珠江河口棘头梅童鱼时空分布以及影响因素的研究进展
2022-10-10熊朋莉徐姗楠陈作志蒋佩文范江涛
熊朋莉, 徐姗楠, 陈作志, 张 帅, 蒋佩文, 4, 范江涛
珠江河口棘头梅童鱼时空分布以及影响因素的研究进展
熊朋莉1, 3, 4, 徐姗楠2, 3, 陈作志2, 3, 张 帅2, 蒋佩文2, 3, 4, 范江涛2, 3
(1. 上海海洋大学海洋科学学院, 上海 201306; 2. 中国水产科学研究院南海水产研究所, 农业农村部外海渔业开发重点实验室, 广东 广州 510300; 3. 南方海洋科学与工程广东省实验室, 广东 广州 511458; 4. 上海海洋大学水产与生命学院, 上海 201306)
根据2019—2020年珠江河口底拖网调查数据, 结合实测环境数据和遥感环境数据, 建立以资源密度为响应变量的珠江河口棘头梅童鱼的广义加性模型(generalized additive models, GAM), 分析棘头梅童鱼()时空分布规律, 筛选影响其时空分布的关键环境因子。结果表明, 棘头梅童鱼主要分布在伶仃洋海域, 相对于冬、春两季, 秋季棘头梅童鱼向南移动。构建最佳GAM模型的变量组合为底层盐度、叶绿素浓度、底层酸碱度和季节, 该模型对资源密度的累计方差解释率为96.8%, 其中叶绿素浓度对资源密度的贡献率最高, 为58.2%, 季节、底层盐度、底层酸碱度对其资源密度的贡献率分别为21.1%、14.8%、2.7%。研究结果可为棘头梅童鱼资源保护提供有效的科学依据。
时空分布; 环境因子影响效应; GAM模型; 棘头梅童鱼; 珠江河口
棘头梅童鱼()是珠江河口生态系统重要的经济物种, 也是中华白海豚()重要的饵料生物, 被列入国家重点保护经济水生动物资源名录[1-2]。现有研究表明, 虽然棘头梅童鱼一年中多批次产卵, 可以承受较高的开发率, 但珠江河口棘头梅童鱼仍处于过度捕捞的状态, 种群出现性早熟现象[3-4]。探究珠江河口棘头梅童鱼的时空分布及其影响因素是阐明资源分布规律以及进行资源评估的基础, 研究结果可对资源可持续利用以及珠江河口生物多样性保护具有重要意义。
渔业资源时空分布与环境因素之间的关系十分复杂, 具有非线性和不可累加的特性[5]。广义加性模型能够较好的处理响应变量和多个解释变量之间的关系, 且该模型在构建过程中有较高的灵活性, 因此被广泛用于渔业资源分布和环境因子关系之间的定性分析[6-8]。在渔业科学研究中, 最佳GAM模型变量组合筛选方法主要分为逐步回归法和全子集回归法, 利用不同的方法筛选出的模型变量不同, 且每种方法都存在一定的缺点[9-10]。逐步回归法在模型评价全面性方面有限制, 全子集回归法虽然可以评价所有可能的模型, 但当模型中预测变量较多时, 其运行效率较低[11]。本研究根据2019—2020年春、秋、冬3个季节的珠江河口底拖网调查资料, 采用逐步回归法和全子集回归法, 筛选出棘头梅童鱼最佳的GAM模型变量组合。该方法保留对响应变量有较高贡献率的因子, 评价更多可能模型的同时, 提高了模型运算效率, 可为GAM模型最佳环境变量方法筛选提供参考。
1 材料与方法
1.1 数据来源
渔场数据来自于2019—2020年在珠江河口进行的底拖网资源调查(图1), 调查时间为冬季(2019年12月、2020年1月)、春季(2020年4月)、秋季(2020年9月)。调查船船长21.68 m, 型宽5.8 m, 型深3.3 m, 主机总功率为143.0 kW, 网具的上纲长度36 m, 囊网网目为3 cm, 网衣全长50 m。调查期间, 每站拖曳时间30 min, 拖速控制在3节左右。采集的渔获全部带回岸上进行种类鉴定并测定体长、体质量以及性腺成熟度等生物学数据。
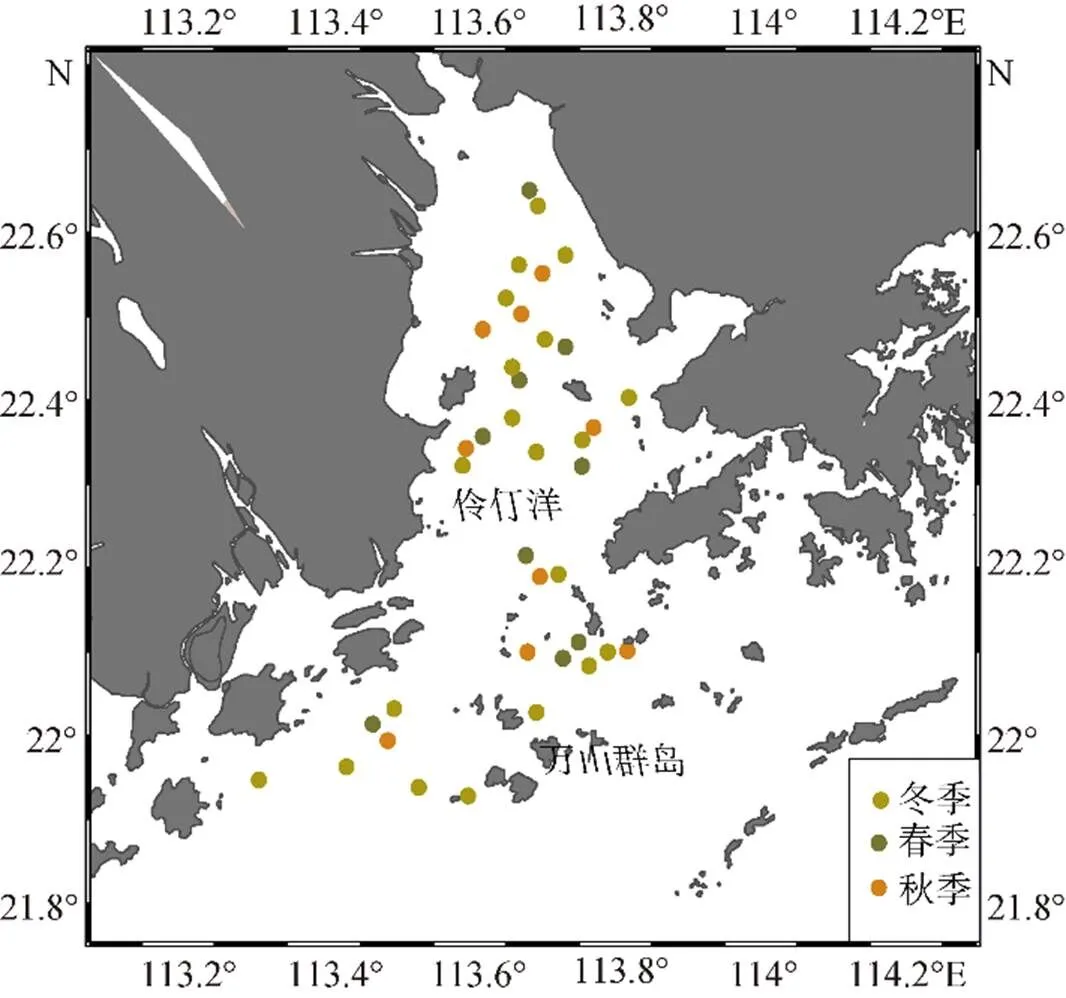
图1 珠江河口调查站位分布
环境数据包括现场测定的站位水深(Depth)、底层盐度(bottom salinity, BS)、底层温度(bottom temperature, BT)、底层溶解氧(bottom oxygen demand, BDO)、底层酸碱度(BpH)等, 叶绿素浓度(chlorophyll-concentration, CHL)数据来自美国国家海洋和大气管理局(https://coastwatch.pfeg.noaa.gov/), 时空因子包括经度(Longtitude, Long)、纬度(Latitude, Lati)、季节(Season)。
1.2 数据处理
采用扫海面积法估算资源密度, 计算公式为:
=/, (1)
式中,为每个站点的资源密度,为每个站点单位小时拖网的渔获量[kg/(网·h)],为网具每小时实际扫海面积[km/(网·h)],为可捕系数, 取0.5[12-13]。
偏度(skewness,S)和峰度(kurtosis,)是量测数据正态分布特性的两个指标, 变异系数(C)可以用来衡量数据变异程度的特征大小[14]。其计算分别为:

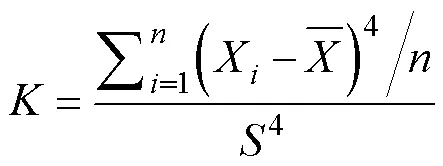
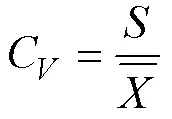
式中,表示标准差,X为平均值,为样本数量,表示第个数值。
1.3 广义加性模型
将Lon、Lat、Depth、BS、BT、BDO、BpH、CHL、Season等因子作为解释变量, 资源密度作为响应变量构建珠江河口棘头梅童鱼的GAM模型, 其中季节为分类离散变量, 其他环境变量为连续变量。
表达式为:
lg()=s(Long)+s(Lati)+s(Depth)+s(BS)+s(BT)+
s(DO)+s(BpH)+s(CHL)+Season+,(5)
式中,为棘头梅童鱼的资源密度, 对其取对数以满足GAM建模对响应变量须符合正态性的要求; s为自然样条平滑函数,为模型误差, 符合高斯分布。
1.4 最佳模型因子组合的选择
采用方差膨胀因子(VIF)对解释变量进行多重线性检验, 避免共线因子叠加作用于模型, 降低误删重要环境因子的概率[7]。当VIF阈值≥10时, 表示变量存在严重的多重共线性。当只有1个解释变量的VIF阈值≥10时, 去除该解释变量, 并对其他解释变量进行共线性检验; 当多个解释变量的VIF阈值≥10时, 把VIF阈值≥10的解释变量逐步加入其他解释变量进行再次共线性检验。逐一构建单个通过VIF检验的因子与资源密度的GAM模型, 并与初始资源密度GAM模型(Lg()=null)的AIC(赤池信息准则, Akaike Information Criterion)值作比较。经过AIC筛选出的因子作基于调整后的2(Adjust2, adj2)全子集回归, 将包含较大相对方差解释率的变量且adjR2最大的子集设为最佳GAM模型变量组合。
2 结果
2.1 珠江口棘头梅童鱼资源密度分析
各个季节棘头梅童鱼资源密度偏度均大于0, 数据分布整体呈右偏。冬季和秋季的资源密度频数成尖顶峰分布(>3), 春季资源密度频数成平顶峰分布(<0), 各季节渔场密度差异均较大(C>0)。棘头梅童鱼季节性分布和集聚强度差异较为明显(2/>1;为标准差,为均值), 其集聚强度依次为: 秋季>春季>冬季(图2)。
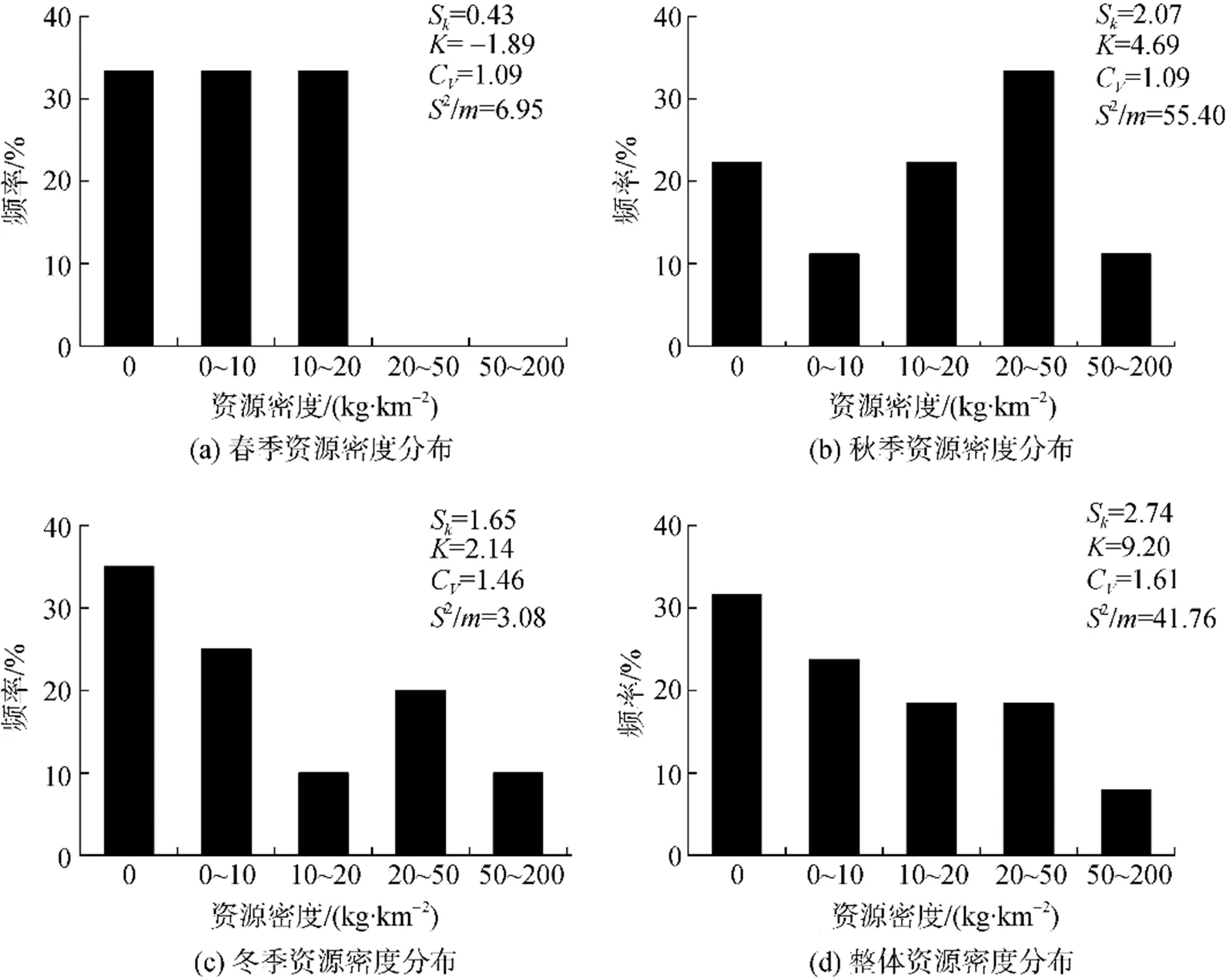
图2 棘头梅童鱼资源密度统计
棘头梅童鱼资源密度高值区主要分布在伶仃洋海域。相对于春季, 冬季资源密度高值区向河口方向移动, 而秋季资源密度高值区向外海方向移动(图3)。调查期间, 春、冬两季在万山群岛站位处未捕捞到棘头梅童鱼, 即该区域棘头梅童鱼资源密度为0, 而秋季有较高的资源密度。
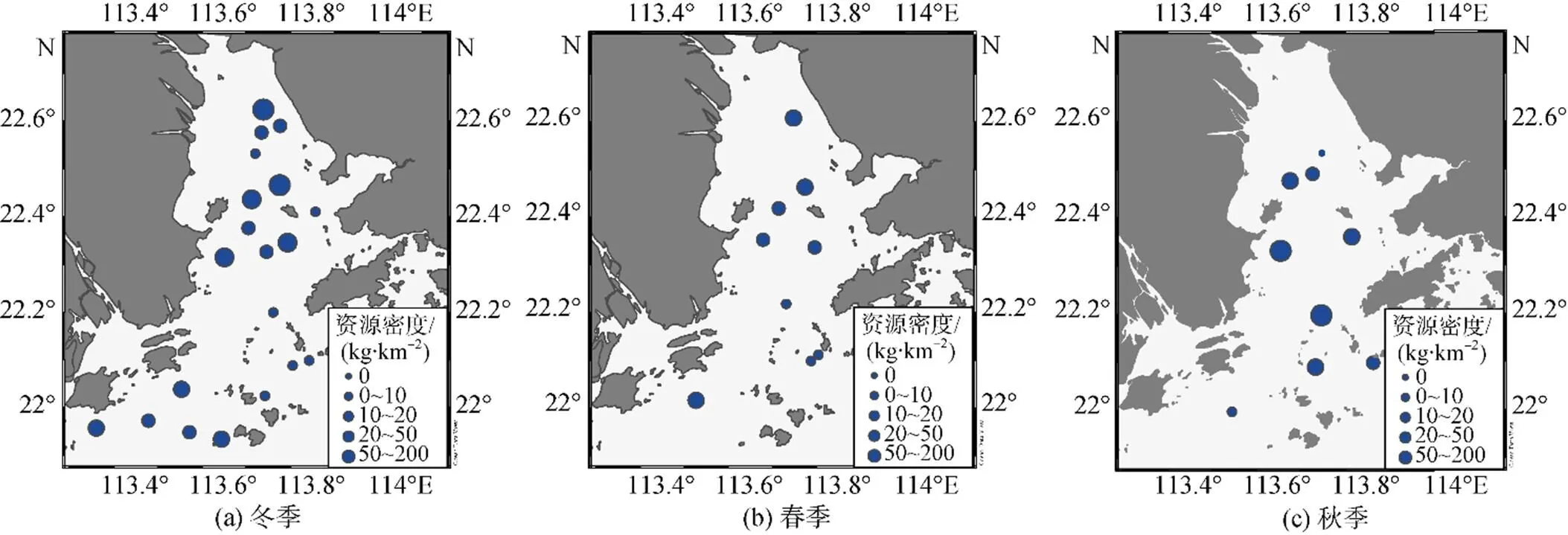
图3 棘头梅童鱼资源密度分布
2.2 共线性检验
本研究选取的9个环境因子中, Season、BpH、Lati、BT的VIF值均≥10, Long、Depth、CHL、BDO、BS的VIF值<10。依据本研究中共线性检验规则, 把Season、BpH、Lati、BT逐步加入到VIF值<10的环境因子中再次共线性检验, 而Lat、BT仍未通过共线性检验, 即移除Lat、BT后, 其余的预测环境因子VIF值均<10(表1)。
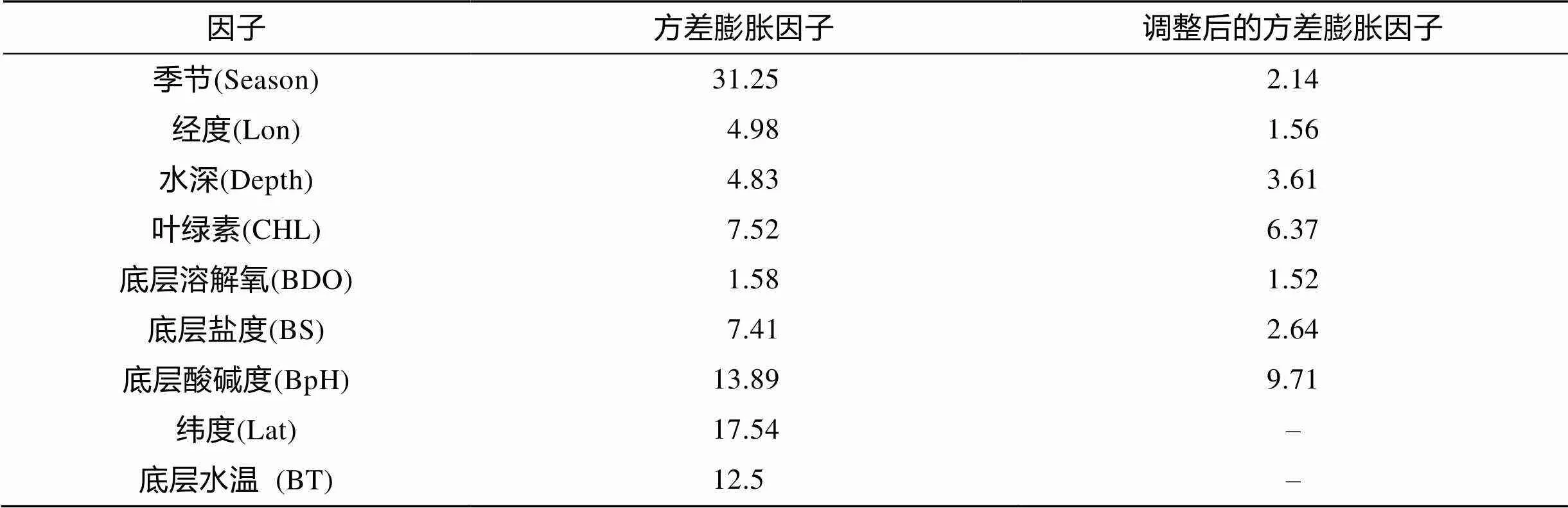
表1 预测变量共线性的检验结果
注: “–”表示去除LAT、BT因子
2.3 最佳模型因子的选择
为降低相关因素对模型因子选择的影响, 采用单因子模型AIC值与初始GAM模型AIC值作对比(表2)。
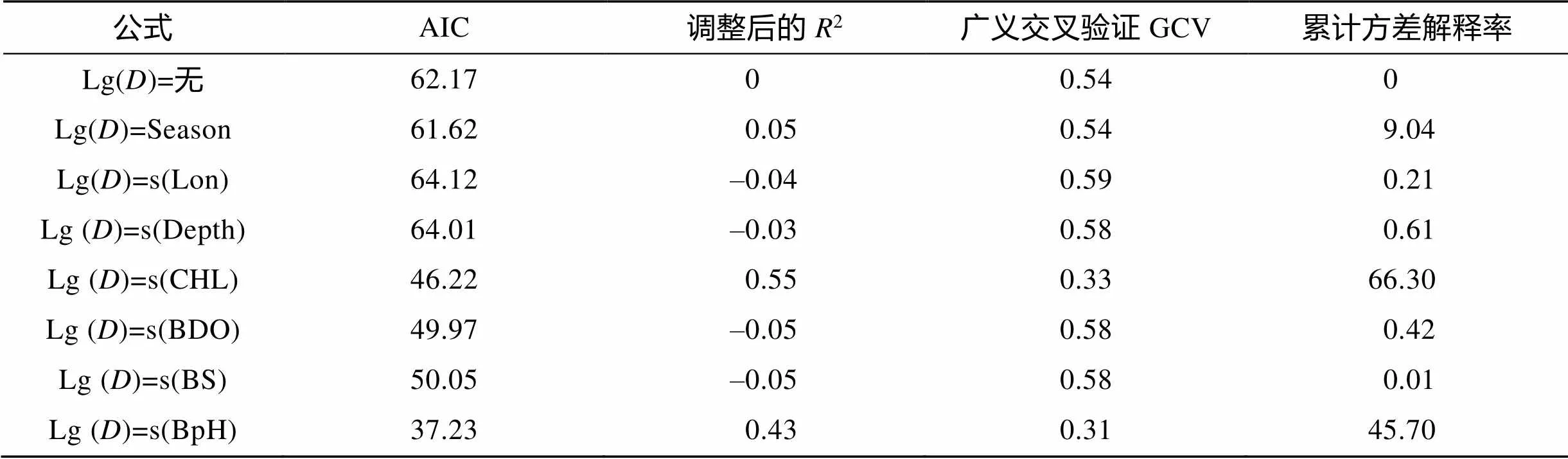
表2 单因子GAM模型参数
各因子的方差解释率差异较大, 其中CHL、BpH对棘头梅童鱼资源的方差贡献率最高(表3)。Lon 和Depth 单因子GAM模型AIC值大于初始GAM模型AIC值, 将剩余的Season、CHL、BDO、BS、BpH等因子进行全子集回归, 挑选包含CHL、BpH且最大调整后2的变量组合作为本研究最佳GAM模型的选择因子(图4)。
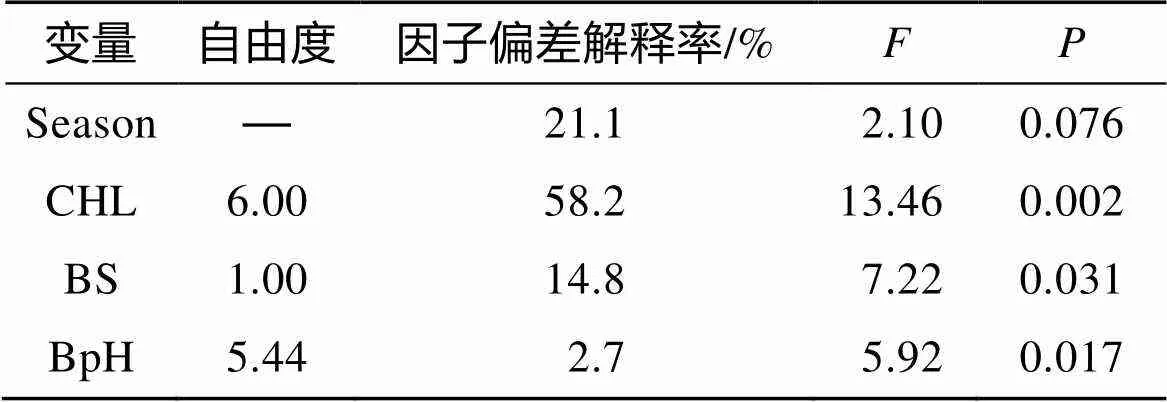
表3 广义加性模型模型分析结果
注: 由于Season为分类离散变量, 无自由度, 用“—”表示。
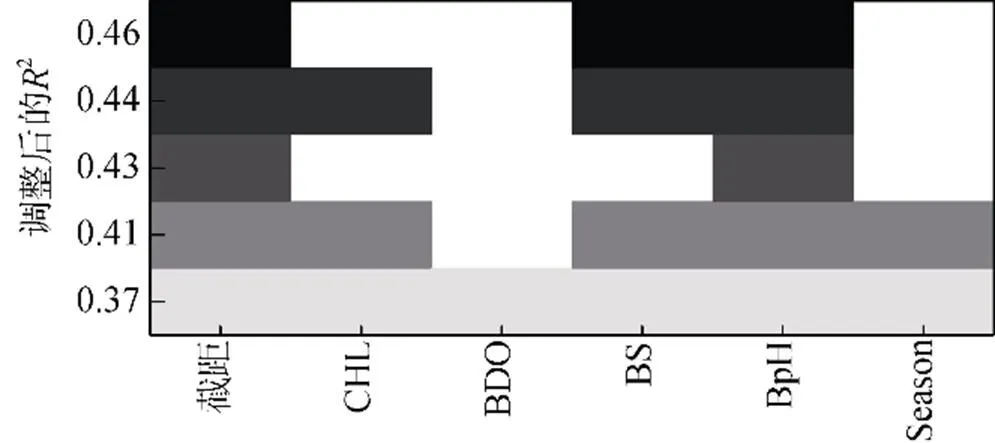
图4 不同子集大小的最佳模型
珠江河口棘头梅童鱼最佳GAM 模型包含CHL、BS、BpH、Season这4个环境要素, 其具体表达式为:
Lg()=0.46+0.28Season+s(CHL)+s(BS)+s(BpH)
其中, 截距为0.46, Season的斜率为0.28, AIC值为2.90。最佳模型对资源密度的累计方差解释率为96.8%, 调整后的2为0.901(<0.001)。其中, CHL贡献率最高, 为58.2%(表3)。
对筛选出的GAM模型进行检验, 残差分布基本符合正态分布, 棘头梅童鱼最佳GAM模型符合要求(图5)。

图5 最佳GAM模型检验
2.4 资源分布与环境因子的关系
分析最优GAM模型中各环境因子的响应特点发现, BpH相较于其他两个因子的置信误差区间较大, 当BpH>9.6时, 棘头梅童鱼资源密度开始大幅度下降(图6a)。CHL在3.1~5 mg·m–3时, 棘头梅童鱼资源密度随着叶绿素浓度的上升而下降, 但置信水平逐渐增加。CHL在5~6.2 mg·m–3范围时, 其资源密度随着CHL值的增加而上升且该部分置信水平较高。当CHL大于6.2 mg·m–3时, 资源密度呈现先降低然后上升的趋势(图6b)。BS与资源密度呈负相关关系, 但资源密度随着BS变化而波动较小(图6c)。
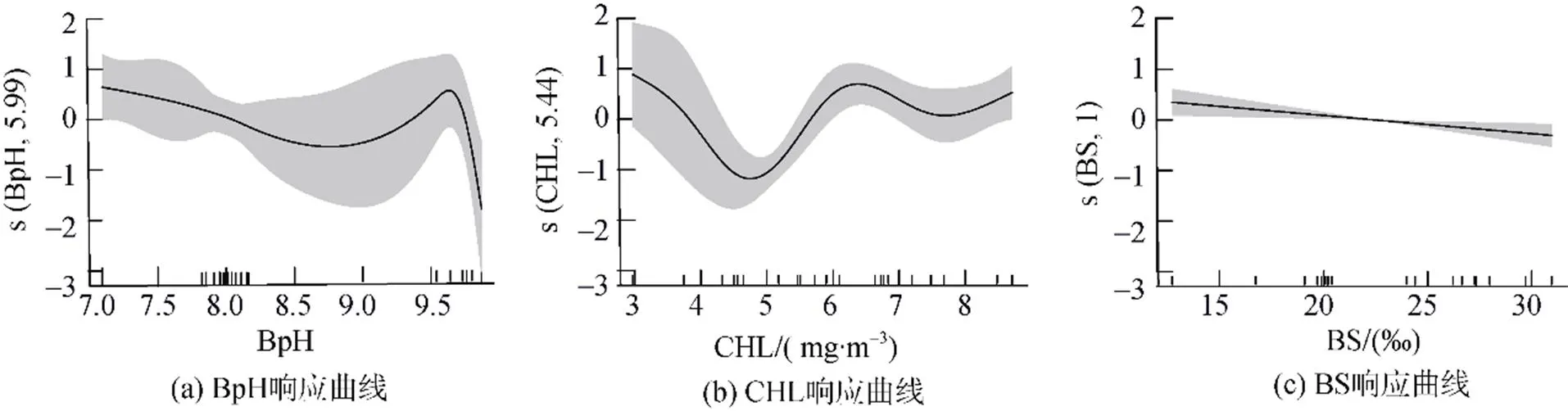
图6 关键环境因子对珠江口棘头梅童鱼资源密度的影响
3 讨论
3.1 棘头梅童鱼资源密度的时空分布
珠江河口棘头梅童鱼资源密度分布具有明显的季节变化特征, 秋季资源密度最高, 冬季次之, 春季最低, 该规律与珠江口的其他鱼类的季节变动趋势基本一致[15-16]。棘头梅童鱼虽一年多次产卵, 但冬季和春季是其产卵高峰期, 该段时期出现大量的棘头梅童鱼幼体, 体质量较低, 秋季是棘头梅童鱼补充群体的育肥期, 该时期棘头梅童鱼生物量明显增加, 因此秋季棘头梅童鱼资源密度明显高于其他季节。
棘头梅童鱼作为咸淡水种类, 其空间分布主要与咸淡水区域的位置变化有关。通过进一步分析调查数据, 发现冬季出现棘头梅童鱼主要集中在伶仃洋水域和珠江支流西江河口区域(图7), 结合珠江流域河流径流量推测导致该现象的主要原因是珠江河口分别在伶仃洋水域和西江河口区域形成了较高生产力的咸淡水区域。珠江干流冲淡水汇入伶仃洋水域, 进入伶仃洋水域平均径流量约为5 668 m3/s, 约占珠江流域出海总量的53.4%[17], 西江是珠江流域的最大支流, 其径流量占珠江流域总径流量的28.3%[18],潮流和径流促进咸淡水混合, 相对于其他区域, 河口的咸淡水混合强度最大[19-20]。咸淡水混合区域汇集了大量的营养盐, 促进浮游动植物的生长, 初级生产力较高[21], 适宜鱼类栖息, 在珠江干流河口和西江河口区域调查站点均具有较高的资源密度。西江流域冲淡水和外海高盐度水在季风影响下分成不同分支汇入不同的方向, 削弱了西江河口冲淡水向南扩散的动量, 西江河口区域咸淡水面积相对于珠江干流河口水域的咸淡水区域小[22]。此外, 棘头梅童鱼属于短距离洄游以及集聚型鱼类, 不做长距离的洄游, 珠江口区域的棘头梅童鱼属同一群体[23], 进一步验证了此推测。
图7 冬季渔业资源重心
棘头梅童鱼渔业资源高值区在秋季明显靠近河口外部区域, 推测秋季是丰水期, 河流径流量大, 珠江口冲淡水向外海区域扩张, 扩大了棘头梅童鱼等咸淡水种类的分布范围。冬季属于枯水期, 外海高盐水区域向河口入侵, 盐淡水区域向河口收缩, 棘头梅童鱼向河口方向移动[16, 23]。
3.2 GAM模型因子筛选
GAM模型因子筛选过程较为灵活, 采用不同的方法可以筛选出不同的最优变量组合, 设置不同光滑函数的(节点, Knots)值以及修改逐步回归中环境因子的添加顺序均会对最佳环境因子的筛选结果产生影响, 通常会根据数据情况、目标鱼类的生活史资料等因素采取不同的方法筛选最佳模型变量组合[24]。如在南印度洋长鳍金枪鱼渔获率与水深、温度关系的研究中, 采用了逐步回归法且以AIC值和显著性水平为准则建立长鳍金枪鱼渔获率的GAM模型[25]; 在长江口鱼类群落时空分布特征的研究中, 采用全子集回归的形式并以AIC作为筛选标准确定最佳GAM变量组合[10], 在沙丁鱼等中上层鱼类的研究中, 将GAM 和GLM相结合探究其渔获量与环境之间的关系[26]。本研究将逐步回归法和全子集回归法相结合, 筛选出季节、叶绿素、底层盐度、底层酸碱度作为构建最佳棘头梅童鱼GAM模型的最佳变量组合, 该方法不仅对资源密度有较高的方差解释率, 而且在评价更多可能模型的同时, 提高了模型的运算效率。无论采取何种方法准则进行环境因子筛选, 都需要与研究背景结合, 以达到统计学“最佳”和生物生态学“最佳”的结合。
3.3 渔场环境要素对棘头梅童鱼资源的影响
海洋生物的栖息地偏好和生活史特征均与海洋水文环境有着密切的关系, 渔业资源的时空分布受盐度、叶绿素浓度等环境因子影响较大[10, 27-28]。从鱼类行为学分析, 盐度变化是支配鱼类活动的重要因素[29], 当盐度发生明显变化时, 会改变鱼类体内外的渗透压平衡, 从而影响其生长和摄食活动[30], 水团和水流会直接影响鱼类的生长速率和生活史各个阶段的开始时间, 进而影响鱼类种群发育[30-31]。叶绿素浓度作为浮游植物生物量的主要指标通过摄食关系和食物网传递与渔业资源密度产生效应关系[7], 其主要影响棘头梅童鱼饵料生物的丰度分布, 影响棘头梅童鱼的食性和分布[32]。
本研究中底层盐度对棘头梅童鱼资源密度的贡献度小于叶绿素, 且棘头梅童鱼资源密度随着底层盐度的变化波动较小, 其主要原因是棘头梅童鱼盐度适应范围广且珠江口咸淡水区域底层盐度变化幅度较小, 盐度不是影响珠江口棘头梅童鱼资源密度的首要因素。鱼类对水质的酸碱度均有一定的耐受阈值, 过高或过低的酸碱度对生物生长发育、繁殖都会带来不利的影响, 特别是对底层生物影响大于中上层生物[33-34], 当底层pH大于9.6时, 棘头梅童鱼资源密度快速下降, 而棘头梅童鱼食性以底层或底栖生物为主[35], 推测pH对棘头梅童鱼时空分布不仅有直接影响, 还会通过影响其饵料分布产生间接影响。
受中国伏季休渔政策的限制, 夏季采样限制, 因而本研究仅采用了春、秋、冬3个季节的调查数据, 虽然满足建模数据的基本要求, 但其时间序列不够长, 存在一定的偶然性和不确定性。除了物理环境因素以外, 非生物因素如饵料丰度也是影响海洋生物的分布的重要驱动因素之一。在今后的研究中, 应增加调查的频次和调查的覆盖范围, 在建模过程中添加非生物因素, 考虑环境变量间的交互作用, 以期得到更全面的结果。
[1] 张帅, 李敏, 闫帅, 等. 基于线粒体Cyt基因序列的棘头梅童鱼种群遗传结构[J]. 中国水产科学, 2021, 28(1): 90-99.
ZHANG Shuai, LI Min, YAN Shuai, et al. Population genetic structure analysis of big head croaker () based on mitochondrial cytochrome b gene sequences[J]. Journal of Fishery Sciences of China, 2021, 28(1): 90-99.
[2] 农业部渔业局. 国家重点保护经济水生动植物资源名录(第一批)[DB/OL]. (2017-12-12). http://jiuban.moa. gov.cn/zwllm/tzgg/gg/201006/t20100606_1538153.htm. 2007.
People’s Republic of China Ministry of Agriculture Proclamation No.948. State list of key economic aquatic animal and plant resources (first batch)[DB/OL]. (2017-12-12). http://jiuban.moa.gov.cn/zwllm/tzgg/gg/ 201006/t20100606_1538153.htm.2007.
[3] 何宝全, 李辉权. 珠江河口棘头梅童鱼的资源评估[J]. 水产学报, 1988, 12(2): 125-134.
HE Baoquan, LI Huiquan. Stock assement ofin Pearl River Estuary[J]. Journal of Fisheries of China, 1988, 12(2): 125-134.
[4] 熊朋莉, 陈作志, 侯刚, 等. 珠江河口棘头梅童鱼生物学特征的年代际变化[J]. 南方水产科学, 2021, 17(6): 32-38.
XIONG Pengli, CHEN Zuozhi, HOU Gang, et al. Decadal change in biological traits ofin Pearl River Estuary[J]. South China Fisheries Science, 2021, 17(6): 32-38.
[5] 陆尧. 粤西海域主要经济鱼类产卵场时空分布的环境效应研究[D]. 上海: 上海海洋大学, 2020.
LU Yao. Enviromental effects of spatial-temporal distribution of spawning ground of major economic fishes in the western Guangdong waters, South China Sea[D]. Shanghai: Shanghai ocean university, 2020.
[6] GUISAN A, THOMAS C E, TREVOR H. Generalized linear and generalized additive models in studies of species distributions: setting the scene[J]. Ecological Modelling, 2002, 157(2): 89-100.
[7] 侯刚, 冯钰婷, 陈妍颖, 等. 北部湾二长棘犁齿鲷时空分布及其与环境因子的关系[J]. 广东海洋大学学报, 2021, 41(4): 8-16.
HOU Gang, FENG Yuting, CHEN Yanying, et al. Spatiotemporal distribution of Threadfin Porgyin Beibu Gulf and its relationship with environmental factors[J]. Journal of Guangdong Ocean University, 2021, 41(4): 8-16.
[8] VENABLESW N, DICHMONT C M. GLMs, GAMs and GLMMs: An overview of theory for applications in fisheries research[J]. Fisheries Research, 2004, 69(2/3): 319-337.
[9] WANG D L, LI J Y, YU J, et al. Response to environmental factors of spawning ground in the Pearl River Estuary, China[J]. Journal of Marine Science and Engineering, 2021, 9(7): 763.
[10] 马金, 黄金玲, 陈锦辉, 等. 基于GAM的长江口鱼类资源时空分布及影响因素[J]. 水产学报, 2020, 44(6): 936-946.
MA Jin, HHUANG Jinling, CHEN Jinhui, et al. Analysis of spatiotemporal fish density distribution and its influential factors based on generalized additive model (GAM) in the Yangtze River estuary[J]. Journal of Fisheries of China, 2020, 44(6): 936-946.
[11] Kabacoff R I. R语言实战[M]. 北京: 人民邮电出版社, 2013: 282-297.
KABACOFF R I. R in action[M]. Beijing: Posts and Telecom Press, 2013: 282-297.
[12] 徐姗楠, 郭建忠, 陈作志, 等. 胶州湾鱼类资源量的时空分布特征[J]. 水产学报, 2019, 43(7): 1615-1625.
XU Shannan, GUO Jianzhong, CHEN Zuozhi, et al. Tempo-spatial distribution characteristics of fish resources in Jiaozhou Bay[J]. Journal of Fisheries of China, 2019, 43(7): 1615-1625.
[13] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995: 59-71.
ZHAN Bingyi. Fish stock assessment[M]. Beijing: China Agriculture Press, 1995: 59-71.
[14] 蔡忠建. 对描述性统计量的偏度和峰度应用的研究[J].北京体育大学学报, 2009, 32(3): 75-76.
CAI Jianzhong. On the application of deviation and kurtosis in descriptive statistics[J]. Journal of Beijing Sport University, 2009, 32(3): 75-76.
[15] 李开枝, 尹健强, 黄良民, 等. 珠江口伶仃洋海域底层游泳动物的季节变化[J]. 生态科学, 2012, 31(1): 2-8.
LI Kaizhi, YIN Jianqiang, HUANG Liangmin, et al. Seasonal variations of bottom nekton in the Lingding Bay of Pearl River Estuary, China[J]. 2012, 31(1): 2-8.
[16] 王迪, 林昭进. 珠江口鱼类群落结构的时空变化[J]. 南方水产, 2006, 2(4): 37-45.
WANG Di, LIN Zhaojin. Spatial and temporal variations of fish community structure in the Pearl River Estuary waters[J]. South China Fisheries Science, 2006, 2(4): 37-45.
[17] 陈炳禄, 张云霓, 王志刚, 等. 伶仃洋水文特征与水质变化趋势分析[J]. 重庆环境科学, 2002, 24(2): 69-72.
CHEN Binglu, ZHANG Yunni, WANG Zhigang, et al. Characteristic of Lingdingyang marine environment[J]. Chongqing Environmental Science, 2002, 24(2): 69-72.
[18] 陈玲舫, 陈子燊, 黄强. 珠江河口磨刀门水道的亚潮振荡特征及其对水文气象要素的响应[J]. 海洋通报, 2014, 33(2): 126-131.
CHEN Lingfang, CHEN Zishen, HUANG Qiang. Sub- tide characteristics of the Modaomen watercourse in the Pearl River Estuary and its response to hydrometeorology elements[J]. Marine Science Bulletin, 2014, 33(2): 126-131.
[19] 范中亚, 林澍, 曾凡棠, 等. 珠江口门枯季动力过程及盐度分布特征[J]. 热带地理, 2013, 33(4): 400-406.
FAN Zhongya, LIN Shu, ZENG Fantang, et al. Characteristics of hydrodynamics and salinity in the estuaries of the Pearl River in dry season[J]. Tropical Geography, 2013, 33(4): 400-406.
[20] 袁菲, 卢陈, 杨裕桂, 等. 珠江口伶仃洋及磨刀门盐淡水混合特征及机制分析[J]. 海洋环境科学, 2021, 40(3): 361-368, 378.
YUAN Fei, LU Chen, YANG Yugui, et al. Comparative analysis of the characteristics and mechanism of the salt-fresh water mixing in Lingdingyang and Modaomen Estuary, Pearl River Estuary[J]. Marine Environmental Science, 2021, 40(3): 361-368, 378.
[21] 施玉珍, 赵辉, 王喜达, 等. 珠江口海域营养盐和叶绿素的时空分布特征[J]. 广东海洋大学学报, 2019, 39(1): 56-65.
SHI Yuzhen, ZHAO Hui, WANG Xida, et al. Distribution characteristics of nutritive salts and chlorophyllin the Pearl River Estuary[J]. Journal of Guangdong Ocean University, 2019, 39(1): 56-65.
[22] 张燕, 夏华永, 钱立兵, 等. 2006年夏、冬季珠江口附近海域水文特征调查分析[J]. 热带海洋学报, 2011, 30(1): 20-28.
ZHANG Yan, XIA Huayong, QIAN Libing, et al. Analysis on hydrological characteristics off the Pearl River Estuary in summer and winter of 2006[J]. Journal of Tropical Oceanography, 2011, 30(1): 20-28.
[23] XU K T, CHEN W J, HUANG S H, et al. Environmental drivers of the functional structure of fish communities in the Pearl River Estuary[J]. Estuarine, Coastal and Shelf Science, 2021, 263: 107625.
[24] GUISAN A, THUILLER M, ZIMMERMANN N E. Habitat suitability and distribution models: With applications in R (ecology, biodiversity and conservation)[M]. Cambridge: Cambridge University Press, 2017: 195-202.
[25] 谢笑艳, 汪金涛, 陈新军, 等. 南印度洋长鳍金枪鱼渔获率与水深温度关系研究[J]. 南方水产科学, 2021, 17(5): 86-92.
XIE Xiaoyan, WANG Jintao, CHEN Xinjun, et al. Study on relationship between albacore catch rate and water depth and temperature in South Indian Ocean[J]. South China Fisheries Science, 2021, 17(5): 86-92.
[26] Agenbag J J, Richardson A J, Demarcq H, et al. Estimating environmental preferences of South African pelagic fish species using catch size- and remote sensing data[J]. Progress in Oceanography, 2003, 59(2): 275-300.
[27] 胡丽娟, 宋超, 耿智, 等. 长江口凤鲚仔稚鱼主要繁殖季节的时空分布特征[J]. 中国水产科学, 2021, 28(9): 1152-1161.
HU Lijuan, SONG Chao, GENG Zhi, et al. Temporal and spatial distribution oflarvae and juveniles in the Yangtze Estuary during primary breeding season[J]. Journal of Fishery Sciences of China, 2021, 28(9): 1152-1161.
[28] WANG D L, LI J Y, YU J, et al. The role of environmental factors on the fishery catch of the Squidin the Pearl River Estuary, China[J]. Journal of Marine Science and Engineering, 2021, 9(2): 131.
[29] 陈新军. 渔业资源与渔场学[M]. 北京: 海洋出版社, 2004: 125-127.
CHEN Xinjun. Fisheries Resources and fisheries Science[M]. Beijing: Ocean Press, 2004: 125-127.
[30] ZHANG Y L, BIN D X, YUP J, et al. Comparison of habitat models in quantifying the spatio-temporal distribution of small yellow croaker () in Haizhou Bay, China[J]. Estuarine, Coastal and Shelf Science, 2021, 261: 107512.
[31] 邹易阳, 薛莹, 麻秋云, 等. 应用栖息地适宜性指数研究海州湾小黄鱼的空间分布特征[J]. 中国海洋大学学报(自然科学版), 2016, 46(8): 54-63.
ZOU Yiyang, XUE Ying, MA Qiuyun, et al. Spatial distribution ofin Haizhou Bay based on habitat suitability index[J]. Periodical of Ocean University of China, 2016, 46(8): 54-63.
[32] 党莹超, 戴小杰, 吴峰. 北太平洋长鳍金枪鱼的垂直分布及其与环境因子的关系[J]. 海洋湖沼通报, 2021, 43(6): 92-99.
DANG Yingchao, DAI Xiaojie, WU Feng. Vertical distribution of albacorein north Pacific and its relation to environmental factors[J]. Transactions of Oceanology and Limnology, 2021, 43(6): 92-99.
[33] 柴学军, 胡则辉, 徐君卓, 等. 盐度和pH对日本黄姑鱼幼鱼耗氧率和排氨率的影响[J]. 浙江海洋学院学报(自然科学版), 2009, 28(2): 146-150.
CHAI Xuejun, HU Zehui, XU Junzhuo. Effect of salinity and pH on oxygen consumption rate and ammonia excretion rate in Juvenile[J]. Journal of Zhejiang Ocean University (Natural Science), 2009, 28(2): 146-150.
[34] 唐贤明, 隋曌, 田景波, 等. 盐度对大菱鲆幼鱼耗氧率和排氨率的影响[J]. 南方水产, 2006, 2(4): 54-58.
TANG Xianming, SUI Zhao, TIAN Jingbo, et al. Effects of salinity on metabolic rate of juvenile turbot ()[J]. South China Fisheries Science, 2006, 2(4): 54-58.
[35] 宋业晖, 薛莹, 徐宾铎, 等. 海州湾3种石首鱼的食物组成和生态位重叠[J]. 水产学报, 2020, 44(12): 2017-2027.
SONG Yehui, XUE Ying, XU Binduo, et al. Composition of food and niche overlap of three Sciaenidae species in Haizhou Bay[J]. Journal of Fisheries of China, 2020, 44(12): 2017-2027.
Spatiotemporal distribution ofin the Pearl River Estuary and its relationship with environmental factors
XIONG Peng-li1, 3, 4, XU Shan-nan2, 3, CHEN Zuo-zhi2, 3, ZHANG Shuai2, JIANG Pei-wen2, 3, 4, FAN Jiang-tao2, 3
(1. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China; 2. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Open-Sea Fishery Development, Ministry of Agriculture and Rural Affairs, Guangzhou 510300, China; 3. Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511548, China; 4. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China)
Based on the bottom trawl survey data of the Pearl River Estuary (PRE) in 2019–2020 and the environmental data obtained via remote sensing andobservation, a generalized additive model (GAM) with resource density as the response variable was established to analyze the spatiotemporal distribution ofin the PRE and to screen the key environmental factors affecting this distribution. Results showed thatwas mainly distributed in Lingdingyang. Compared with winter and spring, the species moved southward in autumn. The optimal combination of GAM model variables includes bottom salinity (BS), chlorophyll a (CHL), bottom pH (BpH), and season; 96.8% of the total variation in resource density was explained by these factors. Of all factors, the most important was CHL, accounting for 14.3%, followed by season, BS, and BpH with contributions of 21.1%, 14.8%, and 2.7%, respectively. The results of this study can provide an effective scientific basis for the conservation of plum fish resources.
spatiotemporal distribution; impacts of environmental factors; generalized additive models (GAM); Collichthy lucidus; the Pearl River Estuary
Jan. 16, 2022
S931.4
A
1000-3096(2022)08-0079-09
10.11759/hykx20220116001
2022-01-16;
2022-03-17
国家重点研发计划(2018YFD0900902); 南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0605); 中央级公益性科研院所基本科研业务费专项资金项目(2020TD05, 2021SD01)
[National Key Research and Development Program of China, No. 2018YFD0900902; Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory, No. GML2019ZD0605; Central Public-Interest Scientific Institution Basal Research Fund South China Sea Fisheries Research Institute CAFS, Nos. 2020TD05, 2021SD01]
熊朋莉 (1997—), 女, 硕士研究生, 研究方向为渔业资源, E-mail: xiong_pengli@163.com; 范江涛(1987—),通信作者, 男, 助理研究员, 从事渔业资源和渔情预报研究, E-mail: tianxiahaiyin@163.com
(本文编辑: 赵卫红)
