不同始温浸种对圆齿野鸦椿休眠种子萌发特性的影响研究
2022-10-08阳先波姜雪茹吴慧平涂淑萍蔡军火
阳先波,姜雪茹,吴慧平,冉 芳,涂淑萍,蔡军火,张 隆
(江西农业大学 园林与艺术学院,江西 南昌330045)
【研究意义】圆齿野鸦椿(Euscaphis konishii)属省沽油科(Staphyleaceae)野鸦椿属(Euscaphis)常绿小乔木,是中国南方特有的药赏两用珍贵乡土树种,其枝条、果皮等部位提取物具有较好的抗炎抑菌、抗癌作用[1-3],其红果吉祥喜庆,且挂果期长,经冬不凋,具有极大的推广应用价值。但是圆齿野鸦椿种子具有深休眠特性,发芽率低、出苗不整齐,是影响其苗木培育和规模化生产的主要原因。因此,从缩短催芽周期角度开展圆齿野鸦椿种子萌发特性相关研究对其种苗的高效繁育具有重要意义。【前人研究进展】以往对圆齿野鸦椿种子的研究表明,种胚发育不完全且种子内含有发芽抑制物是导致其休眠的主要因素[4-6]。在种子采收后直接播种,至第二年春季发芽率仍为零。而在种子采收后湿沙自然层积约450 d后进行播种,发芽率可达到75%[7],说明长时间的湿沙层积促进了种胚的发育和抑制物的减少。圆齿野鸦椿种胚发育存在不一致性[5],在湿沙层积几个月后,通过播种前对种子进行一定温度的热水浸种,能够促进部分休眠种子萌发[8],如张莉梅等[9]的研究发现采用湿沙4 ℃沙层积+60 ℃温水浸泡(自然冷却)对圆齿野鸦椿种子进行处理,仅用90~120 d 萌发率峰值可达48%,而欧斌等[10]将自然沙藏150~180 个月的圆齿野鸦椿种子用始温65~68 ℃热水浸种,可使萌发率达到51%~65%。【本研究切入点】最佳浸种温度可能与湿沙层积时间有一定的联系。而在不同温度热水浸种处理下,圆齿野鸦椿种子的生活力、内源发芽抑制物及内含物质如何变化的还不得而知。【拟解决的关键问题】因此,本实验研究了不同浸种温度对湿沙层积120 d的圆齿野鸦椿种子发芽率和生活力的影响,比较不同浸种处理过程中种子内含物的变化及对种子内源发芽抑制物的影响,并将各项生理指标与萌发率和生活力进行相关性分析,探讨热水浸种处理下种子的生理变化机制,以期为缩短圆齿野鸦椿育苗周期和促进产业市场化发展提供理论基础。
1 材料与方法
1.1 材料
供试种子来源于江西农业大学花卉基地自然生长的14 年树龄的圆齿野鸦椿母树,采集时间为2020 年11 月初,自然沙藏层积120 d。于2021 年3 月初取出种子,经0.3%高锰酸钾溶液消毒和纯水冲洗后[11],采取5 种方式对种子进行预处理:常温水浸泡24 h(CK);始温50 ℃水浸种,自然冷却24 h(T1);始温60 ℃水浸种,自然冷却24 h(T2)、始温70 ℃水浸种,自然冷却24 h(T3);始温80 ℃水浸种,自然冷却24 h(T4)。
1.2 研究方法
1.2.1 种子萌发实验 采用培养皿滤纸法进行萌发试验[12],人工气候箱温度为25 ℃,湿度55%,光周期为光12 h/暗12 h,当胚根伸长至2 mm 时,视为萌发。每个处理3 个重复,每个重复30 例种子,统计时间持续1 个月,1 个月后未萌发的视为不能发芽。期间及时清理污染发霉的种子并更换滤纸,并定时补充纯水保持滤纸湿润。发芽率(Germination Rate,GR/%)=(萌发种子数量/供试种子数量)×100%。
1.2.2 种子生活力测定 生活力测定采用2,3,5-氯化三苯基四氮唑法(简称TTC法)将经过预处理的种子去除外部硬种皮后纵向切开,采用0.3%TTC 溶液于25 ℃黑暗恒温箱中孵育,15 h 后统计圆齿野鸦椿种胚着色情况[13-15]。每个处理3次重复,每个重复20粒种仁,以沸水处理的种仁为对照。
1.2.3 种子生理生化指标测定 分别取CK、T1、T2、T3处理过程中0,6,12,24 h的种子,去除外部坚硬种皮后,采样进行相关生理指标测定,每个处理3个重复。考马斯亮蓝G-250法测定可溶性蛋白含量[16],蒽酮比色法测定可溶性糖含量[17],氮蓝四唑光还原法测定超氧化物歧化酶(SOD)活性[18],愈创木酚法测定过氧化物酶(POD)活性[18],紫外分光光度法测定过氧化氢酶(CAT)活性[19]。
1.2.4 圆齿野鸦椿种子浸提液的制备与使用 首先,制备质量浓度为0.2 g/mL 的圆齿野鸦椿种子浸提液:在20 mL 规格的玻璃试管中加入7 mL 纯水,将试管放入50 ℃水浴锅中,当试管中水温达到50 ℃时,加入2 g 自然层积120 d 的圆齿野鸦椿种子,然后立即关闭水浴锅,试管口用保鲜膜和皮筋封严,自然冷却24 h 后抽取全部浸提液,定容至10 mL[20-21],以上是50 ℃始温热水浸种(T1)提取液制备步骤。CK、T2(60 ℃)、T3(70 ℃)浸种提取液制备相同。采用以上浸提液浸泡白菜种子24 h,将浸泡后的白菜种子置于对应的浸提液润湿的滤纸上,每个培养皿50 粒,每处理3 个重复,于温度25 ℃、湿度55%,光周期为光8 h/暗16 h的培养箱中培养。72 h后统计白菜种子发芽率(以胚轴弯曲、胚根伸长≥4.0 mm作为萌发标准),用直尺测量白菜种子根长、茎长[13,22]。
1.3 数据分析
采用Excel 2019 和Origin 2018 进行数据整理及绘图。采用SPSS 22.0 进行ANOVA 分析(采用Duncans法进行多重比较)和Spearman相关性分析。
2 结果与分析
2.1 种子发芽率和生活力
5 种不同温度浸种方式对种子发芽率和生活力的影响差异较大。与CK 相比,T1、T2 能显著提高圆齿野鸦椿种子的发芽率,T1 的发芽率最高为43.33%±6.29%,T2 热水浸种发芽率次之,为25.56%±3.63%,而T3、T4 热水浸种发芽率则为0(图1A)。为了明确不同温度热水浸泡引起的发芽率差异是否与种子活性有关,笔者测定了CK、T1、T2、T3 处理后的种子生活力,每个处理均以沸水灭活的种子为对照,结果发现,T1 与CK 浸种后圆齿野鸦椿种子生活力无显著差异,均大于等于70.00%,而T2 浸种使生活力下降至58.33%±7.58%,T3 热水浸种处理后种子已经失去活性(图1A,图1B)。以上说明,始温50 ℃热水浸种在不损害种子生活力的基础上还可显著提高种子发芽率,而始温70 ℃热水浸种则会使种子失去活性。
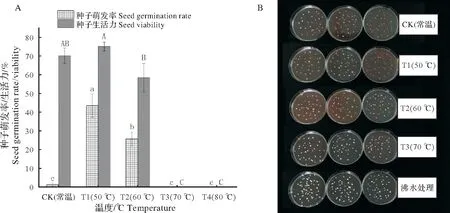
图1 不同始温热水浸种下圆齿野鸦椿种子萌发率和生活力差异Fig.1 Changes of Euscaphis konishii seeds germination rate and viability under seed soaking at different initial temperatures
2.2 种子生理指标
2.2.1 种子可溶性蛋白与可溶性糖含量变化 不同始温浸种处理下,圆齿野鸦椿种子内可溶性蛋白和可溶性糖含量随时间的变化情况如图2 所示。在浸种处理的12 h 内,CK、T1、T2、T3 的可溶性蛋白含量无显著性变化,在浸种处理的24 h 时,T3 的可溶性蛋白含量大幅度上升且显著高于其余处理,而其余处理之间可溶性蛋白含量无显著差异(图2A)。在浸种处理的24 h内,CK的可溶性糖含量呈现先上升后下降再上升的趋势,T1 的可溶性糖含量呈现先上升后下降趋势,T2、T3 的可溶性糖含量一直呈下降趋势;在浸种处理的6 h时,CK 的可溶性糖含量显著高于T2、T3,在12 h时T1的可溶性糖含量显著高于其他处理,在24 h时CK的可溶性糖含量显著高于T1,T1显著高于T2、T3(图2B)。
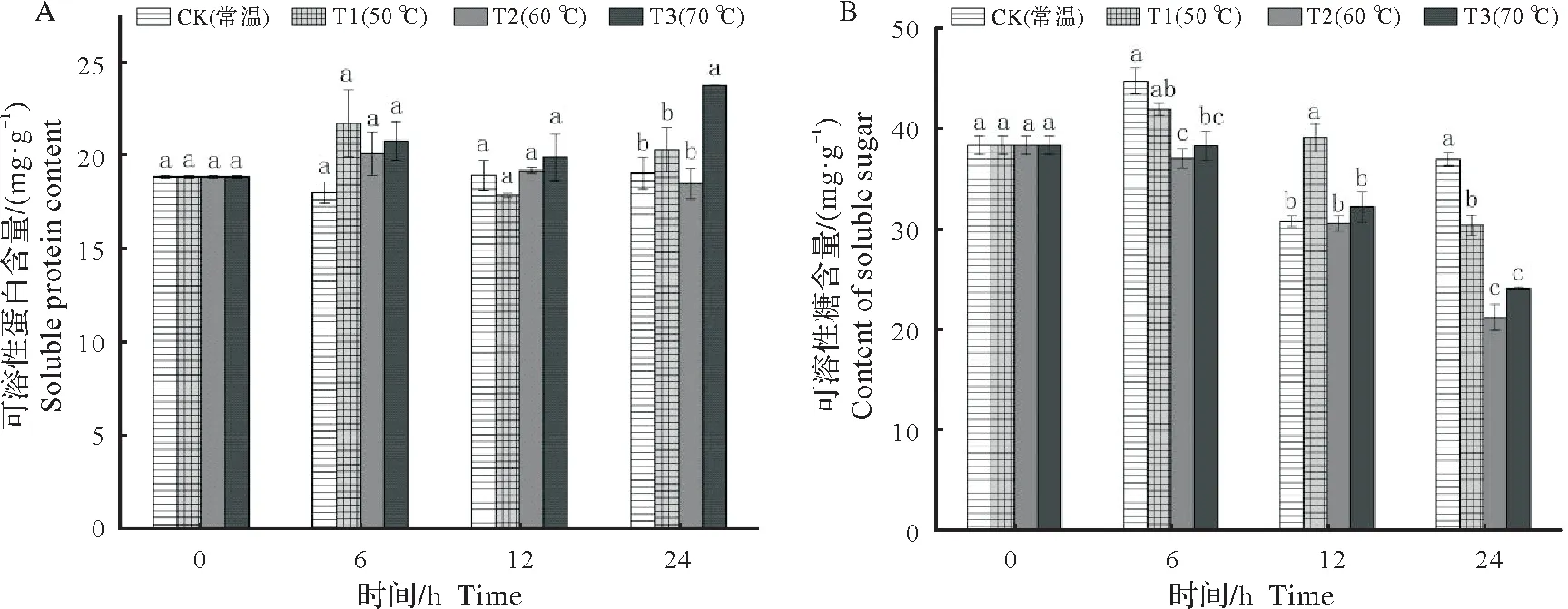
图2 不同始温浸种处理下圆齿野鸦椿种子可溶性糖和可溶性蛋白含量随时间的变化情况Fig.2 The effects of different initial temperatures soaking water treatments on the soluble sugar and soluble protein content of Euscaphis konishii seeds
2.2.2 抗氧化酶活性变化 不同始温浸种处理下,圆齿野鸦椿种子内抗氧化酶活性随时间的变化情况如图3 所示。在浸种处理的6 h 时,T1 的CAT 活性显著高于其他处理,T3 次之,CK 和T2 处理最低;在12 h 时,各个处理之间的CAT 活性没有显著性差异;在24 h 时,CK、T1 处理的CAT 活性显著高于T2、T3,且在整个浸种过程中T2、T3 的CAT 活性一直低于初始值(0 h)(图3 A)。与浸种前(0 h)相比,浸种处理6 h 时CK 的POD 活性显著提高,而T3 的POD 活性大幅度下降;在浸种处理的12 h 时,T2 的POD 活性显著高于其他处理;在浸种处理的24 h 时,CK 的POD 活性显著高于其他处理,T1 次之,而T3 处理的POD 活性在整个浸种过程均较低(图3B)。在浸种处理的6 h 时,T1 的SOD 活性最高,T3 的最低;在浸种处理的12 h 时,CK 的SOD 活性最高,同时T2、T3 活性与未浸种前相比也有了明显地上升;在浸种处理的24 h 时,T1、T2 处理的SOD 活性显著高于T3、T4;在整个过程中CK 和T1 一直保持较高的SOD 酶活性(图3C)。
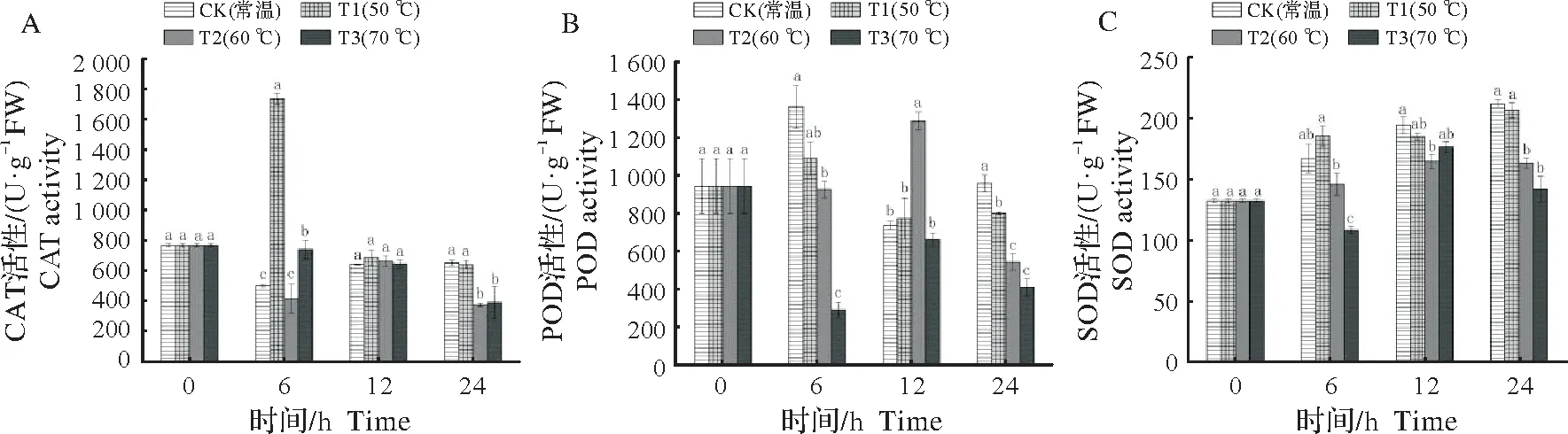
图3 不同始温浸种处理下圆齿野鸦椿种子内抗氧化酶活性的变化Fig.3 The effects of different initial temperature soaking water treatments on antioxidant enzyme activities in Euscaphis konishii seeds
2.2.3 种子生理生化指标相关性分析 为了进一步探究热水浸种过程中种子生理活动的启动与萌发特性的关系,将每个浸种时间段内种子的可溶性糖(SP)、可溶性蛋白(Pro)、抗氧化酶活性与种子最后的萌发率和种子活力进行了相关性分析。由表1可知,浸种处理6 h时的CAT 活性与种子的发芽率呈显著正相关(P<0.05),POD和SOD活性与种子的生活力呈极显著正相关性(P<0.01);浸种处理12 h时的SP与种子发芽率呈显著正相(P<0.05);浸种处理24 h 时的Pro 含量与种子生活力呈极显著负相关(P<0.01),而POD、SOD活性与种子生活力却呈极显著的正相关(P<0.01)。
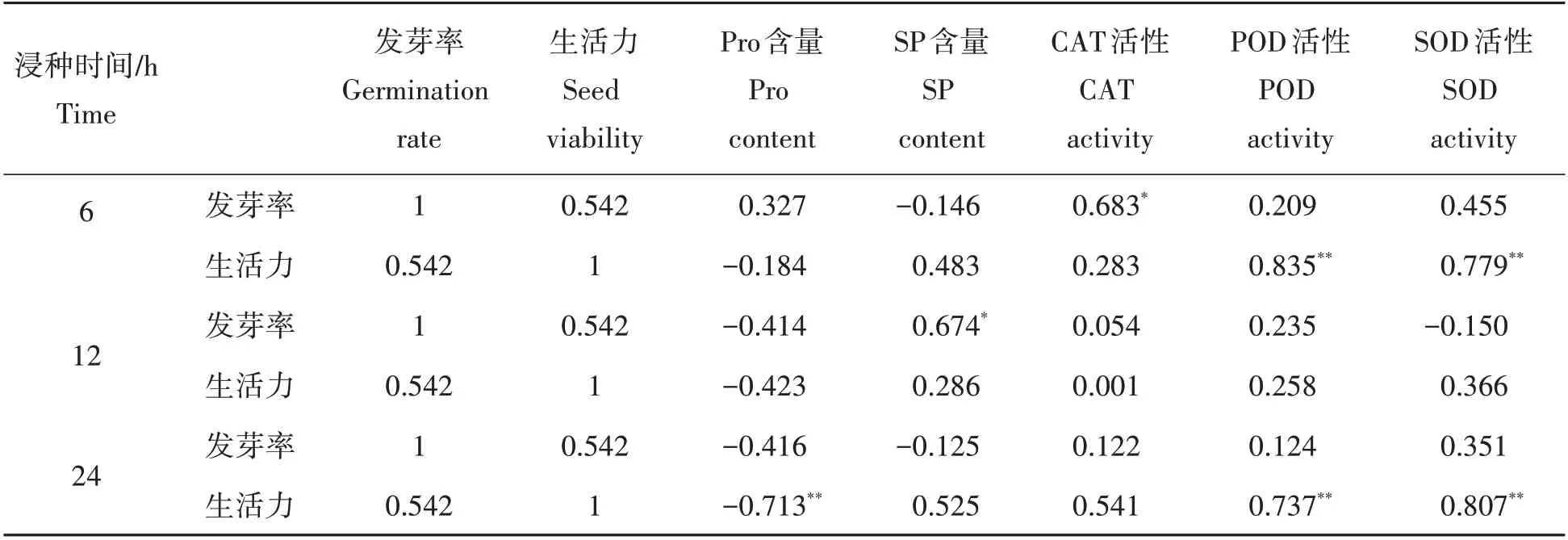
表1 不同浸种时间内圆齿野鸦椿种子发芽率、生活力与生理指标的相关性分析Tab.1 Correlation analysis of germination rate,seed viability between physiological indexes in Euscaphis konishii seeds
2.3 内含抑制物活性
与纯水处理相比,经始温50 ℃(T1)、60 ℃(T2)和70 ℃(T3)热水浸种液处理的白菜种子萌发率显著降低,幼苗根长和茎长却显著增加;T1、T2、T3 浸种液处理之间的白菜种子萌发率和幼苗根长没有显著性差异,而幼苗茎长由高到低依次为T3,T2,T1,纯水(图4 A~C)。综上,T1、T2、T3 浸种液处理显著降低了白菜种子的萌发率,却增加了白菜幼苗的根长和茎长,使得白菜根茎变得十分细长(图4D)。
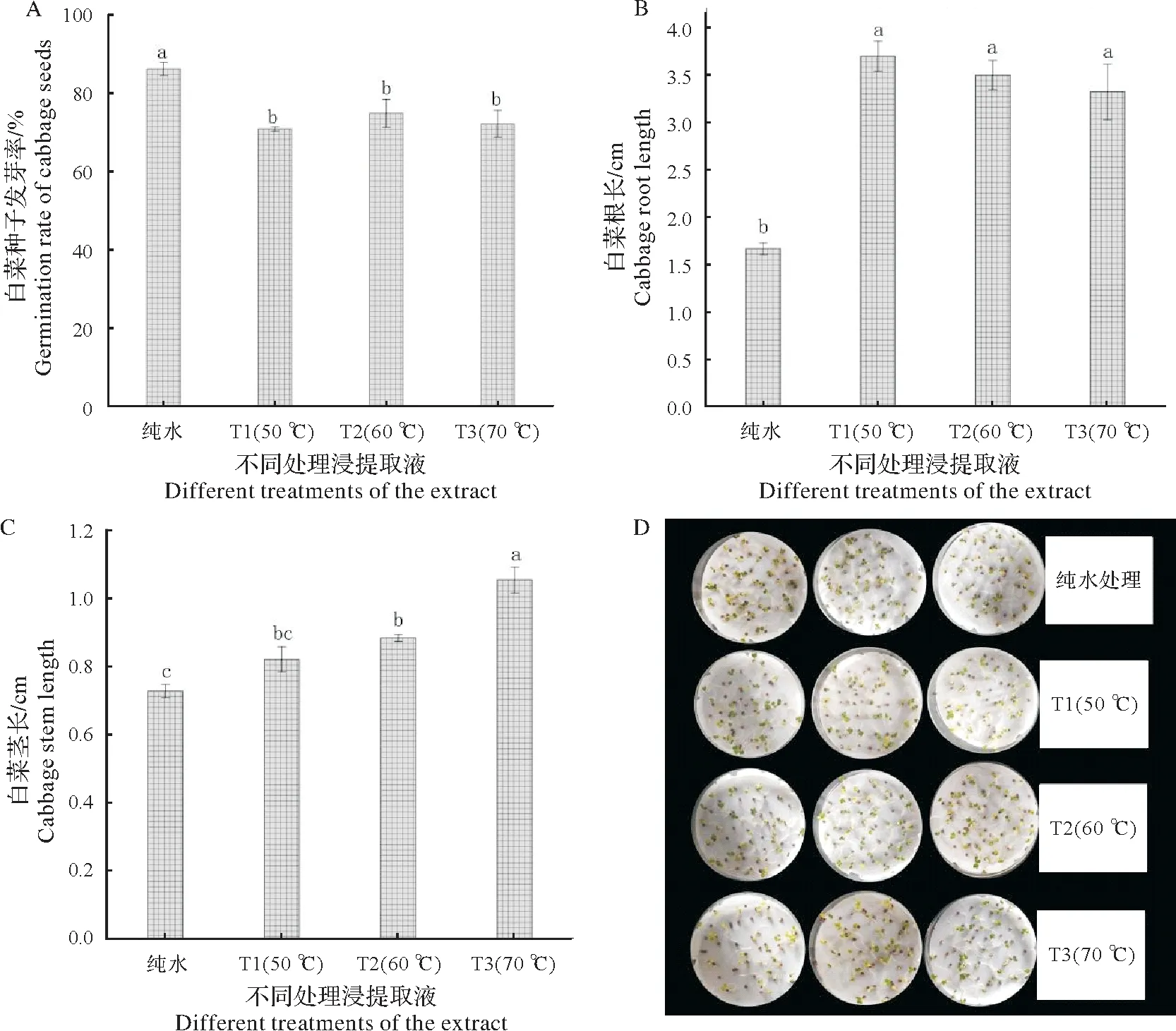
图4 浸种液对白菜种子萌发的影响Fig.4 Effect of seed immersion solution on germination of cabbage seeds
3 讨论与结论
3.1 热水浸种对圆齿野鸦椿种子发芽率和生活力的影响
圆齿野鸦椿种子的休眠类型为形态-生理休眠[12],即休眠原因为胚发育不完全和生理抑制机制强[23-24]。自然沙藏层积120 d 的种子,经常温水浸种24 h 后,萌发率几乎为0,而50 ℃始温热水浸种后平均发芽率可达43.33%±6.29%,种子生活力也与常温水浸种相近,可见50 ℃始热水浸种可明显提高圆齿野鸦椿种子的萌发率,且不会降低种子的整体生活力。当使用60 ℃始温热水进行浸种后,种子生活力开始受到损伤,发芽率也只有25.56%±3.63%,而采用70 ℃和80 ℃始温热水进行浸种的种子已完全失去生活力,所以在进行圆齿野鸦椿种子浸种时应注意严控温度,最好不超过60 ℃。
在实验中笔者观察到,能够发芽的种子绝大多数是浸种后露白的种子。邹双全[7]在圆齿野鸦椿种子自然层积第3 年露白10%时播种,发芽率可达75%,远高于本实验中50 ℃始温热水浸种处理的发芽率。推测原因可能是,层积120 d 的圆齿野鸦椿种子的种胚并没有全部发育成熟,50 ℃始温热水浸种只能使部分发育完全的种胚吸水膨胀后露白进而萌发,而没有萌发却有生活力的那部分种子可能是由于层积时间过短、种胚未发育完全所致。
3.2 热水浸种对圆齿野鸦椿种子内源物质的影响
浸种过程中种子新陈代谢变得旺盛,消耗可溶性糖维持呼吸作用,种子内可溶性糖含量随着时间持续下降[25],这与本实验的研究相一致。Fatemeh 等[26]的研究显示种子在经历外界胁迫时,会通过保持较高的蛋白质合成速率和较低的蛋白质降解速率,维持低渗透势的稳态,以抵御细胞膜受到的损害,这也可能是70 ℃始温水浸泡24 h 时种子内可溶性蛋白含量显著高于其他处理的原因。50 ℃始温热水浸泡处理6 h 时,种子内3 种抗氧化酶的活性均明显升高,说明该处理在一定程度上减轻了活性氧自由基对细胞膜系统造成的伤害[27],有利于种子萌发。而与其他处理相比,在70 ℃始温热水浸泡处理的24 h 内,种子的抗氧化酶活性一直处于较低水平,其原因可能是过高的温度处理破坏了抗氧化酶活性[28]。此外,由相关性分析结果可知,在浸种处理的6 h 时,CAT 活性与萌发率呈显著正相关,12 h 时,可溶性糖含量与发芽率呈显著的正相关,在24 h 时,可溶性蛋白含量与生活力呈极显著的负相关,POD、SOD 活性与生活力呈正相关。以上说明,热水浸种可触发种子内源物质转换,为种子萌发做准备,并且CAT 可能是与圆齿野鸦椿种子萌发率密切相关的酶,而POD 和SOD 与圆齿野鸦椿种子的活力密切相关[29]。
3.3 热水浸种对圆齿野鸦椿种子内发芽抑制物的影响
圆齿野鸦椿种子休眠的主要原因是内含发芽抑制物及种子具有胚后熟特性,即使在经历270 d的变温层积后抑制物活性依旧比较强[4]。与常温水浸种相比,不同始温热水浸种液均显著降低了白菜种子的发芽率,这反向说明了热水浸种处理可稀释圆齿野鸦椿种子内发芽抑制物质的浓度,进而降低内源抑制物质对种子萌发的抑制作用[30]。而与大多数探究种子浸提取液抑制物实验结果不同的是,本实验中圆齿野鸦椿种子热水浸泡液对白菜种子的根、茎都有显著的促伸长作用,但是根茎与CK相比十分细长。
3.4 结论
圆齿野鸦椿种子具有深休眠特性,一般需自然层积450 d 萌发率可达75%。本研究发现,50 ℃始温热水浸种能够使自然层积120 d 的圆齿野鸦椿种子萌发率达到43.33%±6.29%,而没有萌发却有生活力的那部分种子可能是由于层积时间过短、种胚未发育完全所致。热水浸种可激发圆齿野鸦椿种子内源物质转换,降低内源发芽抑制物浓度,CAT 可能是与圆齿野鸦椿种子萌发率密切相关的酶,而POD 和SOD 与圆齿野鸦椿种子的活力密切相关。圆齿野鸦椿种子可能存在种胚发育的不整齐性,有生活力但胚发育不完全的种子在适合的热水浸种环境下仍无法解除休眠,具体原因及解除办法还需要进一步探索。
致谢:江西省教育厅项目(GJJ200446)同时对本研究给予了资助,谨致谢意!
