淹渍胁迫下枳砧温州蜜柑的形态特征和生理响应
2022-10-13李菲菲唐超兰蒋红梅陈鹏韩健郭玲霞许园园卢晓鹏
李菲菲,唐超兰,蒋红梅,陈鹏,韩健,郭玲霞,许园园,卢晓鹏
1.湖南省农业科学院园艺研究所,长沙 410125;2.湖南农业大学园艺学院,长沙 410128;3.湖南省洪江市农业农村局,洪江418100
柑橘是我国南方地区产量最大和分布最广的亚热带果树。由于多数柑橘产区春夏季节雨季较长,降雨量大,橘园常出现雨季积水浸根,甚至洪涝淹树等不同程度的淹渍胁迫。因此,了解柑橘淹渍胁迫后的树体状况及淹渍响应生理机制对于正确评估柑橘淹渍灾害影响及灾后恢复性生产十分重要。
淹渍胁迫对植株的影响主要表现为植株形态和生理改变,引起植株根系腐烂,进而造成植株无氧呼吸加剧[1]、叶绿素降低、光合性能下降[2]、细胞膜伤害、代谢紊乱等,最终使得植株生长受到抑制、萎蔫、局部坏死或整株死亡[3]。研究[4]表明,柑橘砧木幼苗淹渍后表现叶脉黄化,最终导致叶片萎蔫干枯;淹渍条件下不同柑橘砧木品种的耐涝性差异明显,资阳香橙、卡里佐枳橙和印度酸橘为耐涝砧木,枳为中度耐涝砧木,红橘、枸头橙和宜昌橙为不耐涝砧木[5]。枳砧脐橙1 年生苗在淹渍后根系受损、生长量减少,叶片叶脉变黄、脉间相对保持绿色,根系和叶片的解剖结构也相应发生变化[6]。淹渍降低柑橘幼苗的光合作用,影响其生长和生物量分配,引起叶片气孔关闭,降低根系的导水率和pH 值,但对叶片水势和相对含水量无影响[7-8]。柑橘在淹渍胁迫条件下积累活性氧造成细胞膜损伤,长时间淹渍胁迫使植株体内膜脂过氧化程度升高,并抑制某些蛋白质合成,造成可溶性蛋白含量下降、可溶性糖含量升高、抗氧化酶活性升高[6,9]。柑橘根系直接受到淹渍胁迫的伤害,导致根系活力降低,影响根系对养分的吸收和转运,造成C/N 失调[7],也有研究报道淹渍胁迫影响柑橘对Fe的吸收[10]。
我国柑橘产区春夏季节连续阴雨易导致橘园淹渍频繁发生,淹渍胁迫对柑橘坐果及果实发育、产量品质形成、花芽分化均有较大影响,是当前柑橘生产亟需解决的问题之一[5,11]。国内外学者围绕柑橘砧木如枳橙、枳、资阳香橙、酸橙等以及部分柑橘类幼苗分别开展了淹渍胁迫的相关研究[4-10],而温州蜜柑对淹渍胁迫响应机制的系统研究鲜有报道。因此,本研究以盆栽的枳砧‘市文’温州蜜柑为材料,从淹渍后温州蜜柑叶片和枳砧根系的形态学及解剖结构特征等表型变化着手,结合淹渍胁迫后植株的内在生理及生化指标变化,明确枳砧温州蜜柑在淹渍胁迫下的形态特征变化以及生理生化响应机制,以期为今后制定科学有效的栽培技术措施、降低胁迫灾害程度及柑橘耐涝良种选育提供理论指导。
1 材料与方法
1.1 试验材料
试验于2019 年5-6 月在湖南农业大学温室大棚(28°17'99″N,113°07'71″E)内进行。以栽植于黑色塑料桶(直径40 cm,高34 cm)中的9 年生枳砧‘市文’温州蜜柑(Citrus unshiuMarc.‘Ichifumi Wase’)为材料,栽培基质为V锯木屑∶V河沙∶V草炭=2∶1∶0.5。根据该地区春夏之交持续降雨规律性明显的特点,试验于5月16日开始,淹渍处理(waterlogging stress,WⅬ)采用双套盆法,在打孔的桶外再套1个无孔的黑色塑料桶,控制淹水水面超过土壤表面1 cm。淹渍历时设2、5、10、15、20 d共5个处理;同期设置的对照(CK)进行正常水分管理,使基质相对含水量维持在(70±10)%。使用便携式土壤水分监测仪监测水分含量,确保相对含水量稳定。以上每个处理3 次重复。大棚温湿度情况使用温湿度数据记录仪记录(图1)。
1.2 主要仪器与设备
TDR 便携式土壤水分监测仪,美国Spectrum technologies;DT-171温湿度数据记录仪,中国CEM;Pannoramic MIDI 数字切片扫描仪,匈牙利3DHISTECH;ⅬI-6400 携式光合仪,美国ⅬI-COR Inc;Multiskan Sky 全波长酶标仪,美国赛默飞;DDS-11A 电导率仪,上海雷磁;KDN-103F 自动定氮仪,上海纤检;Thermo-902 超低温冰箱,美国赛默飞。
1.3 形态指标的测定
每个处理达到设定淹渍时间后,对叶片和根系拍照并观察其形态学变化;将试验材料整株拔出,先用自来水冲去土壤和杂质,再用蒸馏水冲洗干净,然后用滤纸和吸水纸擦净材料上的水分,选取根尖粗度及成熟度相对一致的根段,放入乙醇-醋酸-福尔马林混合固定液(FAA 固定液)中保存,采用石蜡切片方法观察根系解剖结构[12]。
1.4 生理生化指标的测定
分别取CK 和淹水处理2、5、10、15、20 d 的植株细根和秋梢叶,其中叶绿素和相对电导率测定用新鲜叶片,其余均剪碎混匀、液氮速冻后放入-80 ℃超低温冰箱保存,矿质元素测定将试验材料的根、茎、叶分开,放入105 ℃烘箱杀青30 min 后,烘箱温度调至70 ℃烘干至恒质量,样品粉碎后过孔径0.15 mm筛待测,所有测定指标均设3次重复。
叶绿素含量采用乙醇浸提法[13];相对电导率采用电导率仪法[13];植物氮含量测定采用硫酸-过氧化氢消煮-蒸馏法,全磷(P)测定采用硫酸-过氧化氢消煮-钒钼黄比色法,全钾测定采用硫酸-过氧化氢消煮-火焰光度法,钙、镁、铁、锌测定采用微波消解仪进行消解,ICP 法测定[14]。每处理组结束后,于当日09:00-12:00 用便携式光合测定仪测定秋梢顶端第2~4 片叶的净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、胞间二氧化碳浓度(intercellular CO2concentration,Ci)及气孔导度(stomatal conductance,Gs)等光合参数[13],设置光量子通量密度1 200 μmol/(m2·s),选取叶片中部测定,对照和处理每株5片叶,3次重复。
脯氨酸含量采用磺基水杨酸提取,酸性茚三酮法测定;可溶性蛋白含量采用BCA 法测定[13];可溶性糖含量采用蒽酮比色法测定[13];丙二醛含量采用硫代巴比妥酸法测定[13];脱氢酶采用TTC 法[13];乙醇脱氢酶活性测定参考Ⅼi等[15]的方法。所用试剂均购自苏州科铭生物技术公司。
1.5 数据处理及分析
采用Microsoft Excel 2016 软件和Sigmaplot 10.0 软件进行数据处理和图表制作,采用Photoshop CC 进行图片的整合。运用SPSS 23.0 采用单因素(one-way ANOVA)和Duncan’s 法进行方差分析和显著性检验(α=0.05),用Pearson 法对叶片和根系间生理指标进行相关性分析。图表中数据为“平均值±标准误”。
2 结果与分析
2.1 淹渍胁迫对枳砧温州蜜柑外观形态和根系解剖结构的影响
春末夏初,大棚内日均温度25 ℃左右,相对湿度73%左右,温州蜜柑外观形态和根系解剖结构受淹渍胁迫的影响随淹渍时间的延长而加剧。由图2 可知,温州蜜柑淹渍2 d 根系和地上部无明显变化;淹渍5 d 根系有轻微异味,地上部变化不明显;淹渍10 d,根系轻微腐烂、变臭,地上部老叶出现不同程度的黄化、褶皱且叶脉变黄褐色,新叶失水、脉间黄化、叶尖焦枯,并伴随轻微落叶;淹渍15 d 根系腐烂加剧、部分韧皮部脱落、臭味加重,地上部枝条顶端出现焦枯,大量叶片边缘卷曲、干枯,树体近1/2的叶片自翼叶处产生离层、脱落;淹渍20 d 根系腐烂,韧皮部脱落严重,根系和近基部茎有酒味,整株叶片干枯褐黄,落叶数超过1/2,果实黄褐色形成僵果,部分自行掉落。温州蜜柑淹渍5 d 其根系解剖结构排列整齐,与CK 相比细胞更加饱满膨胀,淹渍20 d 其韧皮部与木质部分离(图3)。
2.2 淹渍胁迫对枳砧温州蜜柑叶绿素含量和光合性能的影响
淹渍胁迫导致温州蜜柑叶片中叶绿素及类胡萝卜素的含量显著降低。其中,温州蜜柑淹渍处理叶片中的叶绿素含量从第10 天始显著降低,且随胁迫时间的延长持续下降。温州蜜柑叶片中叶绿素a(chlorophyll a,Chl a)、叶绿素b(chlorophyll b,Chl b)和类胡萝卜素(carotenoid,Car)的含量在淹渍胁迫5~10 d 变化最为剧烈,特别是淹渍10 d 时各色素组分分别较同期CK 显著降低57.14%、47.73%、21.74%,且随淹渍时间的延长而持续下降,淹渍20 d降至最低值,此时温州蜜柑叶片中Chl a、Chl b、Car的含量分别较同期CK 降低86.07%、78.43%、86.36%,Chl 总含量仅是CK 的11.97%(图4)。淹渍胁迫对温州蜜柑叶片总叶绿素及各色素含量的影响趋势相同;此外,比较叶绿素a/叶绿素b(Chl a/Chl b)的变化可见,Chl a/Chl b 在淹渍胁迫后逐渐降低且Chl a 比Chl b 下降更快(图4E)。Car/Chl 在淹渍胁迫后显著升高,从而引起淹渍植株叶片的颜色更多地呈现黄色(图2A、图4F)。
由表1 可知,温州蜜柑淹渍处理10 d 时,光合作用也受到了强烈抑制,Pn、Gs、Tr分别较CK 显著下降76.01%、68.62%、59.36%,Ci浓度显著升高。

表1 淹渍胁迫10 d后枳砧温州蜜柑光合参数变化Table 1 Changes of photosynthetic parameters of satsuma mandarin leaves with trifoliate orange rootstocks after 10 d of waterlogging stress
2.3 枳砧温州蜜柑渗透调节物质对淹渍胁迫的响应
温州蜜柑根系中的渗透调节物质从淹渍2 d 开始持续下降,叶片中的渗透调节物质在淹渍10 d 后升高。淹渍树体根系中的脯氨酸(proline,Pro)含量随淹渍时间的延长呈下降趋势,淹渍20 d时较CK 显著下降91.44%(图5A);叶片中的Pro 含量显著升高,淹渍10 d 时较同期CK 上升60.90%,差异显著(图5B)。与同期对照相比,淹渍组根系中的可溶性蛋白(soluble protein,SP)含量自5 d 开始显著下降,淹渍10 d 时,较CK 显著降低79.61%,随后变化趋缓直至试验结束(图5C);而淹渍组叶片SP含量表现为先降低后增加的趋势,在淹渍5 d 内较同期CK 显著下降,自淹渍10 d 开始显著上升,淹渍20 d 较CK 增加了1.69 倍,差异显著(图5D)。处理组根系中的可溶性糖(souble sugar,SS)含量随淹渍时间延长而呈下降趋势,淹渍5 d 和20 d 分别较同期CK 显著降低44.96%、36.08%(图5E);处理组叶片SS含量至淹渍15 d 仍与CK 无显著性差异,淹渍20 d 才开始大量积累,并较同期CK 显著上升75.55%(图5F)。根系中的渗透物质先对对淹水胁迫作出响应,温州蜜柑根系中的渗透物质均在淹渍2~5 d 时表现出显著下降的趋势,而叶片中的渗透物质随淹渍时间的延长呈现显著上升趋势。
2.4 枳砧温州蜜柑根/叶膜脂过氧化代谢对淹渍胁迫的响应
淹渍逆境下温州蜜柑根/叶的丙二醛(malondialdehyde,MDA)及相对电导率(relative electrical conductivity,EⅬ)均呈现动态变化。温州蜜柑淹渍处理后,叶片MDA 含量表现为随处理时间延长呈逐渐增加的趋势(图5H)。自淹渍10 d 起,叶片的MDA 含量开始显著增加,达到CK 的1.66 倍;到淹渍15、20 d时,MDA 含量分别是CK 的2.25、2.53 倍,差异显著;根系中的MDA含量表现出下降-上升-下降的变化趋势(图5G)。温州蜜柑叶片的EⅬ淹渍后10 d 开始显著升高且随处理时间的延长表现稳定,在淹渍15 d最高,为同期CK的3.35倍(图5I)。
2.5 枳砧温州蜜柑根系无氧呼吸酶系统对淹渍胁迫的响应
淹渍胁迫下,温州蜜柑根系活力明显降低,无氧呼吸酶活性在淹渍早期急剧升高而后随淹渍时间的延长呈下降趋势。脱氢酶(dehydrogenase,DHA)活性是反映根系活力的重要指标。淹渍后温州蜜柑根系的DHA 活性总体呈下降趋势,且从淹渍10 d 开始显著低于CK,直至淹渍处理结束(图6A)。淹渍胁迫后温州蜜柑根系的ADH 活性呈先升高后降低的趋势。乙醇脱氢酶(alcohol dehydrogenase,ADH)活性在淹渍2、5 d 分别是CK 的1.54 倍、1.21 倍,自淹渍10 d 开始较CK 显著下降78.57%,而后随淹渍时间的增加而持续下降(图6B)。
2.6 叶片和根系生理生化指标的相关性
淹渍胁迫后,叶片与根系的抗逆理化指标多数呈极显著负相关。根系Pro 和SP 与叶片Pro、SP、SS、MDA 和EⅬ呈负相关,根系的SS 与叶片的Pro、MDA 和EⅬ也呈负相关;根系的MDA 和叶片Pro 呈极显著正相关(r=0.669)(表2)。
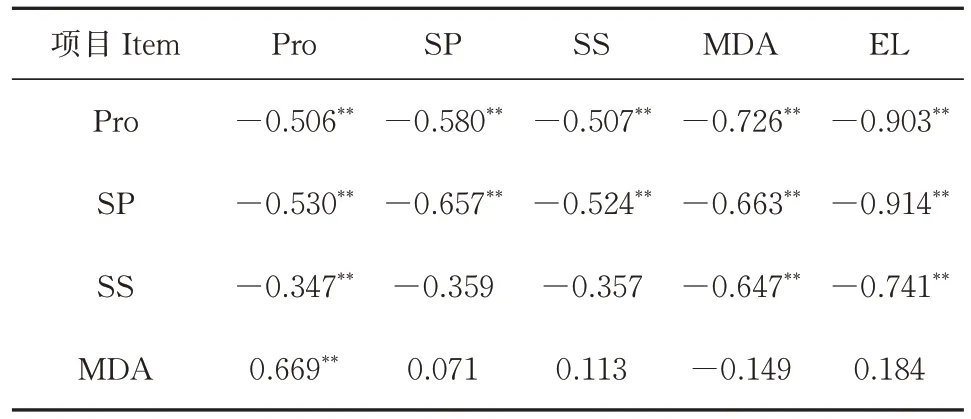
表2 淹渍胁迫下枳砧温州蜜柑叶片和根系生理生化指标的相关系数Table 2 Correlation coefficients of the physiological and biochemical indexes between leaves and roots of satsuma mandarin with trifoliate orange rootstocks under waterlogging stress
2.7 淹渍胁迫对枳砧温州蜜柑树体矿质营养积累的影响
淹渍胁迫导致枳砧温州蜜柑根系中P、K、Ca、Mg 含量减少,茎、叶中的各矿质元素含量无显著变化。根系中P 含量在淹渍5 d 和20 d 时均显著降低,20 d 时较CK 降低40%(图7D);根系中K 含量在淹渍5 d 时无显著变化,20 d 时较CK 显著降低(图7G);与对照相比,淹渍植株茎、叶中的P、K含量无显著性变化(图7E、F、H、I)。根系中的Ca、Mg 含量在淹渍20 d 时均显著下降,分别较CK 下降35.84%和65.7%;淹渍处理下茎、叶中的Ca、Mg 含量无显著变化(图8)。温州蜜柑淹渍后,根、茎、叶中的N、Fe、Zn含量波动较小(图7、图8)。
3 讨论
3.1 淹渍胁迫下枳砧温州蜜柑表型特征和叶片光合指标的变化
淹渍条件下植物气体扩散受阻、叶绿素降解、光合速率下降、产生无氧呼吸,干扰植物正常的生理代谢及生长发育,为了适应淹渍条件,保障正常的生理代谢功能,植株地上部和地下部形态通常会发生改变[16-17]。前人的研究表明,柑橘砧木及脐橙幼苗的叶片在淹渍胁迫时先是叶脉失绿黄化而脉间保持绿色,之后逐步出现黄化、萎蔫等症状[4,6];其中,红橘和枸头橙幼苗分别在淹渍处理的第13 和14 天开始出现萎蔫症状[5],枳砧‘大分四号’温州蜜柑幼树在淹渍4 d开始出现可见症状[18],而枳砧脐橙幼苗在淹渍40 d 才出现淹水胁迫症状[6]。本研究中枳砧‘市文’温州蜜柑在淹渍处理5 d 时叶片无明显可见症状,淹渍10 d 时出现不同程度的失绿黄化、萎蔫及轻微落叶等症状,推测不同柑橘类型淹渍症状出现的时间可能与其耐涝性强弱有关[4-6]。枳砧脐橙幼苗在淹渍胁迫15 d 时叶片外观及微观结构均无明显变化,但在淹渍10 d 时,根皮层细胞连接松散,并出现了便于输送氧气和运输根系中积累的乙醇、二氧化碳等物质的裂生性气腔[6]。本研究中,枳砧温州蜜柑根系也早于叶片(10 d)于淹渍处理5 d 时表现出异常,且多数生理指标此时也发生了显著变化,推测根系是受淹渍逆境直接伤害的器官,淹水后植株根系先发生了一系列形态结构变化和生理生化方面的适应性调节,以减缓伤害。
已有研究表明,水分胁迫下,果树叶片的叶绿素降解加剧[19],叶片衰老,光合性能降低[20]。叶绿素是植物在光合作用中吸收、传递和转换光能的载体[21],类胡萝卜素不仅能辅助植物的光吸收,还能作为抗氧化物清除光呼吸中产生的自由基[22]。本研究中,叶绿素含量的显著下降表明淹渍胁迫降低了枳砧温州蜜柑叶片对光能的同化能力,这与黄翼[5]在枳幼苗、刘超颖等[6]在枳砧脐橙幼苗上的研究结果一致。逆境胁迫条件下叶片净光合速率降低的原因可分为气孔和非气孔限制因素[5]。本研究中,淹渍胁迫下枳砧温州蜜柑叶片的净光合速率、气孔导度和蒸腾速率与同期对照相比显著降低,胞间CO2浓度显著升高,这与钟娟等[23]在金桔上的研究结果一致且符合Farquhar 等[24]提出的光合作用的非气孔限制判断标准。表明淹渍胁迫抑制了温州蜜柑的光合特性,而非气孔限制因素是导致本研究中的砧木温州蜜柑光合作用受抑制的主要因素。
3.2 枳砧温州蜜柑渗透调节物质和膜脂过氧化代谢对淹渍胁迫的响应
植物在淹渍逆境中启动的细胞膜脂过氧化系统可导致生物膜严重损伤[25],为适应淹渍逆境,植物会通过调节相应的代谢机制从而缓解伤害,渗透调节就是其中非常重要的调节机制之一[26]。前人的研究表明,植物在逆境条件下会通过调节体内的有机渗透物质如脯氨酸、可溶性糖和可溶性蛋白等以维持细胞的渗透势和水分平衡[27-28]。根系生理是评价抗逆性能的有效指标[29],脐橙[6,8]淹渍胁迫后根系SP和SS 含量在淹渍早期就出现了变化。本研究结果表明,温州蜜柑叶片和根系的膜脂过氧化程度、渗透调节物质等多数植物逆境响应指标在叶片和根系中呈显著负相关的趋势,表明温州蜜柑地上、地下部响应淹渍胁迫的模式不同;此外,多数逆境响应指标在根系淹渍胁迫5 d 后即显著低于对照,而叶片胁迫10 d 后显著高于对照,淹渍胁迫持续5 d 左右枳砧温州蜜柑根系即发生伤害,但此时地上部分未表现出异常,而进一步的淹渍胁迫导致地上部呈现明显症状时,树体的淹渍伤害已加重。上述结果表明,温州蜜柑叶片和根系组织对淹渍胁迫的响应机制不一致。淹渍逆境造成柑橘叶片和根系组织的形态、生理和代谢发生不同程度的变化,柑橘根系先受到胁迫,通过改变生长形态以期在短期内适应水淹环境。
3.3 枳砧温州蜜柑根系无氧呼吸酶系统对淹渍胁迫的响应
根系是植物从土壤中吸收、运输水分及养分的重要器官,其活力的高低是衡量植物抗逆性的重要指标之一。随着淹水胁迫时间的延长,猕猴桃的根系活力逐渐下降,其下降程度受基因型的影响[29]。桃[30]、火龙果[31]在淹渍胁迫后根系活力均下降,本研究中的枳砧温州蜜柑在淹渍胁迫下根系的DHA活性随着淹渍时间的延长也呈显著下降趋势。灌溉、雨水过多或洪水等易造成植物根系缺氧,而低氧可诱导植物产生多种参与无氧呼吸的酶,其中ADH酶是无氧呼吸时糖酵解途径最后一步的关键酶[32],可将乙醛转化为乙醇,以降低对细胞自身的毒害作用,并维持能量平衡[7]。耐涝性强的中山杉[33]在长期水淹条件下其根系的ADH 酶活性一直保持较高水平,而本研究中砧木温州蜜柑根系的ADH 酶活性在淹渍胁迫下呈先升后降趋势,这与其他经济类果树如甜樱桃[32]、桃[34]的研究结果一致。砧木温州蜜柑根系的ADH 酶活性在淹渍2~5 d 显著升高,表明根系已开始响应淹渍缺氧逆境,在淹渍10 d 后,伴随着ADH 酶活性的迅速下降至较低水平,DHA 活性也显著下降,表明根系活力已经显著减弱,长期持续淹渍胁迫可能对树体已经造成了不可逆的伤害。
ADH 活性及叶片EⅬ作为耐涝性的重要生理指标,已广泛应用于观赏植物[35]、无花果[36]和乌桕[37]的耐涝性鉴定评价,本研究中的枳砧温州蜜柑根系ADH活性在胁迫2 d时显著升高,叶片EⅬ在淹渍10 d显著升高,其变化与淹渍胁迫后对应组织中多数抗逆指标和树体表观症状变化趋势密切相关,推测根系ADH 活性和叶片EⅬ可作为早期判断枳砧温州蜜柑根系和叶片受到淹渍伤害程度的参考指标。
3.4 淹渍胁迫对枳砧温州蜜柑树体矿质营养积累的影响
植物主要依靠根系吸收矿质元素,淹渍胁迫直接对根系造成伤害,减少根系对矿质元素的吸收量。研究表明,玉米[38]、棉花[39]在淹渍胁迫后N、P、K 含量显著降低。柑橘在淹渍胁迫后根系对养分的吸收和转运受到影响,淹渍胁迫会导致卡列里佐枳橙幼苗中的总N 含量和浓度逐渐降低,碳水化合物如淀粉和蔗糖从叶片向根系的转运显著减少,造成C/N失调[7]。也有研究表明,长期淹水会影响柑橘幼苗对Fe 的吸收和分布[10]。本研究中淹渍胁迫下枳砧温州蜜柑根系中的N 含量变化不显著,但P、K、Ca、Mg含量显著降低,可能与不同柑橘类型对矿质营养的吸收存在差异有关[40]。温州蜜柑茎、叶中的矿质养分含量均未受淹渍胁迫的显著影响,淹渍胁迫下柑橘不同组织器官中矿质营养的积累机制可能存在差异,尚需进一步验证。
本研究结果表明:枳砧温州蜜柑的地上部和地下部组织响应淹渍胁迫的敏感程度及响应存在显著差异。根系是直接受到淹渍胁迫伤害的器官,较叶片更早响应淹渍胁迫,根系ADH 活性变化与根系受伤害程度紧密相关,叶片EⅬ与受淹渍胁迫程度密切相关。
