Rab蛋白参与结核分枝杆菌胞内吞噬的研究进展
2022-09-27孙巧玲
孙巧玲,袁 欣,秦 欢,王 玉
(遵义医科大学 基础医学院 微生物学教研室, 贵州 遵义 563003)
结核病(tuberculosis,TB)是由结核分枝杆菌(Mycobacteriumtuberculosis,M.tuberculosis,Mtb)引起的严重传染病。虽然已有疫苗进行预防接种,每年全球仍有超过1 000万新发结核病例,100余万人死于结核相关的疾病[1]。中国每年新增超过80万结核病病例。近年来由于多重耐药菌株的出现、艾滋病双重感染病例的增多以及免疫抑制剂的滥用导致结核病的发病率和死亡率不断上升。
Mtb主要经由呼吸道进入机体,到达肺部远端,随后被肺泡巨噬细胞或树突状细胞捕获,内化形成含Mtb吞噬体。对于普通细菌,细胞内吞噬体能与溶酶体结合形成吞噬溶酶体,再通过吞噬体酸化、水解酶、氧化酶作用等来清除细菌[2]。而Mtb能够通过其菌体成分干扰宿主细胞内吞噬作用,进而阻止吞噬体成熟,使得Mtb能够在细胞内存活和增殖[3]。
Rab蛋白属于小GTP酶家族,在真核细胞中广泛表达,调控了胞内各种膜转运过程,包括吞噬过程。大量研究表明Rab蛋白与结核分枝杆菌胞内吞噬密切相关。因此,本综述着重讨论影响胞内吞噬作用的Mtb菌体成分以及Rab蛋白对Mtb吞噬的影响,以期深入了解Mtb与Rab蛋白在胞内吞噬中的相互作用,探索Mtb的胞内免疫机制。
1 Mtb对于胞内吞噬过程的影响
吞噬是胞内抵抗病原菌感染的重要防御机制之一。吞噬过程包括了病原菌识别、吞噬体的形成、吞噬溶酶体成熟、病原菌降解4个阶段。Mtb首先被机体吞噬细胞如巨噬细胞表面Toll样受体(Toll-like receptors,TLRs)等识别后,内吞进入细胞内形成吞噬体,通过菌体成分干扰和阻止胞内吞噬,细菌能在胞内存活和增殖。由此可见,Mtb菌体成分与吞噬过程的相互作用决定了其胞内吞噬的结局。
1.1 Mtb脂质对胞内吞噬的影响
Mtb脂质是Mtb的主要毒性因子,与结核病的发生密切相关。Mtb存在大量脂质,如脂阿拉伯甘露聚糖(mannose-capped lipoarabinomannan,ManLAM)等。ManLAM广泛分布于Mtb表面,是一种复杂的脂聚糖,它能与甘露糖受体(mannose receptor,MR)结合,进而介导Mtb吞噬。而且ManLAM还能阻断吞噬体与溶酶体的融合,帮助细菌在胞内存活[4]。此外,有些Mtb脂类可通过调节ManLAM来影响胞内吞噬过程。P27是牛型结核分枝杆菌等表达的一种分泌型表面糖脂蛋白,因其能与P55共同协助Man-LAM组装和转运到细胞表面,故P27缺失时,可导致细菌表面Man-LAM表达减少,进而促进巨噬细胞内吞噬体的成熟[5]。此外,P27还能通过与DC-SIGN受体结合,介导Mtb与宿主间的黏附。Mtb细胞壁脂质成分PDIM(phthiocerol dimycocerosates)与Mtb胞内逃逸相关。PDIM可通过排出吞噬体膜中空泡质子ATP酶来阻断吞噬体的酸化,也能通过增加吞噬体膜通透性和造成膜损伤来帮助细菌存活和增殖[6]。
1.2 Mtb蛋白质对胞内吞噬的影响
目前已知结核分枝杆菌能够产生多种酶类,如蛋白激酶G(protein kinase G,PKnG)、酪氨酸激酶A(tyrosine kinase A,PtKA)等。PKnG类似于蛋白激酶,可通过增加胞内信号传导来阻止吞噬体和溶酶体的融合,干扰吞噬体成熟;另一方面,PKnG也具有泛素激活酶和泛素连接酶功能,能够调节肿瘤坏死因子受体相关因子2(tumor necrosis factor receptor-associated factor 2,TRAF2)和β-TGF活化激酶1(TGF-beta-activated kinase 1,TAK1)的泛素化和降解来抑制NF-κB通路激活,从而抑制机体免疫力[7]。PtKA缺失时,Mtb不能抑制吞噬体酸化,导致巨噬细胞内细菌死亡[8]。此外,Mtb H37Rv菌株产生的苹果酸合酶(malate synthase,MS)GlcB是一种帮助Mtb在缺氧环境中以甘油为碳源增殖时必需的酶类。当GlcB缺失时,Mtb与溶酶体标志物LAMP1共定位增加,细菌载量增加,提示GlcB能够阻止吞噬体成熟[9]。肌微管素相关蛋白4(myotubularin-related protein-4,MTMR4)是一种肌微管素-3-磷酸磷脂酰肌醇磷酸酶,表达减少时能够升高巨噬细胞表面免疫球蛋白受体FcγRs(Fc gamma receptors)的表达,并增加3-磷酸磷脂酰肌醇[phosphatidylinositol 3-phosphate,PtdIns(3)P]在吞噬体膜上持续的时间,从而抵抗海洋分枝杆菌诱导的吞噬体阻断作用[10]。此外,PtpA、SapM等磷脂酶缺失时,海洋分枝杆菌可招募更多的Ptdlns3P,故磷脂酶参与了分枝杆菌的胞内逃逸机制,而且PtpA还可通过阻断V-ATP酶的招募来减少含菌囊泡的酸化,阻断吞噬体成熟[11]。Rv3091属于马铃薯糖蛋白样磷脂酶家族,能够促进结核分枝杆菌从巨噬细胞吞噬体中逃逸,促进细菌的胞内存活[12]。O-甘露糖转移酶(protein O-mannosyl transferase,PMT)能够催化结核分枝杆菌O-甘露糖基化蛋白。PMT缺失时,耻垢分枝杆菌不能阻断吞噬体和溶酶体的融合,导致巨噬细胞内细菌增殖减少,同时也引起细胞分泌TNF-α和IL-6减少[13]。
除了酶类外,Mtb还能产生WhiB3、Rv3463等蛋白质。Wbl家族蛋白WhiB3是结核分枝杆菌whiB编码的一种对氧化还原反应敏感的转录因子,能够检测细菌内氧化还原电势。其在囊泡内酸性pH环境中调节胞质内放线硫醇氧化还原电势(EMSH),从而阻断吞噬体的成熟。此外,WhiB3除了通过改变多聚乙酰脂合成来阻止吞噬体的成熟外,也能够通过缓解由过氧化氢异丙苯、过氧化氢和酸化亚硝酸盐和过氧亚硝酸盐导致的氧化还原压力[14]。Rv3463是从Mtb培养悬液中分离得到的一种蛋白质,它能促进Mtb与早期吞噬体标志(如Rab5、VPS34),及晚期吞噬体标志Rab7共定位,帮助吞噬体成熟。转染Rv3463的耻垢分枝杆菌感染小鼠后,小鼠载菌量明显减少,这些结果提示Rv3463可通过促进吞噬来减少Mtb的感染[15]。Mtb产生的分泌性蛋白ESAT-6(the 6-kDa early secreted antigenic target),又称EsxA,是ESX-1型VII分泌系统的一种关键效应分子。ESAT-6与PDIM作用相似,能够破坏吞噬体膜,PDIM还能促进ESAT-6的作用,而且ESAT-6还能诱导细胞IFN-Ⅰ的产生[16],或诱导巨噬细胞线粒体中超氧化物歧化酶(superoxide dismutase,SOD-2)来帮助细菌存活[17]。此外,ESAT-6还可通过促进HIF1α基因表达来增强巨噬细胞的吞噬能力[18]。ESX-1还可通过下调miR-147-3p来抑制促进海洋分枝杆菌的存活[19]。
2 Rab蛋白对Mtb胞内吞噬过程的影响
Rab GTP酶是小GTP酶Ras超家族中最大的成员之一,目前在人体中已发现超过70种Rab蛋白。编码Rab蛋白的基因约为600 bp,序列保守性较高,在不同物种间其序列同源性高达75%~95%。Rab蛋白约含有200个氨基酸,具有一些共同的结构特征,如G结构域等。Rab蛋白与GTP结合后活化,行使调节胞内膜转运的功能;而当Rab蛋白与GDP结合,Rab蛋白处于失活状态。这一转换过程受到鸟苷酸交换因子(GMP exchange factors,GEFs)和GTP酶活化蛋白(GTPase-activating protein,GAPs)的调控。Rab蛋白在真核生物内几乎所有膜相关细胞器中均存在,调节胞内各种生命活动,包括胞内吞噬作用。
最早有关Rab蛋白调节Mtb吞噬作用的研究主要是分离Mtb吞噬体,检测Rab蛋白的表达差异。强毒株Mtb吞噬体招募的Rab蛋白与普通病原菌和弱毒株Mtb均不同,这表明Rab蛋白在Mtb吞噬中具有独特的作用,具有深入研究的价值。
2.1 Rab5和Rab7
Rab5和Rab7分别是早期吞噬体和晚期吞噬体的标志,常用于吞噬体成熟过程的检测,在麻风病患者的病变皮肤中表达较高。BCG吞噬体仅能够募集Rab5,不能募集Rab7。在鸟型结核分枝杆菌研究中发现类似结果:无论野生型Rab5、活化型Rab5(Q79L Rab5)还是抑制型Rab5(S34N Rab5)均能被募集至骨髓来源的巨噬细胞(bone marrow-derived macrophages,BMMs)内吞噬体上;而野生型Rab7、抑制型Rab7(S22N Rab7)和活化型Rab7(Q67L Rab7)并不能。后证实Rab7能够被募集到Mtb吞噬体上,但募集后很快被释放。这可能与活BCG感染抑制Rab7效应分子RILP(Rab7-interacting lysosomal protein)募集,进而阻止了活化型Rab7与RILP的相互作用,阻断晚期吞噬体形成有关。同时也可能是由于Mtb感染促进吞噬体上Rab7释放,进而导致Rab7-RILP-动力蛋白/动力蛋白激活蛋白复合物分解,从而阻断吞噬体与溶酶体的融合[20]。Rab7还通过影响脂滴与吞噬体之间的相互作用来调节结核分枝杆菌吞噬[21]。
2.2 Rab10蛋白
Rab10最早是在犬肾上皮细胞系MDCK细胞中被检测到,它参与了基底膜蛋白的胞内转运。Rab10能够较Rab5更早地被募集至吞噬体,敲除Rab10或者抑制型Rab10可延迟含热处理Mtb吞噬体的成熟;而过表达Rab10或活化型Rab10能够部分地促进吞噬体的成熟[22],从而促进Mtb的胞内清除,有些miRNA,如miR378d下调时可导致Rab10表达增加,从而促进巨噬细胞吞噬清除细菌[23]。
2.3 Rab14蛋白
Rab14定位于早期内吞体、高尔基体和转运高尔基体等膜结构,参与了这些膜结构的胞内转运过程。沉默Rab14或抑制型Rab14表达时,能够促进Mtb吞噬体成熟。相反,过表达Rab14或活化型Rab14表达时,Mtb吞噬体成熟受阻,维持早期吞噬体特征[24]。
2.4 Rab20蛋白
Rab20定位于高尔基体、内质网以及其他内吞体,属于干扰素-γ(interferon-γ,IFN-γ)诱导可产生的GTP酶,其在活动性肺结核病患者的痰液标本中表达较高。沉默Rab20或抑制型Rab20表达时能促进吞噬体的成熟,同时缩短吞噬体上Rab5存在的时间。但Rab20并不影响Mtb的存活。此外,Rab20能够募集Rab5鸟苷酸交换因子Rabex5至吞噬体,能短暂地增加吞噬体上活化型Rab5的表达[25],使得Mtb吞噬体更久地维持在早期吞噬体阶段。
3 Mtb通过调节Rab蛋白而影响胞内吞噬过程
有关Mtb菌体成分通过调节Rab蛋白而影响胞内吞噬作用的研究已有报道,如PknG。Rab7L1(小鼠Rab29)与细胞内膜-反面高尔基体网络(trans-Golgi network,TGN)的完整性相关,能够调节从高尔基体到溶酶体的转运过程,能够被募集到含菌吞噬体上。PknG缺失的耻垢分枝杆菌和BCG能够在敲除Rab7L1的巨噬细胞内存活,当PknG存在时, 能够阻止Rab7L1转变成Rab7L1-GTP,导致吞噬溶酶体形成受阻[26]。
4 问题与展望
Mtb胞内吞噬过程十分复杂,涉及到Mtb菌体成分与胞内吞噬过程的相互作用。一方面,Mtb可通过3条途径来调节吞噬过程(图1):第一,抑制吞噬溶酶体酸化,减少吞噬细胞对细菌的清除;第二,阻断吞噬体成熟过程,帮助细菌在胞内长期存活;第三, 破坏吞噬体膜, 增加胞质内细菌的免疫逃逸。
另一方面,不同Rab蛋白在Mtb吞噬中作用不同,如活化型Rab10等能够抑制Mtb吞噬,而抑制型Rab14等具有阻断Mtb吞噬作用。虽然已有Mtb菌体成分与Rab蛋白在胞内吞噬过程中的相互作用方面的研究,但目前仍然比较少,许多问题尚未解决。如哪些Rab蛋白在Mtb胞内吞噬中发挥了关键作用,哪些Mtb菌体成分能够调控Rab蛋白来阻断吞噬途径。这些方面的深入研究将有利于揭示Mtb胞内免疫机制,为结核病的治疗和疫苗的研发提供新的思路和研究靶点。
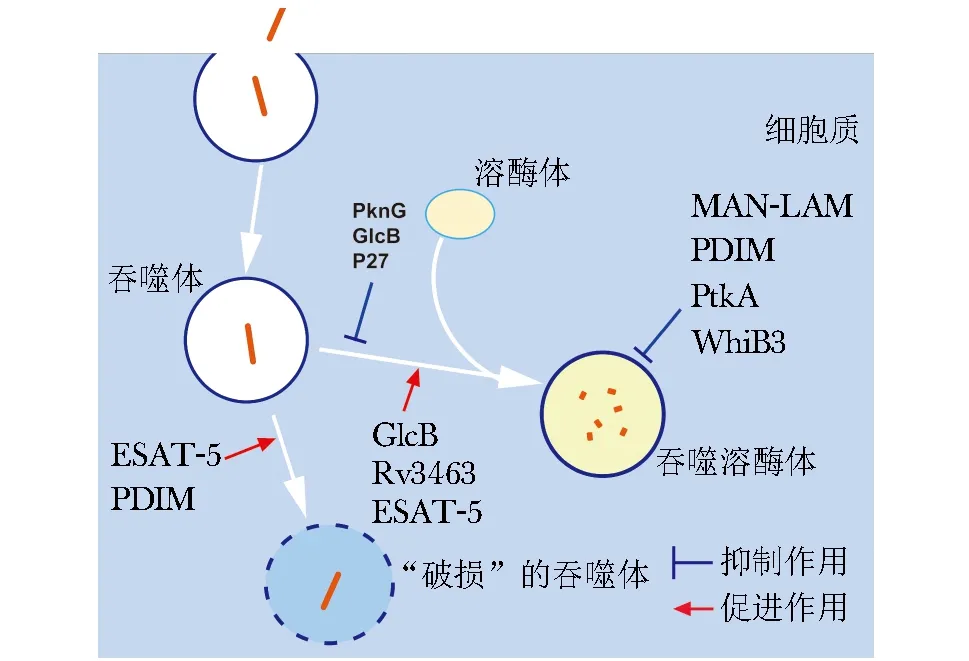
图1 结核分枝杆菌菌体成分对胞内吞噬作用的示意图Fig 1 Schematic diagram indicating the effect of the components of M.tuberculosis on intracellular phagocytosis
