欢捕食斜纹夜蛾3龄幼虫[19]。除此之外,使用甘蓝薄翅螟Crocidolomiapavonana喂养环角犀猎蝽较使用黄粉虫喂养若虫发育历期更短,高龄若虫和成虫体重更大[16],表明鳞翅目幼虫可作为促进犀猎蝽属昆虫生长发育和繁殖的良好饲料。Abdul等认为成虫寿命长可以促进雌成虫与雄成虫进行多次交配提高繁殖力[16],但本研究中使用斜纹夜蛾喂养的黄带犀猎蝽成虫寿命短于其他3组。Ahmad等研究发现同时使用黄粉虫和米蛾幼虫喂养二叉犀猎蝽时,其成虫的寿命更长,个体也更健康[9],今后可以考虑进一步研究不同猎物混合饲养,以弥补单独喂养斜纹夜蛾的不足。
通过单独喂养黄粉虫、果蝇、斜纹夜蛾和棉蚜4种不同的猎物得出黄带犀猎蝽生长发育和成虫繁殖力相关生物学参数表明,黄带犀猎蝽取食斜纹夜蛾时生长发育最快、繁殖力最强,优于其他猎物,可用于室内大规模喂养。此外,我们对野外种群的卵期记录发现3块卵的卵期均为16 d,而在这4组试验中黄粉虫组卵期最短,为(19.5±0.3)d,显然比野外种群卵期要长,其原因有待进一步研究。
