狭义蛇葡萄属东亚支系的系统发生与生物地理演化
2022-10-02何关浩WENJun聂泽龙
王 达,何关浩,孟 然,孟 盈,WEN Jun,聂泽龙*
(1 吉首大学 生物资源与环境科学学院,植物资源保护与利用重点实验室,湖南吉首 416000;2 Department of Botany, National Museum of Natural History, Smithsonian Institution, Washington, DC, 20013-7012, U.S.A.)
葡萄科(Vitaceae)隶属于双子叶植物纲蔷薇亚纲,包括16属近1 000种,主要分布在亚洲、非洲、以及北美洲的热带地区,少数属分布在温带地区[1-2]。葡萄科植物多为木质藤本,其卷须与叶对生,是该科的重要形态特征,卷须是在花序基础上进化而来,以提高该科植物的攀附能力,加强对各种生态环境的适应能力[3]。近年来系统学工作者对葡萄科属的界定、属内分类以及生物地理演化开展了大量研究[4-7]。Soejima和Wen[8]首次使用3个叶绿体数据构建了葡萄科5个分支的整体系统框架,随后基于多个核基因和叶绿体标记的系统发育分析也得到了类似结果[4]。基于下一代高通量测序的基因组数据进一步支持将葡萄科分成5个族[2]:即蛇葡萄族(Ampelopsideae)、爬山虎族(Parthenocisseae)、葡萄族(Viteae)、白粉藤族(Cisseae)和乌蔹莓族(Cayratieae)。蛇葡萄族是葡萄科分化出的第一支,族下包括狭义蛇葡萄属(Ampelopsiss. str.)、羽叶蛇葡萄属(NekemiasRaf.)、RhoicissusPlanch.和ClematicissusPlanch.等4个属[7]。
原蛇葡萄属最初于1803年由André Michaux提出,只包括了A.bipinnata、A.cordata和A.quinquefolia等3个种[9]。Bunge根据卷须形态和花瓣特征发表了A.aconitifolia、A.humulifolia和A.serianaefolia等3个东亚地区的物种[10]。Planchon对原蛇葡萄属分类学地位和形态特征进行了研究,将原蛇葡萄属作为属的地位归于葡萄科下,属内分为14个种,进一步界定了各个种的形态特征和分布范围[11]。Baileri在前面的基础上对原蛇葡萄属的卷须、叶序、花序、子房以及花盘的形态进行了更详细的描述,认为原蛇葡萄属在形态上与葡萄属、爬山虎属和白粉藤属有明显区别,并对属下进行了更详细分类。根据叶序分为单叶组、掌状叶组和羽状叶组,在此基础上,还对单叶组叶下表面的颜色及是否存在绒毛进行了更详细的鉴定,对羽状叶组的叶缘锯齿进行了详细鉴定。在此分类基础上定义了原蛇葡萄属的各个种之间的形态差异,对后来的原蛇葡萄属分类提供了重要参考[12]。Galet基于先前的研究基础对原蛇葡萄属的叶序形态研究认为,原蛇葡萄属下分为2个组:羽状复叶组Leeaceifoliae和单叶或掌状复叶组Ampelopsis[13]。Bernard对原蛇葡萄属的腋芽形态进行了鉴定,结果表明Leeaceifoliae组的腋芽形态和葡萄属的Vitisvinifera相似,具有复杂的腋芽模式,而Ampelopsis组的腋芽为连续分布模式[14]。
原蛇葡萄属的分类地位得到了上述多位分类学家的支持,但是其属内的关系仍然存在一定的争议,传统分类学并没有很好地解决这个问题,直到基于分子数据的系统学研究出现,才有效地界定了蛇葡萄属内各个种的关系。基于3个叶绿体的系统发育分析,发现原蛇葡萄属不是一个单系类群,来自非洲的Rhoicissus的2个种与南美的Cissusstriata复合群嵌套于原蛇葡萄属中,并与原蛇葡萄属的Ampelopsis组构成一支,而Leeaceifoliae组单独构成另外一支,这两支互为姊妹类群[8]。Wen利用核基因GAI1片段的系统发育分析得到了类似结果,不同之处在于Rhoicissus的2个种与Cissusstriata构成的复合群和Leeaceifoliae组关系更近[1]。基于4个叶绿体基因联合数据对原蛇葡萄属进行系统发育分析得出的结果也支持将原蛇葡萄属分为两支,分别对应Leeaceifoliae组和Ampelopsis组[15]。基于先前的分子数据,Wen将Leeaceifoliae组的9个种及1个变种分离出去构成羽叶蛇葡萄属(NekemiasRaf.),Ampelopsis组构成现在的狭义蛇葡萄属(Ampelopsiss. str.)[16]。基于转录组数据的系统发育分析和基于Hyb-seq测序数据的系统发育分析均支持将羽叶蛇葡萄属从原蛇葡萄属中分离[17-18]。
狭义蛇葡萄属为落叶木质藤本,是葡萄科中少数分布在温带地区的类群,总共约15~18个种,除了北美2种和欧洲3种以外,其余种类都分布在东亚,呈典型的北温带间断分布模式[19]。蛇葡萄属卷须通常2~3分叉,末端无吸盘(北美的A.cordata除外),叶通常为单叶或掌状复叶,果实通常为近球型,单果种子1~4颗,种子在被子植物中是独特的,其腹部具有1对皱褶,背部有1个凸起的合点[20-21]。基于4个叶绿体标记的系统发育分析在族水平上对狭义蛇葡萄属的系统发育和生物地理演化进行了进一步的研究,其系统发育分析结果支持北美的A.cordata和A.denudata位于狭义蛇葡萄属的基部,所有东亚类群形成一支和欧洲的A.orientalis互为姊妹类群。在东亚支系中,蓝果蛇葡萄(A.bodinieri)和尖齿叶蛇葡萄(A.acutidentata)形成一支位于亚洲类群的基部,白蔹(A.japonica)单独位于亚洲分支的第二支,其他亚洲类群分成两小支嵌在亚洲分支的内部。和系统发育的结果相对应,其生物地理演化分析结果支持蛇葡萄族的祖先起源于始新世中期的北美,在渐新世沿北大西洋陆桥扩散至欧亚大陆[15]。
虽然狭义蛇葡萄属的系统关系和生物地理演化模式在先前的研究中得到了一定的阐明,但其并没有着重讨论狭义蛇葡萄属内的系统关系,且东亚类群内部的演化机制没有得到阐明。所以为了更好地了解狭义蛇葡萄属内部各个种的系统发育关系以及东亚类群的演化特点,还需要对其进行更完整的取样。本研究通过加强对东亚类群的取样,利用2个核基因(GAI1和ITS)和5个叶绿体标记(trnL-F、rps16、psbA-trnH、atpB-rbcL和trnK-petN),首先验证狭义蛇葡萄属的单系性,并重建狭义蛇葡萄属的系统发育框架,特别是探究狭义蛇葡萄属东亚支系的系统演化关系与生物地理演化特点,为进一步研究它们在北半球的生物地理进化历史提供基础。
1 材料和方法
1.1 材料选择与分子测序
本研究参考先前研究中狭义蛇葡萄属及外类群的界定结果,选取了狭义蛇葡萄属内12个种和3个变种的37个样本(表1),对核基因GAI1和ITS、叶绿体基因rps16及叶绿体基因间隔序列(psbA-trnH、atpB-rbcL、trnK-petN和trnL-F)进行测序。取样涵盖了大多数现存蛇葡萄属的类群,能很好地代表了狭义蛇葡萄属各个种的地理多样性和形态多样性。为了更为全面的取样和综合利用目前已有的数据,从GenBank上获取缺失或取样较少的狭义蛇葡萄属5个种的7个样本,以及同族的外类群Rhoicissus(1种),Clematicissus(1种)及羽叶蛇葡萄属(5个种)的上述序列的数据(表2)。
本研究所用的狭义蛇葡萄属植物材料源于野外采集或苗圃培养,凭证标本现存于中国吉首大学植物标本馆和美国国家标本馆。用改进的CTAB法[22]从硅胶干叶或植物标本中提取总DNA。叶绿体基因扩增和测序参照Soejima和Wen[8]和Liu等[23]对rps16、trnL-F、trnC-petN、atpB-rbcL和psbA-trnH的处理方法,核基因的GAI1和ITS参照Liu等[24]的方法。
1.2 系统发育分析
在获得整理好的数据后,先使用Muscle 3.8.31[25]分别对每个分子标记进行比对,辅之手动微调。采用最大似然法[26]和贝叶斯推导法[27-28]对2个核基因数据(ITS和GAI1)和5个叶绿体片段数据(trnL-F、aptB-rbcL、trnK-petN、psbA-trnH和rps16)单独进行系统发育分析。参照先前研究处理方法[29-30],将核基因与叶绿体基因进行联合处理,使用PAUP 4.0[31]对叶绿体与核基因数据进行一致性检验,结果为P=0.33,支持将叶绿体数据和核基因数据进行联合分析。
首先用RAxML 7.2.6[32-33]推导最大似然树。所有联合分析的数据被划分成对应分子片段分区,允许对每个分区进行独立参数估计。所有的分析对每个分区都采用了GTRGAMMA模型。用bootstrap=1 000次对进化支进行置信度评估。贝叶斯分析用MrBayes[34],用MrModeltest2.3[35]检验确定的核苷酸最佳替代模型,并且对联合数据进行分区处理,每个分区采用独立参数估计。贝叶斯分析从随机树开始,采用4 条链1 000万代的马尔可夫链-蒙特卡罗算法,每1 000代保留一棵树,舍弃初始样本(burn in=2 500)后,根据剩余的样本构建一致性综合树。重复运算2次以检验其收敛性,最后将后验概率PP>95%的分支视为可信度高。
1.3 形态性状演化分析
叶形态性状是狭义蛇葡萄属物种的重要鉴定依据,参考Baileri[12]对狭义蛇葡萄属叶形态的研究利用Mesquite 3.51的MK1进化模型,对狭义蛇葡萄属的叶序、叶表面是否有绒毛以及叶缘锯齿的形态3组形态性状进行演化分析。叶序分为单叶不裂或浅裂、单叶中裂或深裂、掌状复叶3种状态。叶表面的绒毛特征分为叶两面无绒毛、叶下表面有疏绒毛、两面均有疏绒毛3种状态;叶缘的锯齿形态分为叶缘粗锯齿或叶缘细锯齿2种模式。将以上数据进行赋值后,映射到联合数据构建的贝叶斯上进行分析。

表1 用于狭义蛇葡萄属系统发育分析的样本信息
1.4 生物地理研究
利用Beast2对狭义蛇葡萄的分化时间进行估算。另外参考Wen等[2]研究结果,从GenBank中收集了葡萄科的外类群火筒树科的一个种,葡萄科下的其他属的6个种的序列(表2)并结合上述使用的联合数据构建最大似然树用于时间估算。参照Magallón等的结果[36],将葡萄科根部校准为90.65百万年前,并选用Nie等[15]的分歧时间估算结果,将蛇葡萄族根部进行校准为(41.2±4)百万年前,时间标定均选用Normal计算模型。利用松散分子钟模型,tree prior选用Yule Model,以随机数作为起始,4条马尔科夫链(MCMC)同时迭代计算20 000 000代,每1 000代进行一次取样,将开始的25%样本作为burn in舍弃。用Tracer v1.7.1 (https://beast.bio.ed.ac.uk/software/tracer/)检验结果的平稳性和收敛性。
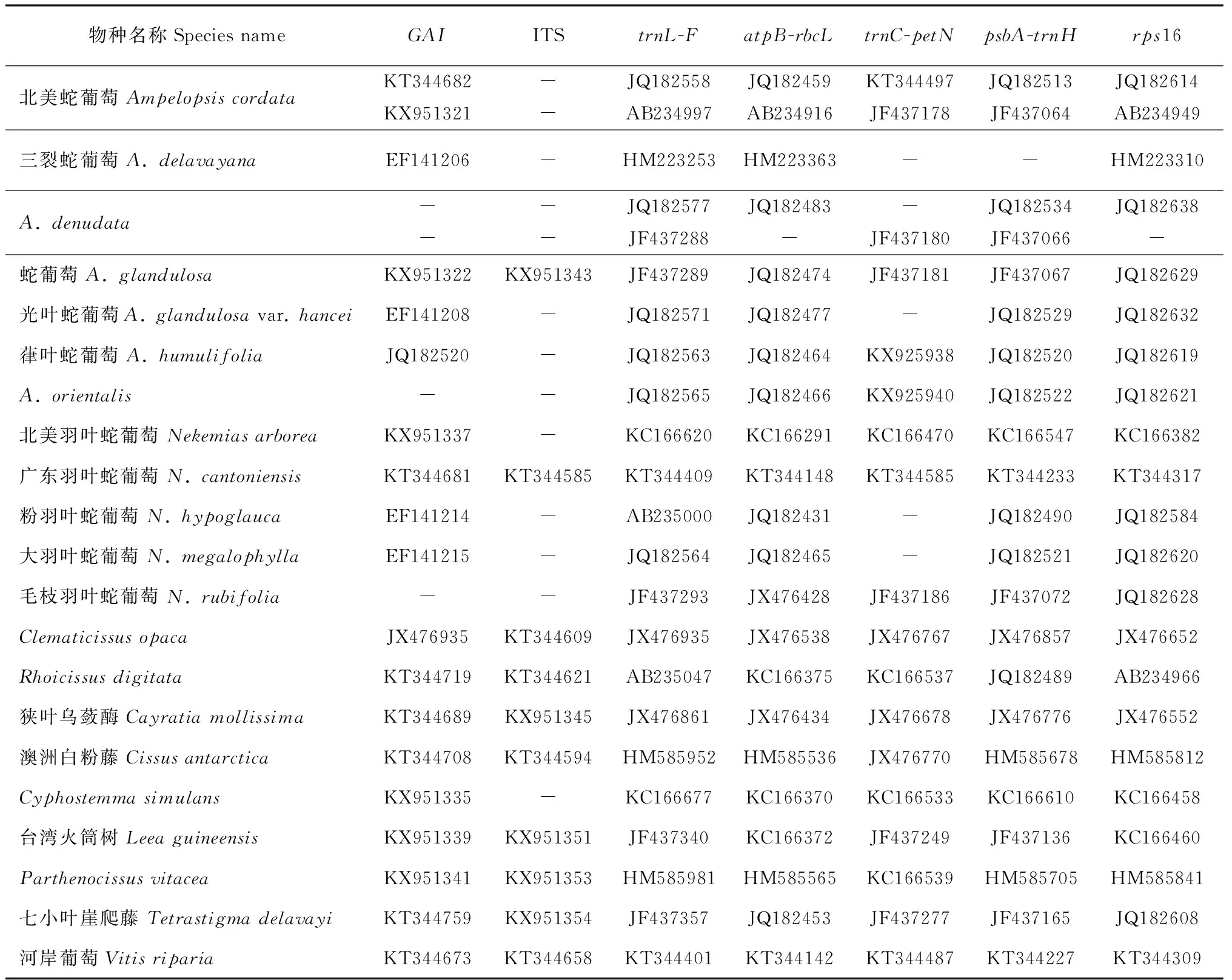
表2 从GenBank上收集的狭义蛇葡萄属及其外类群系统发育分析的序列信息
使用RASP[37]重建狭义蛇葡萄属的祖先地理分布,参数设置参考Ma等[18]的处理方法,使用BBM(Bayesian Binary MCMC)独立分析2次,每次运行10条马尔科夫MCMC链,同时进行随机取样,每条MCMC链运行100 000个循环,温度设定为0.1以保证各个链具有合适的活跃度。根据狭义蛇葡萄属以及其近缘类群的分布特点,生物地理分析的分区划分为:北美(A)、欧洲及西亚(B)、东亚(C)、澳洲(D)和非洲(E)。根据现有类群的分布特点,祖先分布区范围的最大分布地被限制为2个。
2 结果与分析
2.1 系统发育
分子序列特征参数如表3所示,在比对好的核基因序列中,GAI1、ITS长度分别为1 407和800 bp,叶绿体基因序列rps16为800 bp,叶绿体基因内含子区域trnL-F、trnC-petN、psbA-trnH、atpB-rbcL长度分别为989、887、501和784 bp。其中核基因序列的变异相对较高,叶绿体的相对较低(表3)。

表3 狭义蛇葡萄属2个核基因和5个叶绿体分子数据的数据集特征
系统发育分析结果显示,最大似然树和贝叶斯树的拓扑结构基本一致,贝叶斯树总体的支持率较高且具有更好的代表性。在系统发育结果中,叶绿体基因树(图1,A)、核基因树(图1,B)、联合树(图2)的拓扑结构都强有力地支持狭义蛇葡萄属是一个单系类群(PP1A=1.00,LB1A=82%,PP1B=1.00,LB1B=100%)。来自新世界的A.denudata和北美蛇葡萄分别为狭义蛇葡萄属的第一支(PP1A=1.00,LB1A=100%,PP1B=1.00,LB1B=100%)和第二支(PP1A=1.00,LB1A=98%,PP1B=1.00,LB1B=100%)。叶绿体基因树和联合树都支持所有的亚洲类群形成一个分支(PP1A=1.00,LB1A=86%),并且和欧洲的A.orientalis互为姊妹类群(PP1A=1.00,LB1A=96%)。尽管未能收集到A.orientalis的核基因数据,但是叶绿体基因树和联合树的拓扑结构仍然能很好地支持本研究结果。在核基因树(图1,B)中,整个东亚类群的关系都是模糊的,白蔹位于亚洲类群的基部(PP1B=100%,LB1B=100%),其他的东亚类群之间的系统关系都没有得到解决。叶绿体树和联合树中有相对较好的拓扑结构,叶绿体树(图1,A)的拓扑结构支持将东亚的类群主要分为分支 1(PP1A=0.99)和分支 2两大支(PP1A=0.69)。联合树与叶绿体树有着相近的拓扑结构,但是叶绿体树分辨率更高,所以我们讨论中主要参考叶绿体数据的结果。
2.2 形态演化
祖先特征重建结果(图3)揭示掌状复叶是狭义蛇葡萄属的祖征,单叶是衍征;下表面细微绒毛是祖征,叶两面无毛和两面均有细微绒毛是衍征;叶缘锯齿粗细的祖征在数据中并没有很好的验证。形态性状分析表明所有性状在本属内都经历了多次独立的演化。
2.3 生物地理
狭义蛇葡萄属的分歧时间结果如图4所示,北美类群和欧亚类群的分歧时间为26.01百万年前,欧洲类群和东亚类群分歧时间19.32百万年前,东亚类群内部的两支的分歧时间为13.87百万年前。RASP分析结果表明狭义蛇葡萄属的祖先的分布区域在北美,大约在渐新世晚期迁移扩散到欧洲,中新世早期到达东亚并进行快速分化(图5)。
3 讨 论
3.1 系统发生关系
通过对狭义蛇葡萄属5个叶绿体和2个核基因分子数据的独立分析,以及叶绿体数据和核基因数据联合分析重建了狭义蛇葡萄属的系统发育框架,其结果均支持狭义蛇葡萄属的单系性。这与基于3个叶绿体数据的研究结果相一致[8],也与综合核基因以及基因组数据的系统发育结果一致[17-18]。狭义蛇葡萄属分为3个主要的进化支系,首先分化出来的为来自墨西哥的A.denudata,其次是来自北美的北美蛇葡萄(A.cordata),最后是来自欧洲的A.orientalis与所有来自东亚的类群组成的一大支互为姊妹类群。这个结果与该属在北美、欧洲和亚洲的地理分布模式相吻合,进一步印证了先前研究结果中认为狭义蛇葡萄属起源于北美的结论[15]。
与之前的分子数据结果一致[15],叶绿体分子数据和核基因数据都强烈支持东亚类群单独构成一支。但可能是由于核基因的信息位点不足或者狭义蛇葡萄属早期存在基因渗透,导致了核基因树中东亚类群内部各个支的分辨率不高,各个类群互相嵌在一起,因此目前核基因数据对狭义蛇葡萄属东亚类群系统关系不具备很好的参考价值。而叶绿体数据的结果则相对较好地将东亚类群分为两大支。分支1的蛇葡萄(A.glandulosa)和三裂蛇葡萄(A.delavayana)多个样本互相嵌在一起,可能是这两个种之间存在一个长期的杂交进化关系所造成的。这两个种均在中国东南至西南部广泛分布,高度重叠的分布范围为两个种之间的杂交提供了先决条件。且其花期均为8~10月,花的形态、结构都存在高度的相似性[20]。综合分子数据结果和上述条件,推测这两个种间存在较大的杂交渗透可能性。分支2中,蓝果蛇葡萄、异叶蛇葡萄(A.glandulosavar.heterophylla)以及尖齿蛇葡萄有着更近的系统关系,但是它们各自的系统关系不清楚,这3个种之间的亲缘关系仍需要进一步研究。另外,东亚支系两个分支呈现了一个明显的地理分布梯度,其中分支1的11个样本主要分布于中国偏西南的地区,分支2的样本主要分布于中国东南至华北地区。东亚支系两个主要演化支之间的分布重叠边界主要在湖南、湖北和陕西等地,大致符合东亚植物区系内中国-日本森林亚区和中国-喜马拉雅森林亚区的分界线[38]。中国-日本森林亚区从白垩纪以来,没有经历过大规模的地质环境改变,水平分布明显,受湿润的东亚季风影响,生态环境相对稳定。中国-喜马拉雅森林亚区地形复杂,且存在较多的天然避难所,地形垂直变化大,板块运动引起了地形持续且快速上升,且该亚区气候条件复杂多变,同时受到印度季风和东亚季风的影响,从而导致了这两个森林亚界之间的植物区系的生境条件有着较大的差异[38],这两个亚区之间地形地貌和气候条件的差异可能是导致东亚分支内两小支分裂的重要因素。
3.2 叶形态性状演化的复杂性
在葡萄科内,叶的形态性状是一个重要的分类特征,是经典分类学对原蛇葡萄属内各个种界定的主要依据[39]。形态分析结果表明掌状复叶是狭义蛇葡萄属内叶形态的祖征,单叶浅/不裂至深裂为衍征。在北美类群中,掌状复叶(A.denudata)和单叶(北美蛇葡萄)分成了两支,复叶的祖先扩散至欧亚大陆。而在亚洲分支内部,掌状复叶的种和单叶的种混合分布,各个节点间的叶序形态经历了多次的独立进化事件。另外在叶表面的稀疏绒毛有或无以及叶缘为尖锐锯齿型或粗锯齿的形态演化树中,各个种的形态特征均没有与分子结果相对应。例如在叶绿体数据和联合数据表明东亚内部种间的系统关系呈现明显的地域性,与各个种之间的形态特征没有明显关系。这说明叶形态特征的变化可能与生境的不同或变化有关系,并没有反映它们内在的系统演化关系。叶形态在一定程度上受小生境的影响,具有一定的表型可塑性[40]。此外,叶形态还会受到温度、光照、水分等多个条件因素的影响,甚至可以通过改变环境迫使同一植株中同时存在两种或多种叶片形态,例如人鱼藻属(Proserpinaca)、毛茛属(Ranunculus)和水蓑衣属(Hygrophila)等属的植物叶的形态均有这种现象[41]。因此狭义蛇葡萄属内叶形态可能不是一个很好的分类学依据,还有待进一步的研究来揭示叶形态在狭义蛇葡萄属内的演化规律。
3.3 狭义蛇葡萄属的北美起源及在东亚的生物地理演化特点
早在第三纪大量新进化的植物类群在温暖的气候条件下于北半球快速蔓延[42],由于其位于北半球以及其多数类群都具有喜温性,Wolfe[43]将其命名为泛北热带植物群。相较于位于高纬度的白令陆桥,喜温植物更可能通过北大西洋陆桥在新旧世界之间扩散[44-45]。尽管渐新世晚期北大西洋陆桥已经断裂,但据先前的研究表明,北热带植物群仍然能通过陆桥断裂后留下的岛屿进行交流[46]。狭义蛇葡萄属在分布模式和扩散时间上均和上述扩散模式相吻合,所以我们推断其可能是通过当时泛北热带植物区系带,从北美通过北大西洋路桥扩散至欧洲,这也与先前的研究结果一致[15]。而在欧亚大陆,特提斯海的退却开放了欧洲和东亚之间的植物交流通道[47],狭义蛇葡萄属可能是在中新世早期沿着特提斯海退却后的通道扩散进入亚洲。当时喜马拉雅山脉和青藏高原还未完全抬升,在喜马拉雅山脉南麓还存在喜温植物森林带,这为欧洲和东亚之间的物种交流提供了可能性[48]。晚新生代的全球降温和喜马拉雅山脉的进一步抬升导致了欧亚植物交流通道关闭[48],至此狭义蛇葡萄属欧洲支系和东亚谱系分裂,形成了北半球间断分布格局。
狭义蛇葡萄属祖先在中新世早期抵达东亚后,经历了一个快速分化与适应性辐射进化的过程,在较短的地质时期(约1300万年)内物种多样性明显增加,使得东亚成为本属的分化中心。众所周知,适应性辐射进化是植物适应剧烈变化的地理环境和气候条件的一种有效机制[49],如中新世早期的地质运动和季风气候的加强导致了葡萄科的葡萄属(Vitis)的适应性辐射进化[18]。中新世早期印度洋板块和欧亚大陆板块持续碰撞引起了喜马拉雅山脉隆起和青藏高原抬升,并在中国西南部引起了多次造山运动,不仅造成了中国-喜马拉雅森林亚区和中国-日本森林亚区在地理环境上的差异,还进一步导致了东亚季风气候的加强[48]。持续的地质和气候的变化造成了中国-喜马拉雅森林亚区和中国-日本森林亚区之间生境异质性的差异[38]。地质环境的改变和季风气候的形成不仅提高了狭义蛇葡萄属的物种形成率,同时也可能导致了分布在中国-喜马拉雅森林亚区(分支1)和中国-日本森林亚区(分支2)的狭义蛇葡萄属两小支分离。
此外,东亚的山脉多为东西分布,这阻止了第四纪冰川向南迁移[50],有效地降低了冰川作用对东亚植物类群的影响,山脉之间的地理隔离和生态分化提供了适合物种形成的间断地[51],这些地理因素的影响进一步被中新世晚期的气候变化所放大[52]。晚新生代的全球变冷导致了整个北半球高纬度的狭义蛇葡萄属祖先灭绝,并驱使经历灭绝事件后孑遗的狭义蛇葡萄属祖先向南迁移,相比较于当时的欧洲和北美,东亚地区拥有更高的地理多样性,且具有更多的避难所和较低的灭绝率[53-54]。综上所述,狭义蛇葡萄属在东亚地区高的物种多样性是东亚地区地质事件和气候变化以及东亚地区的植被连续性共同作用所导致的。本研究在先前的基础上,探讨了狭义蛇葡萄属的北美起源,随后通过北大西洋路桥迁移至欧亚大陆,并进一步阐明了狭义蛇葡萄属从欧洲扩散至东亚的路径以及其在东亚地区高物种多样性的原因,研究结果可为北半球间断分布植物的生物地理研究以及其在东亚地区植物的适应性辐射进化研究提供参考。
4 结 论
狭义蛇葡萄属是葡萄科少有的北温带类群,具有重要的生物地理学研究意义。本研究结果支持狭义蛇葡萄属是一个单性系,A.denudata和北美蛇葡萄位于狭义蛇葡萄的第一支和第二支,这也与狭义蛇葡萄属起源于北美的理论吻合。所有东亚类群构成一支并与欧洲的A.orientalis互为姊妹类群。叶绿体数据揭示东亚类群内部主要分为两支,分支1内蛇葡萄和三裂蛇葡萄内存在长期的杂交进化关系,分支 2的蓝果蛇葡萄、异叶蛇葡萄以及尖齿蛇葡萄有着更近的系统关系。狭义蛇葡萄属的叶形态性状经历了多次独立进化,导致了叶的形态与系统进化没有明显的关系,形态变异可能与小生境多样化相关。生物地理研究揭示了狭义蛇葡萄属始新世中期起源于北美,渐新世晚期通过北大西洋路桥扩散至欧洲,中新世早期沿着特提斯海退却后的喜温植物森林带迁移扩散至东亚。东亚地区具有较高物种多样性,可能与其在中新世的高物种形成率和其受晚新生代冰川影响相对较小有关。但狭义蛇葡萄属在东亚地区的演化关系复杂,仍然需要进一步研究,特别是需要其基因组数据来进行更准确的系统发育分析。
