半滑舌鳎细胞分裂周期蛋白42 基因克隆与结构预测
2022-10-10刘厚孚谭静胡秀彩管振国吕爱军通信作者孙敬锋
刘厚孚,谭静,胡秀彩,管振国,吕爱军,通信作者,孙敬锋
(1.天津农学院 水产学院 天津市水产生态及养殖重点实验室,天津 300392;2.天津鼎正新兴生物技术有限公司,天津 300383)
半滑舌鳎(Cynoglossus semilaevis)是我国主要的海水经济鱼类之一[1],具有活动范围小,个体大、生长快、营养等级低、食性温和等性状优点。近年来,随着养殖规模日益扩大,半滑舌鳎皮肤溃烂病、腹水症等病害问题也随之而来,对水产养殖行业造成了一定的经济损失。目前,对于半滑舌鳎天然免疫和细胞分子免疫机制知之甚少[2-3]。
细胞分裂周期蛋白42(Cell division cycle 42,Cdc42)属于小Rho GTPase 家族,与GTP 结合时处于活化状态,与GDP 结合时处于失活状态,是一种具有酶活性的Rho GTP 酶[4-5]。研究发现,Cdc42 参与多种细胞功能,包括调节细胞骨架、控制细胞极性等[6-7]。迄今,对人和哺乳动物研究表明,Cdc42基因与疾病和机体免疫应答密切相关,包括癌症、艾滋病等多种疾病发生,且参与免疫与炎症反应[8-9]。BURBAGE 等研究发现Cdc42是B 细胞分化的关键调控因子,是抗病毒体液免疫所必需的因子[10];HADDAD 报道Cdc42与WASP之间相互作用引导SDF-1 诱导T 淋巴细胞趋化[11];WESTERBERG 等研究表明,Cdc42、Rac1 和Wiskott-Aldrich 综合征蛋白参与B 淋巴细胞的细胞骨架调节[12];王缘等[13]在小鼠巨噬细胞中特异性敲除Cdc42基因,并使用LBS 诱导炎症反应,结果发现小鼠死亡率增高,证实Cdc42在免疫调节中发挥了重要作用。
在水产动物中,针对Cdc42基因克隆及表达分析的研究鲜有报道[5,14]。关于半滑舌鳎Cdc42的功能研究尚未见相关文献报道。因此,本研究以半滑舌鳎为研究对象,对其皮肤细胞分裂周期蛋白42 基因进行克隆,并对其基因编码序列进行蛋白结构预测分析,为今后深入研究Cdc42基因功能提供科学参考。
1 材料与方法
1.1 试验材料
半滑舌鳎(Cynoglossus semilaevis)取自天津市海发珍品实业发展有限公司,体重平均(110±10)g,取其皮肤组织立即投入液氮冷冻,后转入-80 ℃冰箱保存,用于总RNA 提取。RNA 提取试剂盒、逆转录试剂盒、PCR Master Mix、PCR Buffer 体系购自TaKaRa 公司,DNA Marker、UNIQ-10 柱式胶回收试剂盒购自上海捷瑞生物工程公司。
1.2 试验方法
1.2.1 引物设计
根据半滑舌鳎Cdc42基因编码序列(登录号:XM_008320479.3),用生物软件Primer5.0 设计1对特异性引物(表1),引物由苏州金唯智生物有限公司合成。
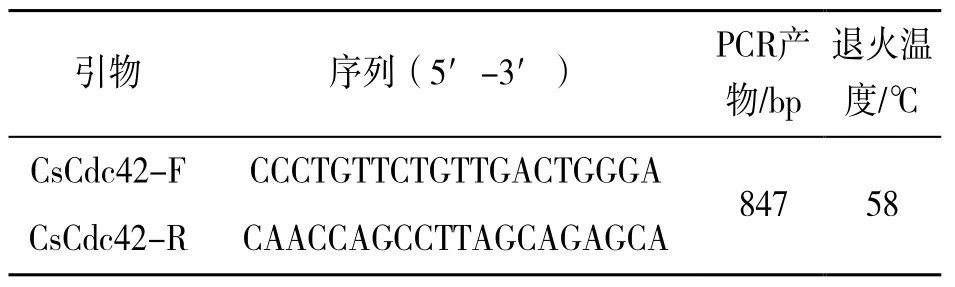
表1 Cdc42 的特异性引物
1.2.2 总RNA 提取和cDNA 合成
参照HUSAIN 等[15]的提取方法,使用TRIzol Reagent 试剂提取半滑舌鳎皮肤组织的总RNA,并分别加入氯仿、异丙醇和乙醇,离心、震荡、弃去上清液等。使用超微量分光光度计测量提取的总RNA 浓度,并通过琼脂糖凝胶电泳进行检测。以提取的总RNA 为模板,用TaKaRa 的反转录试剂盒(PrimeScriptTM II 1st Strand cDNA Synthesis Kit)合成cDNA。合成的cDNA 保存于-20 ℃备用。
1.2.3CsCdc42基因克隆
参照舒欢欢等[16]的克隆步骤进行试验,用25µL 体系扩增半滑舌鳎皮肤的Cdc42基因。反应体系具体为:Cdc42基因DNA 模板2 µL,上下游引物各1 µL,无菌超纯水11 µL,2×Taq PCR Mix 10µL。PCR 反应条件为:95 ℃预变性2 min;95 ℃40 s,55 ℃ 45 s,72 ℃ 1 min 30 s,30 个循环;72℃延伸5 min。使用1%琼脂糖PCR 检测,切胶回收,并与pMD18-T 载体连接、转化到感受态大肠杆菌DH5α 后,挑选阳性单克隆菌液送到天津金唯智生物工程有限公司测序。
1.2.4CsCdc42基因序列与结构预测
参照谭静等[17]的文献进行。采用ORF 程序(https://www.ncbi.nlm.nih.gov/orffinder/)查找序列的开放阅读框,利用在线分析软件EXPASY、PHYRE2、SMART 和PSIPRED 进行生物信息分析及结构预测,用DNAStar 软件进行基因蛋白抗原性、亲水性等理化特性分析,用ClustalW 及MEGA 7.0 进行序列比对及构建系统进化树分析。
2 结果
2.1 半滑舌鳎CsCdc42 基因克隆
以半滑舌鳎皮肤组织cDNA 为模板,采用RT-PCR 方法扩增Cdc42基因得到目的条带,并与预期片段大小一致(图1)。切胶回收、将其亚克隆至pMD18T 载体,转化大肠杆菌DH5α 后、挑选pMD18T-Cdc42 阳性克隆基因测序,测序获得半滑舌鳎Cdc42目的基因序列及其引物(图2),命名为CsCdc42。
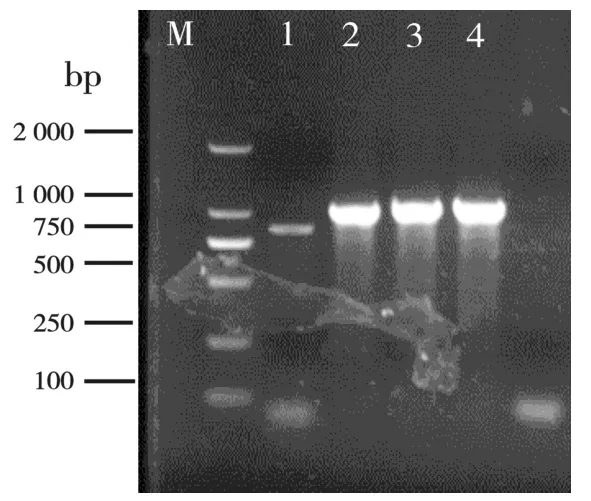
图1 半滑舌鳎CsCdc42 基因PCR产物琼脂糖凝胶电泳
图2 半滑舌鳎CsCdc42基因的引物序列
2.2 半滑舌鳎 Cdc42 cDNA 编码蛋白的序列特征及生物信息学分析
半滑舌鳎 CsCdc442 基因测序获得目的基因序列的片段长度为847 bp,推测编码191 个氨基酸(图3)。预测理论等电点(PI)为6.73,分子量为21.26 kD。通过生物信息学方法预测表明,Cdc42基因没有信号肽,推测 Cddc42 可能不是分泌蛋白。对半滑舌鳎CDc42蛋白跨膜区分析表明,该蛋白无跨膜区,说明该蛋白为非跨膜蛋白。亲疏水性分析结果显示疏水性最小值约为-3.01,最大值约为3.00,,整个多肽链中为负值的氨基酸占多数,推测CsCdc42 为亲水性蛋白(图4),蛋白平均系数GRAVY 为-0.195。

图3 半滑舌鳎CsCdC42 cDNA编码区序列及其推导的氨基酸序列
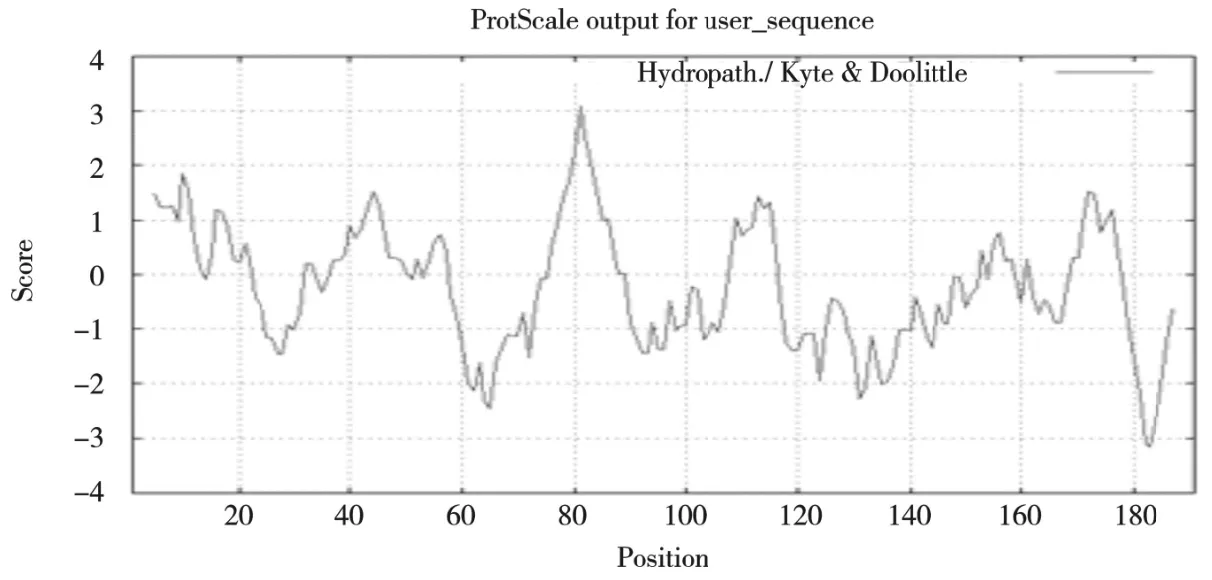
图4 半滑舌鳎CsCdc42 亲疏水性分析
预测CsCdc42 含有7 个主要抗原表位(27-28、61-63、90-92、120-122、131-133、164-166、181-183 aa),说明该基因蛋白抗原性良好(图5)。
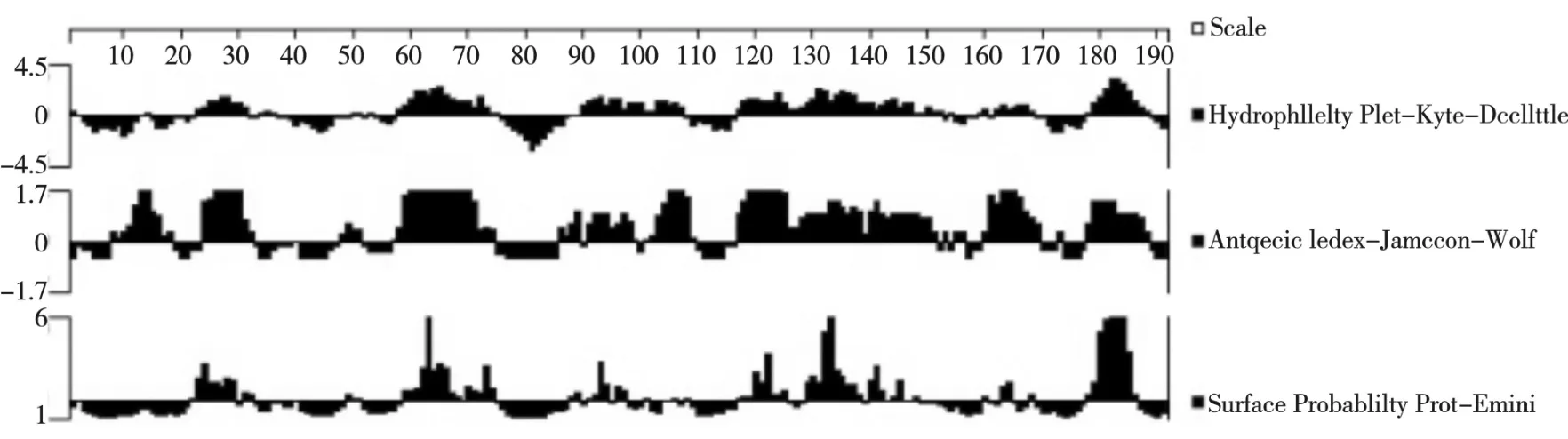
图5 半滑舌鳎CsCdc42 蛋白表面和抗原指数分析
进一步对CsCdc42 蛋白修饰位点预测发现CsCdc42 没有糖基化位点,有多个磷酸化位点,当阈值≥0.5 时共有17 个磷酸化位点,其中8 个丝氨酸(Ser)磷酸化位点(S22/30/70/82/84/89/90/126),6 个苏氨酸(Thr)磷酸化位点(T2/24/25/109/117/161),3 个酪氨酸(Tyr)磷酸化位点(Y30/64/157)。此外,阈值≤0.5 时,丝氨酸磷酸化位点主要分布在80-90 aa之间,苏氨酸磷酸化位点主要分布在20-40 aa 之间,酪氨酸磷酸化位点在20-40 aa 之间显著分布(图6)。
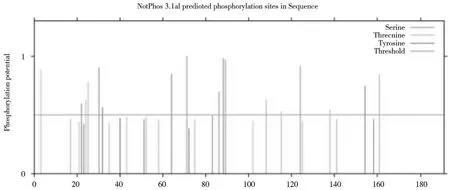
图6 半滑舌鳎Cdc42 基因的磷酸化位点分析
2.3 Cdc42 多序列比对与系统发育树分析
采用Blast 软件在线搜索结果显示,CsCdc42与斑马鱼(Danio rerio,NP_956926.1)的氨基酸同源性最高为99.43%,其次与人(Homo sapiens,pdb|2ODB|A)和青鳉(Oryzias melastigma,XP_024148790.1)分别为98.43%和97.91%,与虹鳟(Oncorhynchus mykiss,ACO08171.1)、大菱鲆(Scophthalmus maximus,AWP03434.1)、果蝇(Drosophila willistoni,XP_002075304.1)、凡纳滨对虾(Penaeus vannamei,AIY53010.1)、双斑蛸(Octopus bimaculoides,XP_014784384.1)的同源性分别为95.81%、94.76%、93.19 %、91.10%、91.10%。进行CsCdc42基因氨基酸多重序列对比分析,发现在人、斑马鱼、青鳉、果蝇等模式生物以及多种水产动物中,Cdc42 蛋白序列含有RHO 结构域、ATP/GTP 结合位点、酰基化位点3个高保守区域,这可能与细胞分裂周期蛋白功能有关(图7)。从NCBI 数据库下载人、斑马鱼、大菱鲆、青鳉、虹鳟等17 个Cdc42相关基因序列,构建系统发育树分析结果显示,CsCdc42与斑马鱼(Danio rerio,NP 956926.1)、人(Homo sapiens,pdb|2ODB|A)、大菱鲆(Scophthalmus maximus,KAF0035709.1)、青鳉(Oryzias latipes,XP 011475757.1)、虹鳟(Oncorhynchus mykiss,XP 021472057.1)亲缘关系最近,自然聚为一支;与南美白对虾(Penaeus vannamei,AIY53010.1)、果蝇(Drosophila melanogaster,AAD43792.1)亲缘关系较近;与南极鳕(Notothenia coriiceps,XP_010773360.1)、杂色鳉(Cyprinodon variegatus,XP_015226325.1)、斗鱼(Betta splendens,XP_029023734.1)、鲫(Carassius auratus,XP_026109495.1)等亲缘关系较远,聚为另一支(图8)。

图7 不同物种Cdc42 氨基酸序列多重比对分析
2.4 半滑舌鳎Cdc42 结构预测
对半滑舌鳎Cdc42 结构预测结果表明,Cdc42只有一个RHO 结构域,氨基酸位点在6-179 aa。二级结构预测α 螺旋占30.9%、β 折叠占24.6%、无规则卷曲占44.5%,说明Cdc42 主要由α 螺旋和无规则卷曲构成(图9A);由三级结构预测可以看出CsCdc42 是一个球形蛋白质(图9B)并且和二级结构结果基本一致,同时预测在氨基酸10-18 aa 为CsCdc42 的ATP/GTP 结合位点,在54-60 aa 为CsCdc42 的酰基化位点(图9C)。
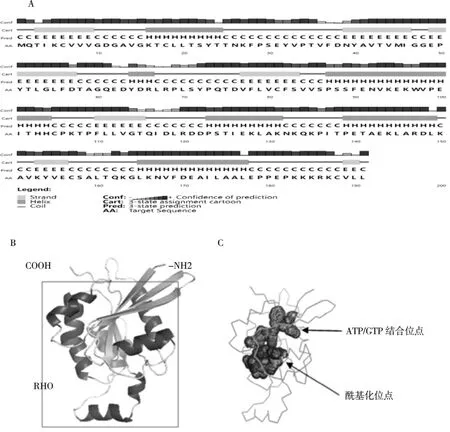
图9 Cdc42 基因编码的蛋白结构预测
3 讨论
迄今为止,对于鱼类的Cdc42基因研究相对较少,关于半滑舌鳎Cdc42功能研究尚未见相关文献报道[5-6]。胡默俨[14]对草鱼Cdc42基因克隆表达分析,结果表明Cdc42基因编码191 个氨基酸,为不稳定性蛋白,无跨膜结构域和信号肽,并只含有一个保守结构域RHO,这与本试验半滑舌鳎CsCdc42基因序列分析结果基本一致。进一步通过Cdc42氨基酸序列构建系统发育进化树分析,结果显示半滑舌鳎CsCdc42 属于Rho 超家族酶蛋白家族,而且与斑马鱼、人、青鳉和虹鳟Cdc42亲缘关系最近。此外,预测半滑舌鳎CsCdc42含有7 个主要抗原表位(27-28、61-63、90-92、120-122、131-133、164-166、181-183 aa),说明该基因蛋白抗原性良好,其抗原保护性功能值得进一步研究。
Cdc42 是Rho 蛋白家族中研究比较深入的成员,可以调节肌动蛋白细胞骨架和各种细胞黏附,对于机体发育与繁殖有重要影响[7]。在对人类和哺乳动物的研究中发现Cdc42与许多器官以及各种疾病密切相关。LI 等[18]通过将Cdc42-flox 小鼠与肌球蛋白轻链(MLC)2a-Cre 小鼠杂交试验,确定Cdc42在心肌细胞增殖和细胞黏附中有重要作用。OLENIK 等[19]通过采用原位杂交和Western blot方法研究Rho GTPases 在成年大鼠大脑中的表达情况,在海马神经元、齿状回颗粒细胞和肺门细胞中检测到大量RhoA、RhoB、Rac1 和Cdc42 mrna,Western blot 检测到海马、小脑、丘脑和大脑皮层中含有RhoA和Cdc42蛋白,所以认为Rho GTPases在大脑细胞功能调节中发挥作用。ZAN 等人通过逆转录聚合酶链反应检测20例食管癌及癌旁正常组织中Cdc42和Rb基因表达,结果发现Cdc42基因异常表达影响了食管癌发生[20]。CHERNICHENKO等[21]通过单个细胞轨迹分析对鸟嘌呤核苷酸交换因子(GEFs)进行siRNA 筛选,认为Cdc42在癌细胞神经侵袭及治疗癌症方面有重要意义。目前,对水产动物的Cdc42基因研究发现,Cdc42可能对于水产生物防御病原体的细胞以及分子机制有重要作用。李丽丽等[5]通过对斑节对虾Cdc42基因克隆表达分析,研究斑节对虾在哈维弧菌(Vibrio harveyi)胁迫下的表达情况,结果发现Cdc42在血淋巴中的表达较高,并且在哈维弧菌胁迫情况下发现Cdc42上调显著,证明Cdc42可能具有重要免疫调节作用;胡默俨[14]在草鱼Cdc42克隆试验中,通过荧光定量PCR 技术发现Cdc42基因在头肾、中肾、肝、脾等免疫器官中均有不同程度表达,但Cdc42在水产动物中的具体免疫机制仍不清楚,值得进一步研究。
本研究采用RT-PCR 方法对半滑舌鳎皮肤Cdc42基因进行克隆,并对其编码蛋白质进行序列和结构预测分析,为今后深入研究Cdc42基因功能打下基础。关于鱼类细胞分裂周期蛋白42 基因序列特征及其蛋白功能,今后仍有待进一步试验研究证实。
