Long non-coding RNA SNHG16 promotes human placenta-derived mesenchymal stem cell proliferation capacity through the PI3K/AKT pathway under hypoxia
2022-10-11XuDongFengJiaHangZhouJunYaoChenBingFengRuiTianHuJianWuQiaoLingPanJinFengYangJiongYuHongCuiCao
Xu-Dong Feng, Jia-Hang Zhou, Jun-Yao Chen, Bing Feng, Rui-Tian Hu, Jian Wu, Qiao-Ling Pan, Jin-Feng Yang, Jiong Yu, Hong-Cui Cao
Xu-Dong Feng, Jia-Hang Zhou, Jun-Yao Chen, Bing Feng, Jian Wu, Qiao-Ling Pan, Jin-Feng Yang,Jiong Yu, Hong-Cui Cao, State Key Laboratory for The Diagnosis and Treatment of Infectious Diseases, National Clinical Research Center for Infectious Diseases, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou 310003, Zhejiang Province,China
Rui-Tian Hu, Department of Chemistry, Duke University, Durham, NC 27708, United States
Jian Wu, Jinan Microecological Biomedicine Shandong Laboratory, Jinan 250117, Shandong Province, China
Hong-Cui Cao, Key Laboratory of Diagnosis and Treatment of Aging and Physic-chemical Injury Diseases of Zhejiang Province, The First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou 310003, Zhejiang Province, China
Abstract BACKGROUND The effect of hypoxia on mesenchymal stem cells (MSCs) is an emerging topic in MSC biology. Although long non-coding RNAs (lncRNAs) and messenger RNAs(mRNAs) are reported to play a critical role in regulating the biological characteristics of MSCs, their specific expression and co-expression profiles in human placenta-derived MSCs (hP-MSCs) under hypoxia and the underlying mechanisms of lncRNAs in hP-MSC biology are unknown.AIM To reveal the specific expression profiles of lncRNAs in hP-MSCs under hypoxia and initially explored the possible mechanism of lncRNAs on hP-MSC biology.METHODS Here, we used a multigas incubator (92.5% N2, 5% CO2, and 2.5% O2) to mimic the hypoxia condition and observed that hypoxic culture significantly promoted the proliferation potential of hP-MSCs. RNA sequencing technology was applied to identify the exact expression profiles of lncRNAs and mRNAs under hypoxia.RESULTS We identified 289 differentially expressed lncRNAs and 240 differentially expressed mRNAs between the hypoxia and normoxia groups. Among them, the lncRNA SNHG16 was upregulated under hypoxia, which was also validated by reverse transcription-polymerase chain reaction.SNHG16 was confirmed to affect hP-MSC proliferation rates using a SNHG16 knockdown model.SNHG16 overexpression could significantly enhance the proliferation capacity of hP-MSCs,activate the PI3K/AKT pathway, and upregulate the expression of cell cycle-related proteins.CONCLUSION Our results revealed the specific expression characteristics of lncRNAs and mRNAs in hypoxiacultured hP-MSCs and that lncRNA SNHG16 can promote hP-MSC proliferation through the PI3K/AKT pathway.
Key Words: Human placenta-derived mesenchymal stem cell; Hypoxia; Long non-coding RNAs;Proliferation; Mesenchymal stem cell
INTRODUCTION
Recently, mesenchymal stem cells (MSCs) have gained much attention due to their therapeutic effects and potential applications in regenerative medicine[1]. MSCs have recently been shown to have therapeutic efficacy in various disease models and clinical diseases such as liver injury, coronavirus disease 2019, and Crohn’s disease[2-5]. MSCs have been reported to be present in bone marrow, placenta, umbilical cord, and adipose tissue and can be efficiently isolated[6-8]. However, the application of MSCs is limited due to the difficulty in obtaining the large numbers of MSCs required for clinical treatment (3 × 107cellsperinfusion)[9]. Tissue-derived primary MSCs occur in small numbers and requirein vitroexpansion before transplantation. Human placenta-derived MSCs (hP-MSCs) are a ubiquitous type of MSCs with lower immunogenicity and higher proliferative potential compared to bone marrow-derived MSCs, but these advantages may be compromised by inappropriate culture or changes in thein vitroenvironment.
To improve the proliferation potential of hP-MSCsin vitro, most researchers use different methods to stimulate the microenvironment of MSCsin vivo[10]. Among the proposed approaches to mimic the natural cellular microenvironment, hypoxia has garnered enormous interest. Hypoxia has been observed in different tissue niches, including the placenta (1%-5% O2) where hP-MSCs reside[11]. Since the oxygen concentration (almost 21%) in theex vivoculture system is much higher than the physiological oxygen concentration in the body, hypoxia could act as a physiological stimulus with a significant influence on cell fate. Numerous studies have reported that hypoxia can affect various biological properties of MSCs, such as proliferation capacity, multidirectional differentiation potential,migration, and apoptosis[12-14]. However, the underlying molecular mechanisms by which hypoxia regulates MSC biology remain unclear.
Long non-coding RNAs (lncRNAs) are RNAs longer than 200 nt with no protein-coding potential[15]. LncRNAs are the coordinators of the cellular biological regulatory network, participating in a variety of biological and pathological cellular processes such as cellular survival, proliferation, or migration through regulation of gene expression at transcriptional, post-transcriptional, or translational levels[16]. With advancements in gene sequencing technology, more and more lncRNAs related to cellular functions have been identified. However, the impact of hypoxia on the lncRNA expression profile of MSCs remains unclear. In addition, the roles of hypoxia-responsive lncRNAs remain to be explored. In this study, we investigated the effect of hypoxia on the proliferation potential of hP-MSCs and explored the role of lncRNAs in it.
MATERIALS AND METHODS
Cell culture
The protocols for hP-MSC isolation and hypoxic culture were as previously described[7]. All protocols for the processing of human tissues and cells were approved by the Ethics Committee of The First Affiliated Hospital of Zhejiang University (No. 2020-1088).
Colony-forming unit-fibroblast assay
For the colony-forming unit-fibroblast assay, 1000 hP-MSCs were plated on six-well plates in triplicate and cultured in complete medium for 14 d under normoxic or hypoxic conditions with medium changes every 3 d. Colonies were fixed with paraformaldehyde and then stained with crystal violet for enumeration.
Cell counting kit-8 assay
The corresponding cells were inoculated into 96-well cell culture plates at a density of 2000 cellsperwell. After 24, 48, 72 or 96-h culture, 10 μL of cell counting kit-8 reagent (Dojindo, Kumamoto, Japan)was added into each well to incubate for 2 h. The optical density value at 450 nm was measured using a microplate reader.
Flow cytometry analysis of cell cycle
The cells were collected with trypsin and fixed with cold 70% ethanol for 2 h. Fixed cells were then treated with propidium iodide staining solution (Beyotime, Nanjing, China). The cells were finally analyzed by flow cytometry. The proportions of cell population in G0/G1, S, and G2/M phases of the cell cycle were fitted and calculated using ModFit software.
Quantitative real-time polymerase chain reaction
Total RNA of cells was obtained using Trizol reagent (Invitrogen, Carlsbad, CA, United States)following the manufacturer’s protocol; the concentration of total RNA was quantified using a NanoDrop-2000 (Thermo Fisher Scientific, Waltham, MA, United States). cDNA was synthesized by reverse transcription reaction using a commercial lncRNA quantitative reverse transcription polymerase chain reaction (PCR) Starter Kit (RiboBio, Guangzhou, China). The final relative expression levels of genes were analyzed through the 2-ΔΔCtmethod usingGAPDHas the internal control. Primers were as follows:GAPDH: (forward) 5’-ACAACTTTGGTATCGTGGAAGG-3’, (reverse) 5’-GCCATCACGCCACAGTTTC-3’;SNHG16: (forward) 5’-GTTGCCACCCACAACCATT-3’, and (reverse) 5’-GCGGAGACACCAGGAGAACT-3’.
Western blot assay
The cellular protein was harvested using RIPA lysis buffer supplemented with protease and phosphatase inhibitor cocktail (Beyotime). The protein concentrations were detected using a BCA kit(Beyotime). The western blot was conducted as previously described[7]. The primary antibodies were anti-β-actin (Abcam, Cambridge, United Kingdom), anti-GAPDH (Abcam), anti-hypoxia-inducible factor 1α (HIF-1α) (Cell Signaling Technology, Danvers, MA, United States), anti-c-MYC (Abcam), antiproliferating cell nuclear antigen (Abcam), anti-CDK2 (Abcam), anti-CDK4 (Abcam), anti-CDK6(Abcam), anti-CyclinD1 (Abcam), anti-CyclinE1 (Abcam), anti-AKT (Abcam), and anti-phospho-AKT(Abcam).
RNA sequencing
Whole-transcriptome sequencing was quantitatively analyzed by Oebiotech (Shanghai, China). The libraries [including lncRNA and messenger RNA (mRNA)] were generated using TruSeq Stranded Total RNA with Ribo-Zero Gold (Illumina, San Diego, CA, United States) according to the manual. RNA was then sequenced on a HiSeq 2500 instrument (Illumina). Differential expression analysis of lncRNA and mRNA between the hypoxic and normoxic groups was conducted using the DESeq software package.The differentially expressed genes were identified with the criteria of fold change > 1.5 andP< 0.05. The Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were performed for differentially expressed mRNAs to explore their biological functions.Association analysis between lncRNAs and transcription factors and lncRNA-mRNA co-expression analysis were conducted to investigate lncRNA functions in cell biology.
Cell transfection
Lentivirus-mediated short hairpin RNA for silencingSNHG16in cells and recombinant lentivirus forSNHG16overexpression were constructed by Genomeditech (Shanghai, China). Transfection was performed following the manufacturer's instructions. Lentiviruses were added to infect cells at a multiplicity of infection of 50:1.
Statistical analysis
All data were expressed as the mean ± SD. Statistical evaluation of two groups was conducted using Student’sttest; aPvalue < 0.05 was considered to indicate statistical significance.
RESULTS
Hypoxic culture facilitated hP-MSC proliferation
HIF-1α is a critical regulator of cellular adaptation to the hypoxic microenvironment. When the expression of HIF-1α protein under hypoxia was assessed by western blot, hypoxia induced the expression of HIF-1α in hP-MSCs and stabilized its expression level during cell growth (Figure 1A). In addition, hP-MSCs cultured under hypoxia appeared to be relatively small, with a spindle-shaped morphology (Figure 1B). The cell counting kit-8 assay showed that hP-MSCs had higher proliferation potential (P< 0.0001) when they were maintained under hypoxia (Figure 1C). Similarly, the colonyforming unit-fibroblast assay indicated that hypoxia enhanced the hP-MSC proliferation rate. Although the difference in the number of hP-MSC colonies between the hypoxia and normoxia groups was not significant (P =0.249), the colony size of the hypoxia group was larger with darker staining, indicating a higher number of cells (Figure 1D). c-MYC and proliferating cell nuclear antigen are molecules closely related to cell proliferation and can be adopted to determine the status of cell proliferation. As expected,hypoxia significantly increased the expression of c-MYC and proliferating cell nuclear antigen,indicating that cells proliferated more rapidly under hypoxia (Figure 1E).
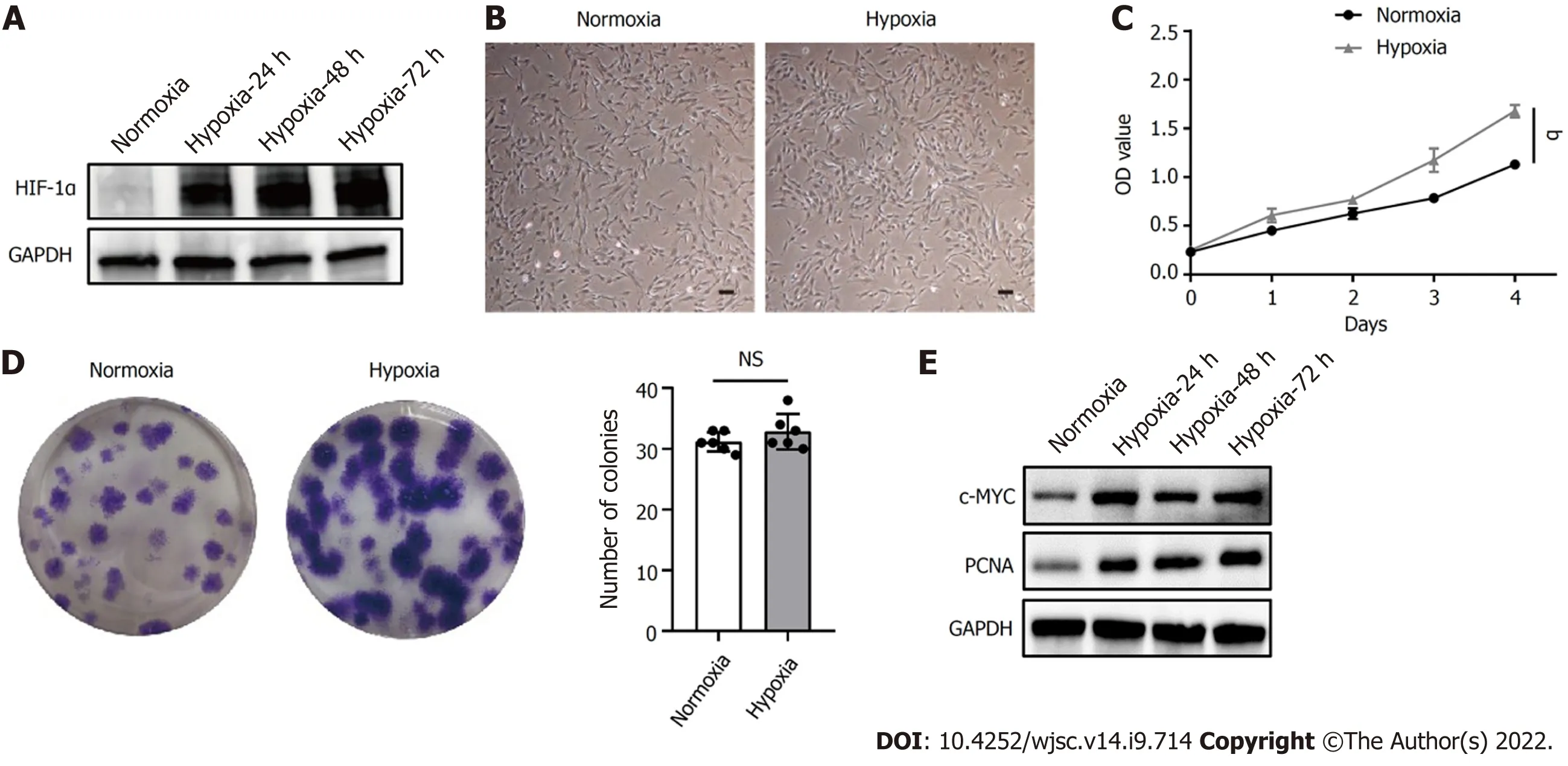
Figure 1 Hypoxia facilitated human placenta-derived mesenchymal stem cell growth and proliferation. A: Western blot analysis of hypoxiainducible factor 1α expression in human placenta-derived mesenchymal stem cells (hP-MSCs) under hypoxic culture for 24, 48, or 72 h; B: Morphology of the cultured hP-MSCs under hypoxia (scale bars, 100 μm); C: Proliferation curves of hP-MSCs were established based on cumulative cell numbers at different incubation times(0, 1, 2, 3, and 4 d) under normoxia or hypoxia; D: Colony size and colony number of hP-MSCs under normoxic or hypoxic culture (n = 6); E: The protein expression of c-MYC and proliferating cell nuclear antigen in hP-MSCs under hypoxic culture for 24, 48, or 72 h. Data are presented as means ± SD. aP < 0.05. NS: No significance; PCNA: Proliferating cell nuclear antigen; HIF-1α: Hypoxia-inducible factor 1α.
Hypoxia specifically altered the lncRNA and mRNA expression profiles of hP-MSCs
To further investigate the influence of hypoxia on hP-MSCs, whole-transcriptome sequencing was performed. First, six high-throughput sequenced transcriptomes were generated, containing over 650 million clean reads, among which three were from the normoxic group and three were from the hypoxic group. More than 96% of the raw reads were high-quality clean reads (Table 1). Ultimately, 10387 putative lncRNAs and 16041 mRNAs were identified. We further identified 289 differentially expressed lncRNAs (135 upregulated and 154 downregulated) and 240 differentially expressed mRNAs (156 upregulated and 84 downregulated) in the hypoxia group compared to normoxia group (Figure 2).Heatmap analysis clearly distinguished the hP-MSCs cultured under hypoxia from those cultured under normoxia. The top 20 differentially expressed lncRNAs and mRNAs are summarized in Tables 2 and 3.
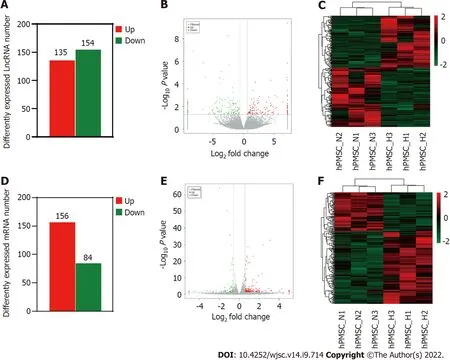
Figure 2 Long non-coding RNAs and messenger RNA expression profiles under hypoxia and normoxia. A: Number of differentially expressed long non-coding (lnc)RNAs between hypoxia and normoxia; B: Volcano plot depicting differentially expressed lncRNAs between hypoxia and normoxia; C: Heatmap of all differentially expressed lncRNAs identified in hypoxia vs normoxia; D: Number of differentially expressed messenger (m)RNAs; E: Volcano plot of differentially expressed mRNAs; F: Heatmap showing hierarchical clustering of differentially expressed mRNAs.
Differentially expressed mRNAs participated in cell proliferation function
The top ten enriched GO terms in biological process, molecular function, and cellular component were determined. For biological process, the differentially expressed mRNAs were related to regulation of cell growth (GO: 0001558), positive regulation of MAP kinase activity (GO: 0043406), and response to hypoxia (GO: 0001666) (Figure 3A). KEGG pathway enrichment analysis revealed several significantly enriched pathways, such as the HIF-1 signaling pathway (KEGG: hsa04066), Jak-STAT signaling pathway (KEGG: hsa04630), and Rap1 signaling pathway (KEGG: has04015) (Figure 3B). We further performed GO and KEGG analyses on the upregulated and downregulated genes separately.Upregulated genes were involved in regulation of cell growth (GO: 0001558) and regulation of cell proliferation (GO: 0042127) (Figure 3C) and were enriched in the HIF-1 signaling pathway (KEGG:hsa04066), Jak-STAT signaling pathway (KEGG: hsa04630), and AMPK signaling pathway (KEGG:hsa04152) (Figure 3D). Thus, hypoxia mainly affected cell functions, such as proliferation, by upregulating the expression of certain genes through several signaling pathways.
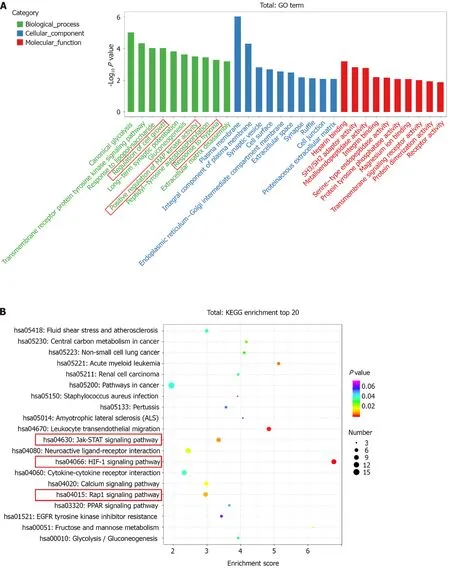
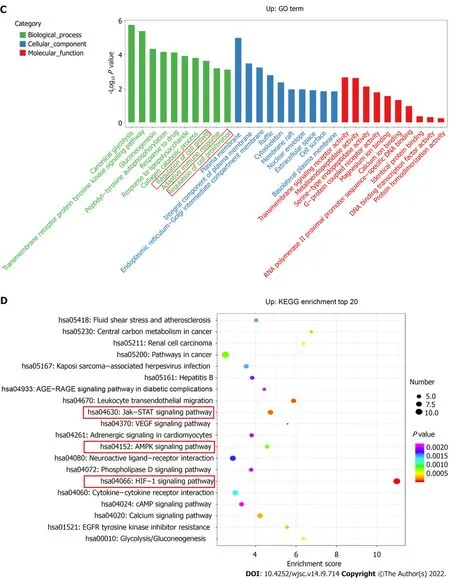
Figure 3 Gene Ontology terms and Kyoto Encyclopedia of Genes and Genomes pathway analyses. A: Enrichment of biological process, cellular component, and molecular function in all differentially expressed messenger RNAs (mRNAs); B: Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment of all differentially expressed mRNAs; the top 20 are listed; C: Gene Ontology (GO) annotation and functional enrichment of upregulated mRNAs; D:KEGG pathway enrichment analysis of upregulated mRNAs.
SNHG16 was a potential promotor of hP-MSC proliferation ability
Hypoxia affected cell proliferation by regulating cell cycle progression as the percentages of S (P= 0.011)and G2/M phase cells (P= 0.014) were larger under the hypoxic condition (Figure 4A). At the same time, the PI3K/AKT pathway was activated under hypoxia (Figure 4B). The PI3K/AKT pathway is responsible for coordinating a diverse range of cell functions, including proliferation and survival.These findings suggest that hypoxia can activate the PI3K/AKT pathway and modulate the cell cycle.To explore whether there are specific hypoxia-responsive lncRNAs that play a role in hypoxia-promoted cell proliferation, association analysis between lncRNAs and transcription factors and lncRNA-mRNA co-expression analysis were performed.SNHG16was related to the expression ofPIK3R5, a gene encoding the regulatory subunit of the PI3K gamma complex (Figure 4C and D).SNHG16was also correlated withFOSB, a key transcription factor in the cell cycle (Figure 4E). Quantitative reverse transcription PCR analysis confirmed that hypoxia induced the expression ofSNHG16(P= 0.003),consistent with the results of RNA sequencing (Figure 4F). Thus,SNHG16is a potential promoter of hPMSC proliferation under hypoxia.
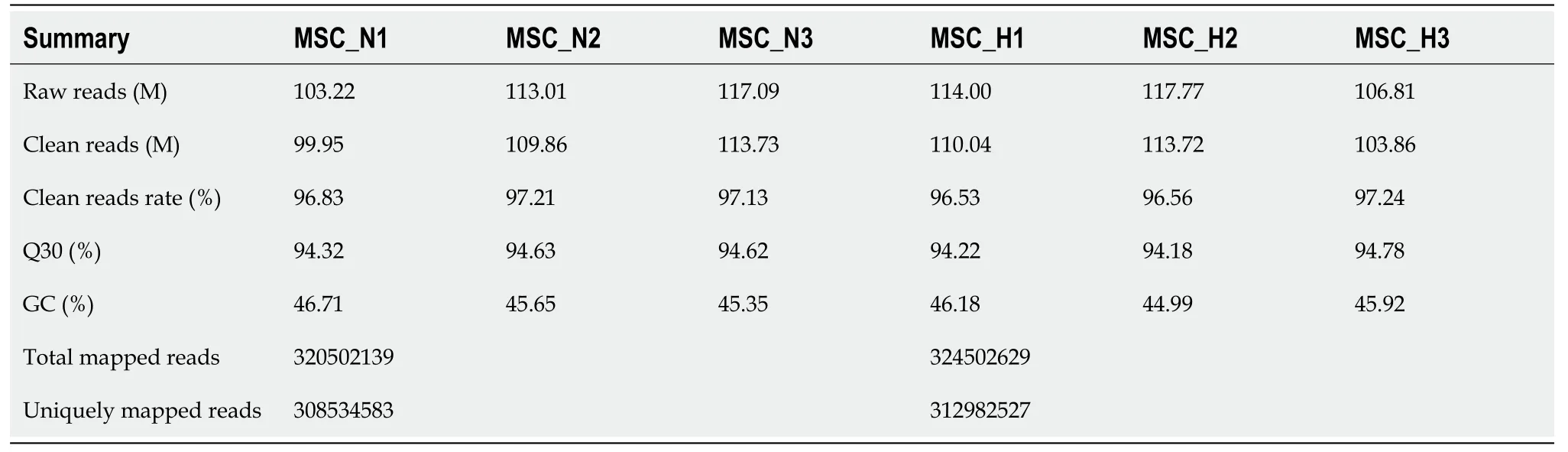
Table 1 Summary of the RNA sequencing data
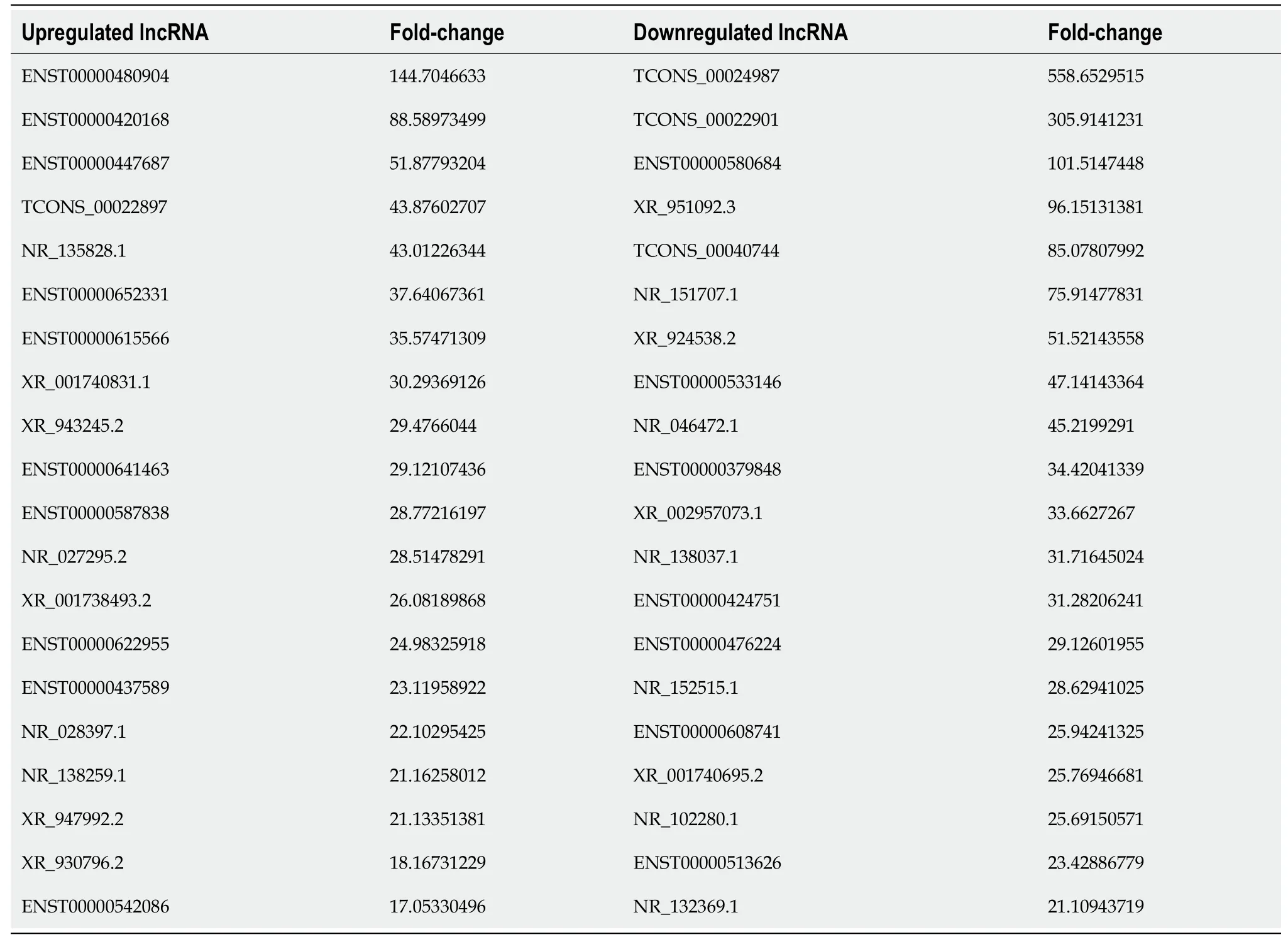
Table 2 Summary of the top 20 differentially expressed long non-coding RNAs
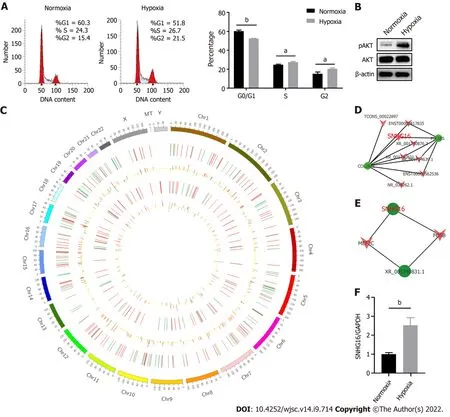
Figure 4 SNHG16 was a potential promotor of human placenta-derived mesenchymal stem cell proliferation ability. A: Cell cycle analysis of human placenta-derived mesenchymal stem cells (hP-MSCs) under hypoxic culture via flow cytometry; B: Western blot analysis of AKT phosphorylation in hP-MSCs exposed to hypoxia; C: Circos plot of the long non-coding RNAs (lncRNAs)-messenger (m)RNA co-expression network. The outermost circle is the autosomal distribution. The second and third circles are the distribution of differentially expressed lncRNAs on chromosomes. The red line represents upregulation, and the green line represents downregulation. Higher bars indicate a greater number of differential genes in the interval. The fourth and fifth circles are the distribution of differentially expressed genes on chromosomes, with the same interpretation as lncRNA; D: Part of lncRNA-mRNA interaction network analysis visualized using the Cytoscape software; E: Part of the association analysis of differentially expressed lncRNAs and transcription factors; F: Effects of hypoxia on the expression of SNHG16 in hP-MSCs by quantitative reverse transcription polymerase chain reaction. Data are presented as means ± standard deviation. bP < 0.01.
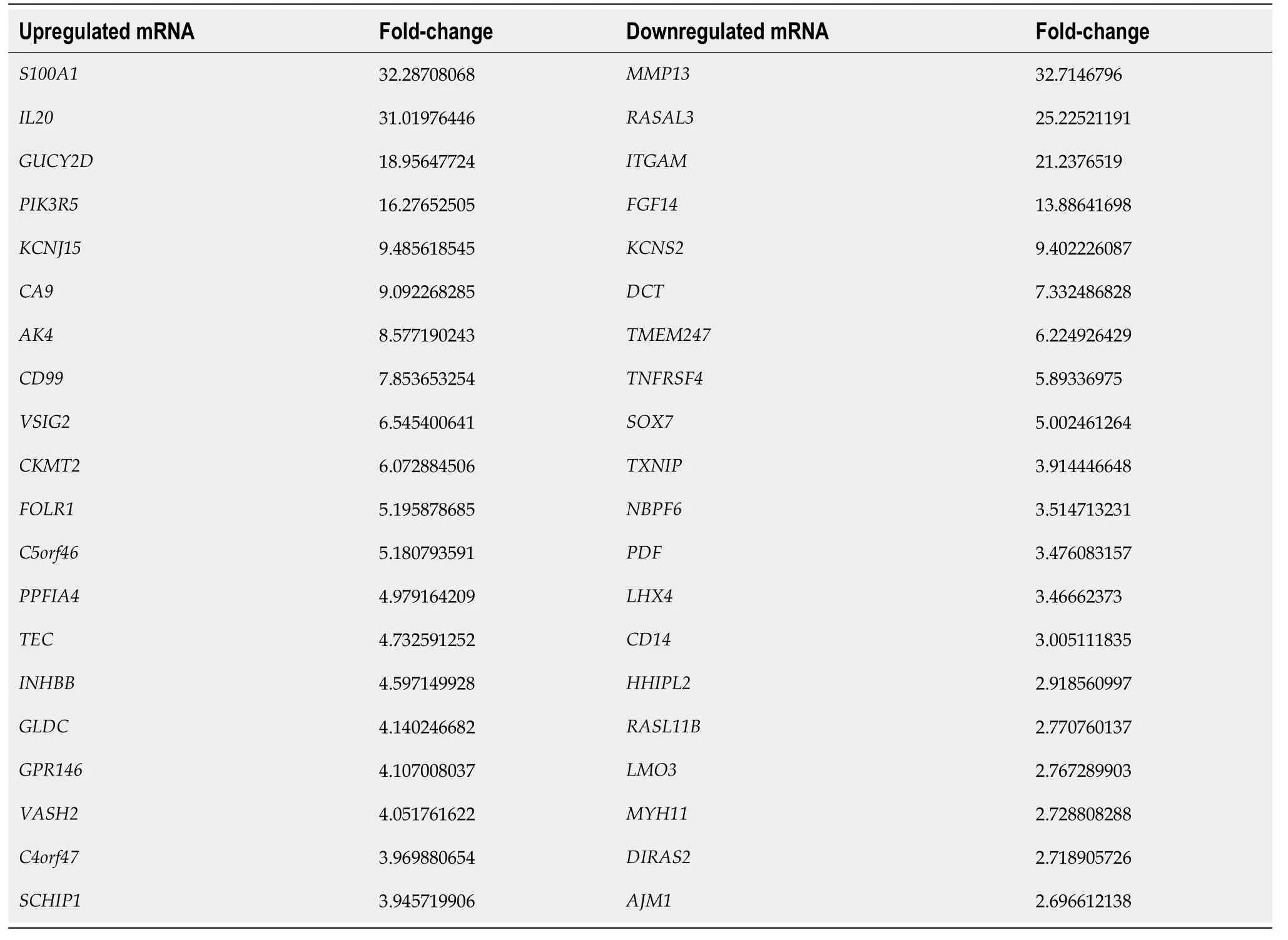
Table 3 Summary of the top 20 differentially expressed messenger RNAs
SNHG16 promoted proliferation of hP-MSCs via the PI3K/AKT pathway
To further confirm the biological function ofSNHG16in hP-MSCs, short hairpin RNA was used to specifically knock downSNHG16, whereas lentivirus overexpressingSNHG16was used to increaseSNHG16expression. By transfectingSNHG16short hairpin RNA, we found that sh-SNHG16significantly downregulatedSNHG16expression (P< 0.0001) by up to 80% (Figure 5A). The cell counting kit-8 assay then revealed that depletion ofSNHG16could attenuate the proliferation ability of hP-MSCs under both normoxia (P= 0.0003) and hypoxia (P= 0.0007) (Figure 5B and Supplementary Figure 1).Moreover,SNHG16knockdown decreased the cell numbers in S phase (P= 0.022) and increased the ratio of cells in the G0/G1 phase (P= 0.003) (Figure 5C). Furthermore, western blot showed that knockdown ofSNHG16downregulated the phosphorylation of AKT and the expression of several important cell cycle regulators, including CDK2, CDK4, CDK6, cyclin E1, and cyclin D1 (Figure 5D).Subsequently, we evaluated the effect ofSNHG16overexpression on cell proliferation and cell cycle transition. Quantitative reverse transcription PCR indicated thatSNHG16was upregulated approximately 8-fold (P= 0.0001) when transfected with lentivirus overexpressingSNHG16(Figure 6A).Overexpression ofSNHG16greatly enhanced the proliferative rate of hP-MSCs (P< 0.0001) and caused a mild increase in the ratio of cells in the S (P= 0.027) and G2/M phases (P= 0.003) (Figure 6B and C).The expression of G1 to S phase transition-related genes in theSNHG16overexpression group increased along with the activation of the PI3K/AKT pathway (Figure 6D). Overall, these data demonstrated thatSNHG16could facilitate the growth and cell cycle transition of hP-MSCs through activating the PI3K/AKT pathway.
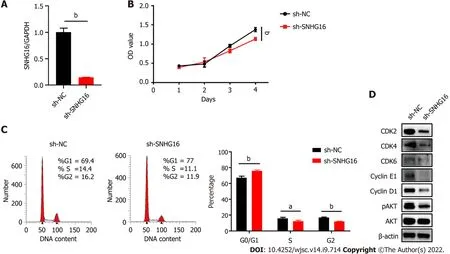
Figure 5 Knockdown of SNHG16 attenuated the proliferation ability of human placenta-derived mesenchymal stem cells. A: Quantitative reverse transcription polymerase chain reaction analysis of relative SNHG16 expression after transfection of SNHG16 short hairpin RNA (sh-SNHG16) and the corresponding controls (sh-NC) in human placenta-derived mesenchymal stem cells; B: Cell proliferation capacity evaluated by cell counting kit-8 assay; C: Cell cycle measured by flow cytometry; D: The G1 to S phase transition-related proteins and p-AKT detected by western blot analysis. Data are presented as means ± standard deviation. bP < 0.01.
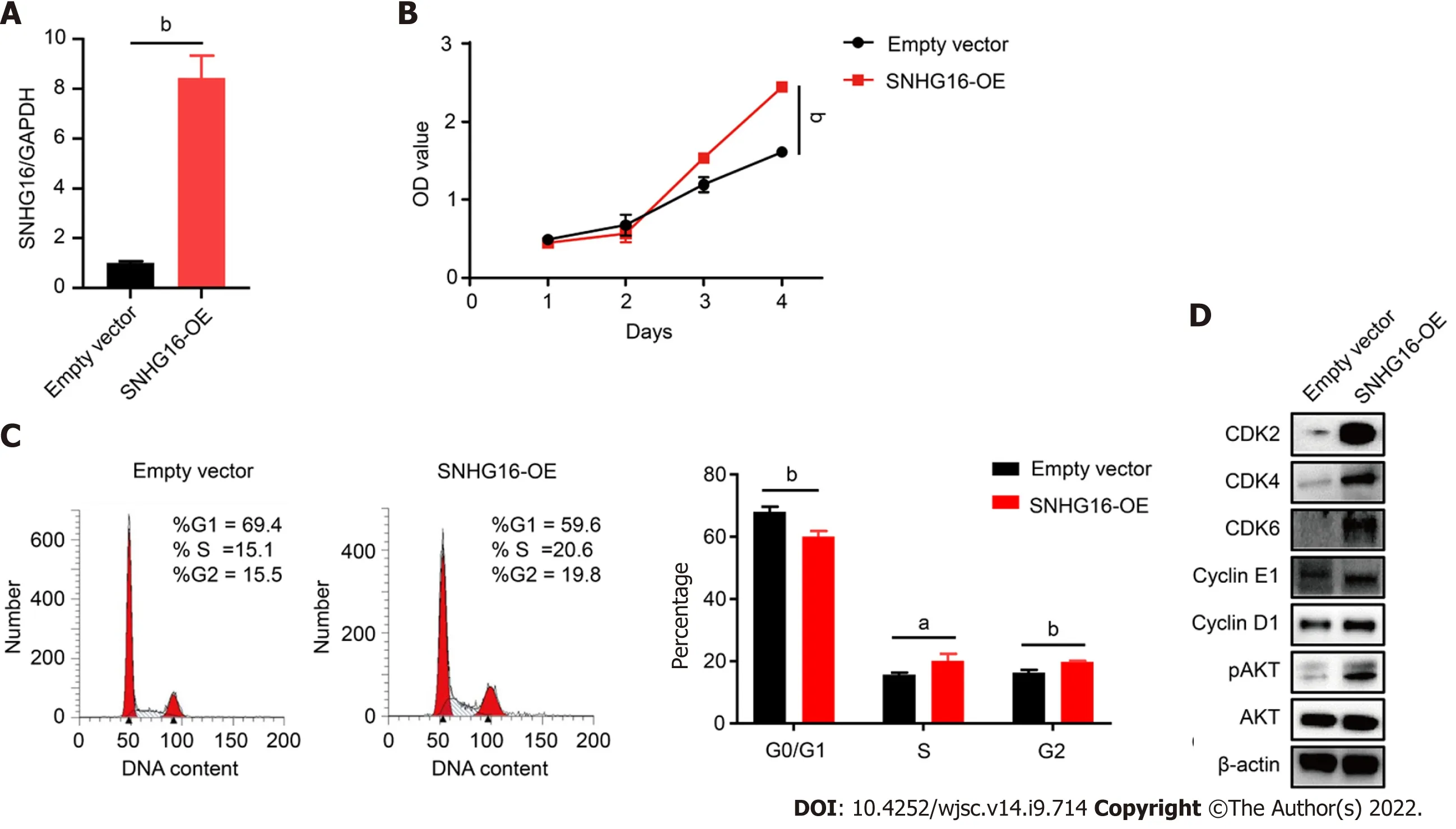
Figure 6 SNHG16 overexpression resulted in activation of the PI3K/AKT pathway and a significant enhancement in the proliferative rate of human placenta-derived mesenchymal stem cells. A: Quantitative reverse transcription polymerase chain reaction analysis of relative SNHG16 expression after transfection of lentivirus overexpressing SNHG16 (SNHG16-OE) and the corresponding empty vector in human placenta-derived mesenchymal stem cells; B: Cell proliferation after SNHG16 overexpression was evaluated by cell counting kit-8 assay; C: Cell cycle distribution after SNHG16 overexpression was evaluated by flow cytometry; D: The expression levels of CDK2, CDK4, CDK6, cyclin E1, cyclin D1, and phosphorylated AKT. Data are presented as the means ± SD obtained from three separate experiments. bP < 0.01.
DISCUSSION
MSCs have great potential to cure a variety of diseases, as evidenced by the rapid growth in the number of published preclinical and clinical studies. However, MSCs are found in very small numbers in most adult tissues, such as bone marrow, placenta, adipose tissue, umbilical cord, amniotic fluid, and muscle[17,18]. To generate sufficient clinical therapeutic quantities,in vitroexpansion is necessary[19].Managing and modifying culture conditions during amplification of MSCsin vitrois critical for the manufacture of effective cell therapies, as thesein vitroculture conditions affect the cell properties and cell behaviors after transplantation[20].
MSCs are widely located in the hypoxic microenvironment[21,22]. This physiological oxygen concentration is significantly lower than normoxic conditions typically used for MSC culture in the laboratory.Therefore, the application of physiological oxygen tension in stem cell research has attracted attention.Culturing MSCs under hypoxia has been consistently associated with increased cell proliferative rate,increased clonogenicity, decreased spontaneous differentiation, transcriptional alterations, and other cellular behaviors[11,23-25].
In the current study, we focused on the influence of hypoxia on hP-MSC proliferation ability. We found that hypoxic culture could facilitate hP-MSC proliferation, but enhanced clonogenicity under hypoxia was not observed in hP-MSCs. This finding provides a basis for exploring the underlying mechanism of the increased proliferation of hP-MSCs under hypoxic conditions.
Previous findings suggested that lncRNAs could exert regulatory function in MSC proliferation or differentiation. For example, Menget al[26] revealed thatlincRNA-p21promotes the migration and survival capabilities of mouse bone marrow-derived MSCsviathe HIF-1α/CXCR4 and CXCR7 axis under hypoxia[26]. In addition,LINC01119negatively regulates osteogenic differentiation of human bone marrow-derived MSCs, while the lncRNAsLOC100126784andPOM121L9Pimprove the osteogenic differentiation of human bone marrow-derived MSCs[27,28]. However, the role of lncRNAs in hP-MSCs has rarely been reported.
Here, we employed RNA sequencing technology to obtain a comprehensive and systematic understanding of lncRNAs in hP-MSCs under hypoxia condition. A total of 289 lncRNAs (135 upregulated and 154 downregulated) and 240 mRNAs (156 upregulated and 84 downregulated) were differentially expressed between the hypoxia and normoxia groups. Expression profiles of these differentially expressed genes were clustered hierarchically. GO and KEGG analyses suggested that the most enriched genes were positioned in the plasma membrane and related to regulation of cell growth and HIF-1 signaling pathway.
The results of the bioinformatic analysis were consistent with our experimental results. Combined with the individual analysis of upregulated genes, we found that hypoxia affected multiple cellular functions, mainly through upregulating the expression of certain genes. Moreover, hypoxia could mediate cell cycle progression and activate the PI3K/AKT pathway. Similarly, lncRNA-mRNA coexpression network analysis indicated thatSNHG16, a hypoxia-responsive lncRNA, is associated with key genes in the cell cycle or PI3K/AKT pathway. Therefore,SNHG16was selected as a potential promoter of the hP-MSC proliferative rate under hypoxia.
SNHG16is a member of the SNHGs and is well-documented for its oncogenic properties in numerous types of malignancies[29].SNHG16is reported to be involved in multiple cell biological functions,including cell cycle progression, proliferation, and migration[30-32]. In our study, we found that hypoxic culture could induce the expression ofSNHG16in hP-MSCs. We further verified thatSNHG16could promote cell cycle progression and cell proliferation of hP-MSCs by using knockdown and overexpression models. Moreover, we demonstrated that overexpression ofSNHG16could increase the phosphorylation of AKT with a simultaneous elevation in the expression levels of G1 to S phase transition related proteins, including CDK2, CDK4, CDK6, cyclin E1, and cyclin D1.
However, howSNHG16becomes integrated in the PI3K/AKT signaling pathway in the study remains unknown. There are some related articles on the mechanism by whichSNHG16regulates the AKT pathway in other models. For example,SNHG16could activate the PI3K/AKT pathway throughSNHG16/miR-338-3p/PLK4axis in cisplatin-resistant neuroblastoma cells[33]. Moreover,SNHG16was found to facilitate proliferative diabetes-related abnormalities in cell proliferation through regulatingmiR-7-5p/IRS1to activate PI3K/AKT pathway in HG-stimulated hRMECs[31]. It can be seen thatSNHG16mainly acts as a competing endogenous RNA to participate in the regulation of the PI3K/AKT signaling pathway. Our follow-up studies will take this as a starting point to elucidate the detailed mechanism ofSNHG16regulation of the PI3K/AKT pathway.
CONCLUSION
In this study, we have shown that hypoxia enhanced hP-MSCs proliferation ability and could specifically alter the lncRNA and mRNA expression profile. Furthermore, we identified a hypoxiaresponsive lncRNA,SNHG16, which may serve as a regulator of promoting hP-MSCs proliferation under hypoxia. Mechanically,SNHG16was shown to activate the PI3K/AKT signaling pathway and upregulate the expression of key cell cycle regulators to induce cell cycle transition.
ARTICLE HIGHLIGHTS
Research background
As the role of hypoxia on mesenchymal stem cells (MSCs) is an emerging topic of MSCs biology,increasing studies are devoted to researching the regulation mechanisms of hypoxia on the biological functions of MSCs. Long non-coding RNAs (lncRNAs) and messenger RNAs (mRNAs) are reported to possess a critical role in regulating MSC biological characteristics. Nonetheless, the specific expression and co-expressed profiles of lncRNAs and mRNAs in human placenta-derived MSCs (hP-MSCs) under hypoxia and underlying mechanism of lncRNAs on hP-MSCs biology are still unknown.
Research motivation
Although some studies have explored the effects of hypoxia on MSCs, the role of lncRNAs in them remains unclear.
Research objectives
In this study, we aimed to reveal the specific expression profiles of lncRNAs in hP-MSCs under hypoxia and initially explored the possible mechanism of lncRNAs on hP-MSCs biology.
Research methods
Here, we used a multigas incubator (92.5% N2, 5%CO2 and 2.5% O2) to mimic a hypoxia condition and observed that hypoxic culture can significantly promote the proliferation potential of hP-MSCs. RNA sequencing technology was applied to identify the exact expression profiles of lncRNAs and mRNAs under hypoxia. After establishment of SNHG16-knockdown and SNHG16-overexpression hP-MSCs, the effect of SNHG16 on proliferation capacity of hP-MSCs was analyzed via cell counting kit-8 and cell cycle analysis. Finally, the underlying mechanism was analyzed by western blot.
Research results
We identified 289 differentially expressed lncRNAs and 240 differentially expressed mRNAs between hypoxia group and normoxia group. Among them, the lncRNASNHG16was upregulated under hypoxia, which was also validated by reverse transcription polymerase chain reaction.SNHG16was confirmed to affect hP-MSCs proliferation rates by studying theSNHG16knockdown model.SNHG16overexpression could significantly enhance proliferation capacity of hP-MSCs, activate PI3K/AKT pathway, and upregulate the expression of cell cycle-related proteins.
Research conclusions
Our results revealed the specific expression characteristics of lncRNAs and mRNAs in hypoxia-cultured hP-MSCs and identified that hypoxia-responsive lncRNASNHG16can promote hP-MSC proliferation through the PI3K/AKT pathway.
Research perspectives
This study may contribute to understanding the role of noncoding RNAs in MSC biology.
FOOTNOTES
Author contributions:Feng XD contributed to the study design, experiments, data collection, and manuscript writing;Zhou JH, Chen JY, Feng B, Hu RT, and Wu J contributed to data collection and analysis; Pan QL, Yang JF and Yu J contributed to revising the manuscript; Cao HC contributed to design of the study, revision of the manuscript, and funding acquisition; All authors have read and approved the final manuscript.
Supported byStem Cell and Translational Research from National Key Research and Development Program of China, No. 2020YFA0113003; and National Natural Science Foundation of China, No. 81971756.
Institutional review board statement:All protocols for the processing of human tissues and cells were approved by the Ethics Committee of The First Affiliated Hospital of Zhejiang University, No. 2020-1088.
Informed consent statement:Informed consent was waived.
Conflict-of-interest statement:All the authors report no relevant conflicts of interest for this article.
Data sharing statement:The data presented in this study are available within the article text and figures.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORCID number:Xu-Dong Feng 0000-0002-4103-7737; Jia-Hang Zhou 0000-0003-0623-517X; Qiao-Ling Pan 0000-0002-3771-8193; Jin-Feng Yang 0000-0001-6284-1700; Jiong Yu 0000-0002-1821-1178; Hong-Cui Cao 0000-0002-6604-6867.
S-Editor:Fan JR
L-Editor:Filipodia
P-Editor:Zhang YL
杂志排行

World Journal of Stem Cells的其它文章
- Strategies to improve the effect of mesenchymal stem cell therapy on inflammatory bowel disease
- Overexpression of GATA binding protein 4 and myocyte enhancer factor 2C induces differentiation of mesenchymal stem cells into cardiac-like cells
- Intercellular mitochondrial transfer as a means of revitalizing injured glomerular endothelial cells