外源氮对滨海围垦稻田土壤N2O还原速率的影响机制研究
2022-10-03饶旭东薛梦琪张耀鸿
饶旭东,谢 晴,薛梦琪,周 聪,张耀鸿
(南京信息工程大学,气象灾害预报预警与评估协同创新中心/江苏省农业气象重点实验室,江苏 南京 210044)
N2O是仅次于CO2和CH4的第三大温室气体,其百年尺度上的增温潜势约为CO2的298倍[1],对温室效应的贡献约为6%[2],对全球气候变化产生重要影响。农业土壤是最重要的N2O来源;研究发现,农业生产活动排放的N2O占人类活动总排放量的52%[3]。然而,排放到大气中的N2O气体是土壤N2O产生与消耗过程的综合结果。目前,绝大多数研究主要关注于不同土壤中N2O的净排放通量,而忽略了N2O净消耗量的潜力。事实上,土壤不仅可以是大气N2O的源,也可以消耗大气N2O,成为大气N2O的汇。土壤中N2O的消耗过程包括N2O在土壤剖面内扩散传输中被截留的物理过程以及微生物参与的生化代谢过程。其前者受到土壤结构、土壤水气状态等物理特性的影响。后者是指含有氧化亚氮还原酶基因(nosZ)的微生物将N2O还原为N2的过程,是土壤N2O消耗的唯一生物途径。土壤类型、含水量、土壤发育程度、有机碳含量、农作物生长等对这类土壤微生物具有深刻的影响效应,进而对土壤N2O的消耗潜力也会产生重要的影响。例如,研究发现不同类型土壤N2O的消减潜力差别很大,其中红壤对N2O的消耗速率高达3.7 mg·kg-1·h-1[4]。淹水或厌氧的环境有利于土壤N2O的吸收,水稻田土壤具有很强的N2O消耗能力[5]。然而,不同发育程度的湿地土壤N2O消耗潜力及影响要素还缺乏深入了解。
随着我国区域经济的发展,对土地的需求越来越迫切,滩涂围垦改农田成为解决土地问题的有效途径[6]。崇明岛东滩是中国长江口规模最大、发育最完善的河口型滩涂湿地,在中华人民共和国成立后经历过多次大规模的围垦造田工程[7]。围垦后的湿地不再受潮汐影响,随后的农作物种植、翻耕、施肥等农业管理措施,形成了一系列不同熟土化进程的围垦区湿地。不同围垦年代的湿地土壤,代表了土壤发育的不同时期。其土壤母质本底相对一致、具有完善的年代发育序列,其土壤发育的空间变化可以反映时间尺度上的变化。许多研究发现,随着围垦年限增加、熟土化进程的推进,湿地土壤的盐分含量下降、pH值降低、SO42-浓度减小、有机碳含量增加,表现为土壤综合肥力增加、可耕性增强[8-10]。在土壤氮转化方面,李玲等[11]发现,闽江河口湿地围垦后表层沉积物N2O的产生潜力没有发生明显变化。李小飞等[12]认为,崇明东滩湿地中低潮滩土壤的硝酸盐还原潜力较高,可能与土壤可溶性有机碳密切相关。林黎等[13]则发现,河口湿地围垦后土壤理化性质的变化强烈影响着土壤微生物的群落结构和数量。郭俊丽[14]发现,外源N2O添加可使不同母质土壤的N2O消耗速率提高,显著提高N2排放量。那么,在滨海滩涂围垦改稻田这一熟土化过程中,不同发育进程的湿地土壤N2O消耗潜力如何变化?以及化学氮肥施用对其有何影响?这些科学问题的探索对于深入了解农田土壤N2O消耗潜力的演替特征至关重要。
基于此,本研究以长江口崇明岛东滩自然湿地和围垦后的稻田土壤为研究对象,以空间代替时间的方法,研究外源氮对不同围垦年限土壤N2O还原过程的影响,阐明土壤发育进程对湿地土壤N2O消耗潜力的影响特征,为我国农田温室气体减排提供科学依据。
1 材料与方法
1.1 土壤样品采集
供试土壤采自上海市崇明岛东滩湿地和围垦区稻田(31°27′~31°51′N,121°09′~121°54′E),该地区属于亚热带季风气候,年均气温为15.3℃,年均降水量为1003.7 mm。本研究选取东滩湿地保护区的高滩位芦苇湿地,作为围垦0年的对照样点,以LW表示。根据Cui等[15]的文献说明,在围垦区内选取围垦19、27、51、86年的稻田土壤作为不同围垦年限的稻田样点,分别记作WK19、WK27、WK51、WK86。采样时,在每个样区以S形走线设置6个采样点,每个点间隔10~15 m,然后用土钻取0~10 cm表层鲜土,将该6个点的土壤均匀混合成1个混合样本,重复3次。将土样装入密封袋,放入冰盒中运回实验室,于4℃下保存。
1.2 土壤理化性质测定
将采回的土壤样品进行本底理化性质的分析测定。土壤有机碳(SOC)用浓硫酸-重铬酸钾消煮、硫酸亚铁滴定法测定。土壤全氮(TN)用凯氏定氮法测定。土壤铵态氮(NH4+-N)和硝态氮(NO3--N)含量采用2 mol·L-1KCl溶液浸提后,AA3流动分析仪测定。土壤pH以水土比2.5∶1混匀静置后,用数字酸度计测定。土壤电导率(EC)以水土比5∶1浸提溶液用电导率仪测定。土壤硫酸根(SO42-)使用离子色谱法测定。
1.3 试验处理及培养
选取上述5种不同围垦年限的土壤,研究4个不同形态氮添加处理下湿地土壤的N2O还原速率。外源氮添加处理分别为(1)添加NH4+-N,(2)添加NO3--N,(3)添加尿素,氮最终浓度均为5 mmol·L-1;以及不添加氮(添加去离子水)的对照处理,每个处理重复3次。采用厌氧培养方法分析土壤的N2O还原速率。具体步骤为:取2.00 g鲜土放入12 mL的Labco顶空瓶中,加入2 mL去离子水,抽真空-充氩气重复进行3次。室温25℃避光预培养3 d;预培养结束后再进行抽真空-充氩气操作3次,向培养瓶中打入2 mL N2O气体,再加入20 μL氮溶液,使培养瓶中氮最终浓度为5 mmol·L-1。充分摇晃形成均匀泥浆,分别在加入底物后第0、2、4、7、14和21 d,测定培养瓶中的顶空N2O浓度。
1.4 土壤N2O还原速率测定
每次采气前充分摇晃培养瓶,用微量采样器采集100 μL顶空气体转入12 mL的Labco顶空瓶中,采集之后补充相同量高纯氩气以保持培养瓶内气压的平衡。采集的气体用气相色谱仪(Agilent 7890B)分析N2O浓度,仪器参数为:前检测器温度250℃,H2、空气和尾吹气N2流量分别为50、450和25 mL·min-1,后检测器温度为250 ℃,尾吹气氩甲烷流量为2.000 mL·min-1,柱箱温度为50 ℃,镍转化炉温度为375 ℃。土壤N2O还原速率通过分析培养期间瓶内N2O浓度的变化情况进行计算,计算公式如下:
式中:P是N2O的还原速率,μg·g-1·d-1;dc/dt为培养期间培养瓶上部气相N2O浓度变化率,μL·L-1·d-1;V为培养瓶上部气体体积,L;W为培养的鲜土质量,g;MW为N2O摩尔质量,g·mol-1;MV为标准状态下气体摩尔体积,22.4 L·mol-1。
1.5 数据处理分析
试验数据用SPSS 22.0和Excel 2010进行统计分析,其中显著性差异用单因素方差分析和LSD检验;Pearson相关性分析检验土壤N2O还原速率与土壤理化性质之间的关系。
2 结果与分析
2.1 土壤理化性质变化特征
芦苇湿地(LW)的土壤有机碳(SOC)含量为13.53 g·kg-1,围垦稻田SOC含量范围为9.55~18.34 g·kg-1(表1)。而 滩 涂 湿 地WK19土壤SOC含量下降明显,比LW湿地显著降低了29.42%(P<0.05);其后随着围垦年限增长SOC显 著 增 加(P<0.05),表 现 为WK27、WK51和WK86均显著高于LW(P<0.05)。湿地土壤总氮(TN)含量范围为0.77~1.20 g·kg-1,其中LW样点的土壤TN显著高 于WK19、WK27和WK51土壤(P<0.05),而与WK86则无显著差异。土壤铵态氮(NH4+-N)和硝态氮(NO3--N)含量分别为5.46~23.37和8.86~18.67 mg·kg-1,WK51土壤NH4+-N和NO3--N含量均显著高于其他样点土壤(P<0.05),而WK19土壤则显著低于其他土壤(P<0.05)。WK土 壤pH值 显 著 低 于LW土 壤(P<0.05),且随围垦年限增加,pH值下降。LW土壤的电导率(EC)为4.80 mS·cm-1,围垦后EC显著降低62%~85%(P<0.05),且随围垦年限增加而显著降低(P<0.05)。WK土壤硫酸盐(SO42-)含量范围为196.07~328.10 mg·kg-1,且随围垦年限增加而降低,相对于LW土壤,WK土壤的SO42-显著降低43%~66%(P<0.05)。
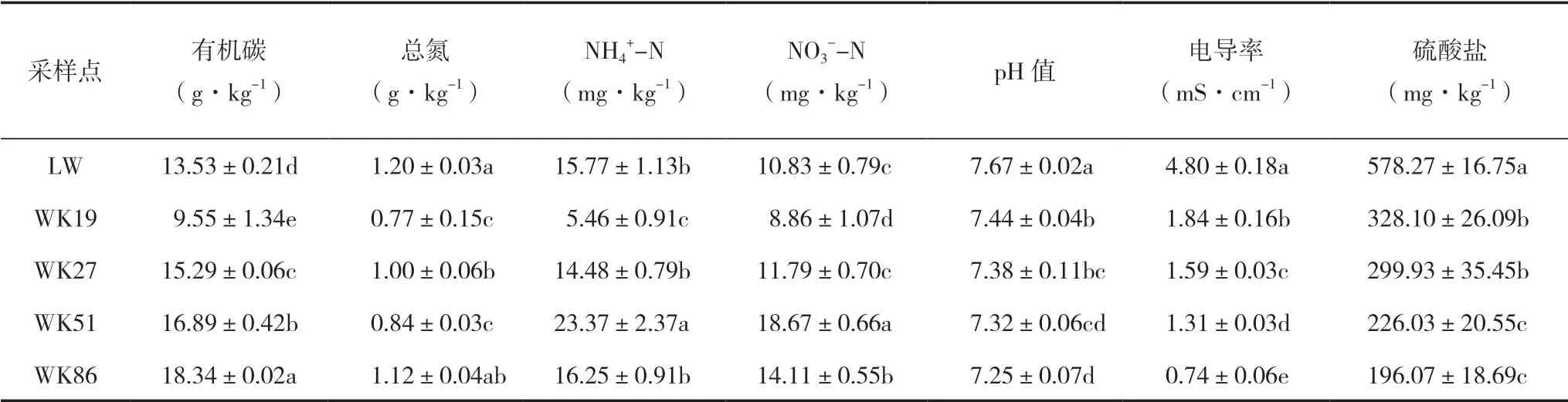
表1 不同围垦年限土壤理化性质
2.2 培养过程中N2O浓度变化特征
高浓度的N2O底物供应可反映出所试土壤的最大N2O还原潜力。培养起始各处理的顶空N2O浓度均高于850 μg·g-1(图1)。随着厌氧培养进行N2O浓度均呈现明显下降的趋势。总体而言,在整个培养过程中第0~2 d N2O浓度下降最为明显,之后第4~21 d保持较为稳定,直至培养结束,说明培养期间的前2 d各土壤样品表现出最大消耗潜力。此外,无氮对照CK处理中,第2 d后LW土壤的N2O浓度明显低于其他4种土壤。添加NO3--N处理中,第2 d后WK19和WK27土壤的N2O浓度明显高于其他3种土壤。

图1 不同氮添加处理后N2O浓度随厌氧培养时间的变化规律
2.3 湿地土壤的N2O还原速率
根据上述结果,选取第0~2 d的N2O浓度变化计算出湿地土壤的N2O还原潜力。发现CK处理及添加NH4+-N、NO3--N和尿素处理下,N2O还原速率 分 别 为18.0~36.6、11.5~39.5、15.2~59.5和17.2~38.9 μg·g-1·d-1(图2)。无氮CK 处理下,围垦稻田土壤N2O还原速率分别为18.0、26.0、25.3和31.2 μg·g-1·d-1,芦苇湿地土壤N2O还原速率显著高于围垦稻田土壤,是围垦稻田土壤的1.2~2.0倍(P<0.05)。添加NH4+-N处理下,WK86土壤的N2O还原速率显著高于其他土壤(P<0.05),WK19土壤显著低于LW土壤的N2O还原速率(P<0.05),而WK27、WK51土壤与LW土壤之间没有显著差异。添加NO3--N处理下,LW土壤的N2O还原 速率显著高于WK19、WK27和WK51土壤,WK86土壤的N2O还原速率比WK19土壤显著增加了291.2%(P<0.05)。添加尿素处理下,LW与WK51土壤的N2O还原速率分别为36.9和38.9 μg·g-1·d-1,两者之间没有显著差异,但显著高于其他3个样地土壤(P<0.05),而且WK86土壤的N2O还原速率最低,为17.2 μg·g-1·d-1。
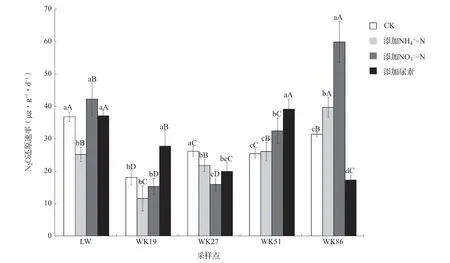
图2 氮添加处理下各土壤的N2O还原速率
对于每一个土壤而言,添加外源氮对N2O还原速率的效应各异。WK86土壤在添加氮后变化最为明显,其中NH4+-N和NO3--N处 理 下 的N2O还原速率相对无氮CK处理显著升高了27%和91%,而添加尿素处理下土壤N2O还原速率显著降低了45%(P<0.05)。氮添加处理均增加了WK51土壤的N2O还原速率,其中以添加尿素最为显著,提高了54.1%。相反,WK27土壤在氮添加条件下均显著下降,其中以NO3--N效应最为显著。对于LW和WK19土壤而言,添加NH4+-N处理显著抑制了LW土壤的N2O还原速率,而添加尿素处理则显著提高了WK19土壤的N2O还原速率。
2.4 回归分析及相关性分析
对稻田土壤的N2O还原速率与围垦年限进行了回归分析(图3)。结果表明,无氮CK处理以及添加NH4+-N、NO3--N处理下,土壤的N2O还原速率与围垦年限呈极显著的线性正相关(P<0.01),表明随着围垦年限的增长稻田土壤的N2O还原速率持续增加。其中添加NO3--N处理的N2O还原速率年增长率最高,为0.69 μg·g-1·d-1·年-1,添加NH4+-N处理的年增长率为0.37 μg·g-1·d-1·年-1,分别比CK处理高出343%和140%。相反,添加尿素处理下稻田土壤的N2O还原速率虽随围垦年限增加呈下降趋势,但未达到显著水平。
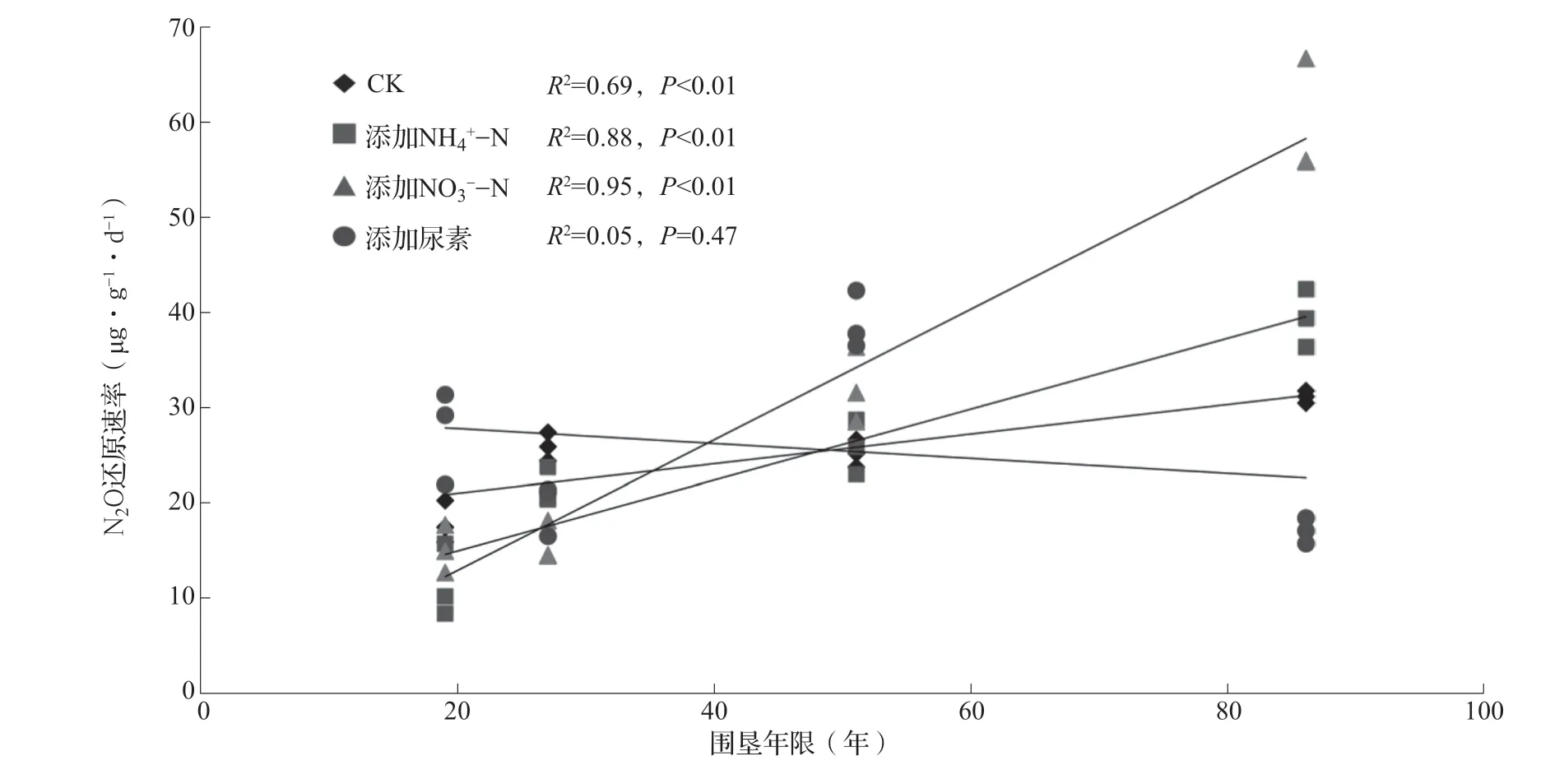
图3 土壤N2O还原速率与围垦年限的关系
将稻田N2O还原速率与土壤理化性质进行相关性分析,结果见表2。CK、添加NH4+-N和NO3--N处理下,稻田N2O还原速率与SOC、TN均呈显著(或极显著)正相关。相反,CK、添加NH4+-N和NO3--N处理下,稻田N2O还原速率与土壤EC、SO42-均呈显著(或极显著)负相关。土壤NH4+-N含量与稻田N2O还原速率在无氮CK和NH4+-N添加条件下呈显著正相关,土壤pH值与稻田N2O还原速率在添加NH4+-N和NO3--N条件下呈显著负相关。值得注意的是,添加尿素条件下稻田N2O还原速率与土壤理化性质的相关性均未达到显著水平。
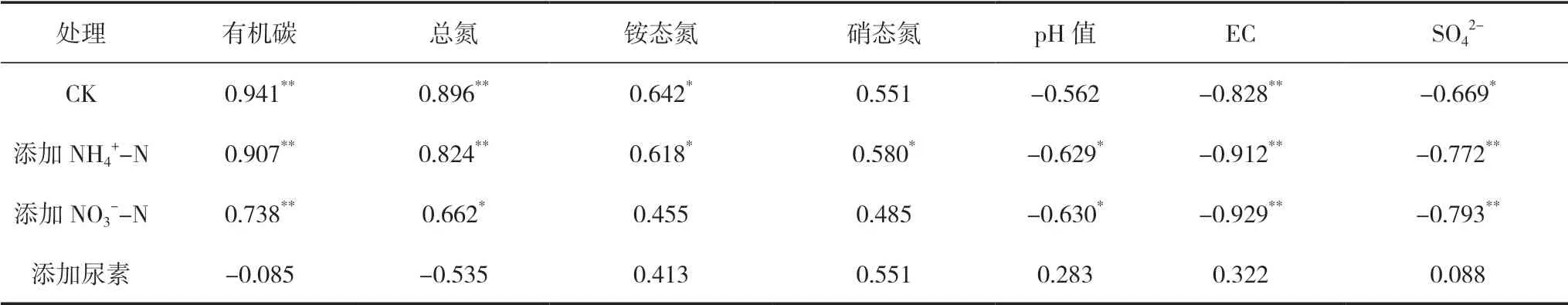
表2 围垦稻田N2O还原速率与土壤理化性质相关分析
3 讨论
3.1 滨海湿地土壤理化性质的变化特征
大规模围垦活动下,滨海滩涂湿地受到干扰,会深刻影响湿地土壤的理化性质[16]。本研究中,芦苇湿地SOC相较于围垦19年稻田土壤高出42%。Davidson等[17]认为由于湿地的大部分有机碳没有被土壤固定,在向陆地性农田转变的过程中,好氧条件增强导致有机碳的损失。随后由于盐分逐渐下降开始种植农作物、施肥和农作物残渣以及耕作管理措施使有机碳开始恢复[10,18],Mitra等[19]和Iost等[20]也有同样的报道。本研究发现,围垦86年SOC含量最高,且该稻田SOC含量超过芦苇湿地碳水平的36%;而芦苇湿地土壤的TN含量是围垦稻田土壤的1.1~1.6倍,可能与芦苇湿地表层存在丰富的固氮微生物有关[21]。土壤pH值、EC和SO42-均表现为芦苇湿地显著高于围垦稻田,且随围垦年限增加而呈逐渐下降的趋势。这与以往的研究报道一致[8-10,22],表明围垦后湿地土壤由海洋性特征向陆地性特征演变,将可能会对土壤N2O还原过程产生深刻影响。
3.2 滨海湿地土壤N2O还原潜力的特征比较
无氮添加处理下,湿地土壤的N2O还原速率反映了原状土壤的N2O消耗潜力。本研究发现,在围垦稻田中,86年稻田的N2O还原潜力最高,比19年稻田高出73.7%。相关性分析发现围垦稻田N2O还原速率与SOC呈极显著正相关。厌氧反硝化作用通常被认为是调控土壤N2O消耗还原的主要过程[23],在反硝化过程中,有效碳源的供应能够促进土壤反硝化微生物的生长,从而促进对N2O的吸收量显著增加[24]。而且,SOC激活了微生物的呼吸作用,加快了土壤中O2的消耗,从而形成厌氧环境,促进N2O还原成N2。此外,当土壤中NO3--N含量很低时,N2O就可能成为反硝化作用的唯一电子受体而被还原为N2。而在围垦86年稻田中的NO3--N含量显著高于19年稻田,并不有利于N2O的还原过程。事实上,针对有机碳和氮素对N2O还原的综合影响而言,有机碳与硝态氮的比值可能更好地反映出两者对N2O还原过程的整体影响效应。本试验中围垦86年稻田的有机碳与硝态氮的比值高于围垦19年稻田,可能是N2O还原速率存在差异的重要原因。不仅如此,土壤中SO42-作为重要的电子受体进行SO42-还原作用,将与N2O还原过程产生强烈的竞争作用[25]。而围垦86年稻田中最低的SO42-浓度极大地降低了对N2O还原过程的底物电子竞争,有利于N2O被还原为N2。
值得注意的是,本研究发现芦苇湿地的N2O还原速率显著高于围垦稻田。这可能与芦苇湿地处于低潮滩区域,土壤含水量较高,长期处于极度厌氧状态有关。以往研究报道土壤处于极度缺氧的环境中,N2O还原酶活性较高,有利于N2O还原成N2[26]。而且,芦苇湿地的pH值较高,有报道认为偏碱性pH值较高的土壤中N2O还原酶活性较强,促进N2O还原为N2[27]。此外,芦苇生长期根系分泌到土壤中的活性有机碳可能不同于水稻根系,导致N2O还原微生物的活性产生差异。聂文婷[28]研究发现,添加葡萄糖和蔗糖会增加N2O的排放量,而添加柠檬酸则会降低微生物活性,使N2O排放量低于其他碳源。
在添加不同形态外源氮后,各个土壤的N2O还原潜力发生改变。其中,NO3--N对围垦稻田N2O还原速率的促进效应明显高于NH4+-N,且在围垦86年稻田土壤中表现最为突出。添加NO3--N条件下,NO3--N可作为底物促进反硝化微生物活性,包括N2O还原微生物,可进一步促进N2O被还原。汪方圆等[29]对本试验区域进行研究,发现nosZI基因是围垦稻田土壤N2O还原过程的主要贡献者;而该基因存在于典型的反硝化型微生物中。因此,添加NO3--N可促进反硝化过程各个反应步骤,同时也有利于nosZI基因介导的N2O还原过程。同时也发现,在围垦19、27年稻田中添加NO3--N后,N2O还原速率均有不同程度地降低。这可能是因为该稻田土壤中有机碳较低,提供的电子较少,导致外源NO3--N与N2O争夺电子,进而N2O还原过程被抑制。另外,NH4+-N是大多数微生物氮源吸收的偏好形态。当土壤中添加NH4+-N时,会产生较强的微生物吸收固持作用,导致参与硝化和反硝化作用的底物氮素减少,这可能是导致添加NH4+-N对N2O促进效应低于NO3--N的重要原因。尽管添加NO3--N对稻田土壤N2O还原速率的正效应最高,但它本身是阴离子,不易被土壤胶体吸附而随水流失;NH4+-N的正效应虽然低于NO3--N,但是容易被土壤胶体吸附,易被土壤微生物吸收利用。从氮素利用率和生态效应两个方面综合比较而言,施加NH4+-N对土壤N2O还原速率的综合效应优于NO3--N。需要指出的是,本试验采样厌氧培养试验研究不同外源氮输入对土壤N2O还原潜力的影响效应,没有对相关微生物的功能基因进行分析。在此基础上,可进一步设置培养试验研究功能基因nosZ的不同亚群在N2O还原过程中的相对贡献,揭示其微生物机制,从而建立土壤N2O还原潜力与反硝化微生物丰度和群落结构的内在关系。
4 结论
滨海湿地围垦改为稻田后,显著降低了土壤pH值、EC值和SO42-含量。无外源氮添加条件下芦苇湿地土壤N2O还原速率显著高于围垦稻田土壤。添加NH4+-N和NO3--N极大提高了围垦86年稻田土壤N2O的还原速率。
致谢:在本试验的野外采样过程中,上海崇明东滩湿地自然保护区给与了大力支持和协助,在此表示衷心感谢!
