腾格里沙漠东北缘人工植被对土壤物理性质的影响
2022-09-24赵晨光李慧瑛鱼腾飞陈薇宇谢宗才张斌武
赵晨光, 李慧瑛, 鱼腾飞, 陈薇宇, 谢宗才, 张斌武, 张 军
(1.阿拉善盟林业草原研究所,内蒙古 阿拉善 750306;2.中国科学院西北生态环境资源研究院阿拉善荒漠生态水文试验研究站,甘肃 兰州 730000;3.内蒙古巴丹吉林自然保护区雅布赖工作站,内蒙古 阿拉善 737300)
荒漠化是困扰全球的主要环境问题之一[1],而人工植被恢复是干旱、半干旱区荒漠化防治的重要途径之一[2]。相较于传统的植苗造林,飞播造林因具有速度快、范围广、省劳力、投入少、能深入人烟稀少的偏远地区等优势[3],成为我国干旱、半干旱区一项快速而有效的生态恢复措施。阿拉善盟地处亚洲大陆腹地,境内分布着巴丹吉林、腾格里和乌兰布和三大沙漠,境内沙化土地面积约占国土面积的73.67%,沙化土地治理任重而道远[4]。自1984年在腾格里沙漠东缘的头道沙子飞播花棒、沙拐枣和籽蒿获得成功以来[4],截至2019 年底,已成功飞播造林41.2×104hm2,覆盖度在30%以上的保存面积达到了24.5×104hm2[5]。阿拉善地区飞播造林具有历时长、面积大、自然环境条件复杂等特点[6],是干旱区(降水量100~250 mm)飞播造林的典型区域。因此,深入研究飞播造林对土壤理化性质的影响,对指导干旱区人工植被建设具有重要的理论与实践意义。
已有大量研究表明:传统的植苗造林能够显著改变干旱沙区土壤的理化性质[7-9]。首先,人工植被建植后沙地土壤黏粒和粉粒显著增加,土壤质地明显改善[9-13]。例如,巴丹吉林沙漠西南缘梭梭人工建植46 a 后,冠层下和冠层外表层0~10 cm 黏粒分别增加了5.8%和3.9%,粉粒分别增加了4.6%和4.4%[10];腾格里沙漠东南缘草方格+人工植被建植46 a 后,表层0~20 cm 黏粒增加了11.0%,粉粒增加高达22.6%[13]。可见,人工植被修复对土壤理化性质的影响存在明显的区域差异。其次,随着土壤黏粒、粉粒含量的增加,土壤容重降低[12-14],土壤含水量增加[14]。上述研究大多集中在人工植苗造林对土壤理化性质的影响方面,而就飞播造林对土壤理化性质的研究相对较少,如钱洲等[15]对半干旱区毛乌素沙地飞播造林后土壤性质变化进行了研究。因此,以腾格里沙漠东北缘1984—2018年飞播植被群落为研究对象,调查典型年份飞播群落的植被和土壤状况,分析干旱沙区飞播造林对土壤物理性质的影响,以阐明飞播造林群落植被-土壤系统演变规律。
1 材料与方法
1.1 研究区概况
研究区位于贺兰山以西的腾格里沙漠东北缘(图1),地形由东南向西北倾斜,属贺兰山洪积扇边缘,地势开阔,衔接起伏的沙丘,地表风蚀作用强,主要以固定、半固定沙地为主[6];土壤类型以非地带性风沙土为主,土壤质地均为砂土;气候属典型的温带大陆性气候,据研究区周边3 个国家基准气象站(阿拉善左旗、吉兰泰和巴彦淖尔公)1978—2017年气象资料,该区年均降水量在100~200 mm之间,平均为142.0 mm,降雨主要集中在6—9月,约占全年的62%;年均气温为8.7 ℃,7月最高气温可达41.1 ℃,1月最低气温达-34.4 ℃;年均风速为7.1 m·s-1,春季和冬季大风天数居多,最大风速可达26 m·s-1。
研究区原生植被以旱生草本植物为主,如沙米(Agriophyllum squarrosum)、沙 蒿(Artemisia desertorum)、沙鞭(Psammochloa villosa)等。自1984 年飞播造林实施以来,植被主要以飞播引种的次生植被花棒(Hedysarum scoparium)、沙拐枣(Calligonum mongolicum)、籽蒿(Artemisia sieversiana)等灌木为主,并伴有蒙古虫实(Corispermum mongolicum)、蒙古韭(Allium mongolicum)、猪毛菜(Salsola collina)、砂蓝刺头(Echinops gmelini)、雾冰藜(Bassia dasyphylla)、小叶棘豆(Oxytropis microphylla)、拐轴鸦葱(Scorzonera divaricata)、针茅(Stipa capillata)等草本。
1.2 试验设计
于2021 年6—7 月在植物生长旺季选择7 个不同年份的飞播区:2021 年(CK)、2018 年(3 a)、2016年(5 a)、2010 年(11 a)、2004 年(17 a)、2001 年(20 a)和1984 年(37 a)为调查样地,以周边未飞播流动沙地为对照组(CK)(图1)。每个年份的飞播区内按照2~3 km 间距布设30 m×30 m 重复样地5 个,总计布设样地35个。每个样地内沿着西南-东北对角线设立10 m×10 m 调查样方3 个,作为飞播区内植物群落特征、土壤理化性质等调查的基本单元。研究区不同恢复年限的飞播群落植被状况及地表形态特征见表1,其中,灌木层组成由优势灌木多度归一化得到,灌木高度为飞播灌木各树种的平均值,灌木层盖度以测定的冠径按圆形计算面积除以样方面积得到,海拔为调查点周边200 m 半径范围内数字高程模型(DEM)提取的平均值和标准差,景观照片为无人机拍摄。
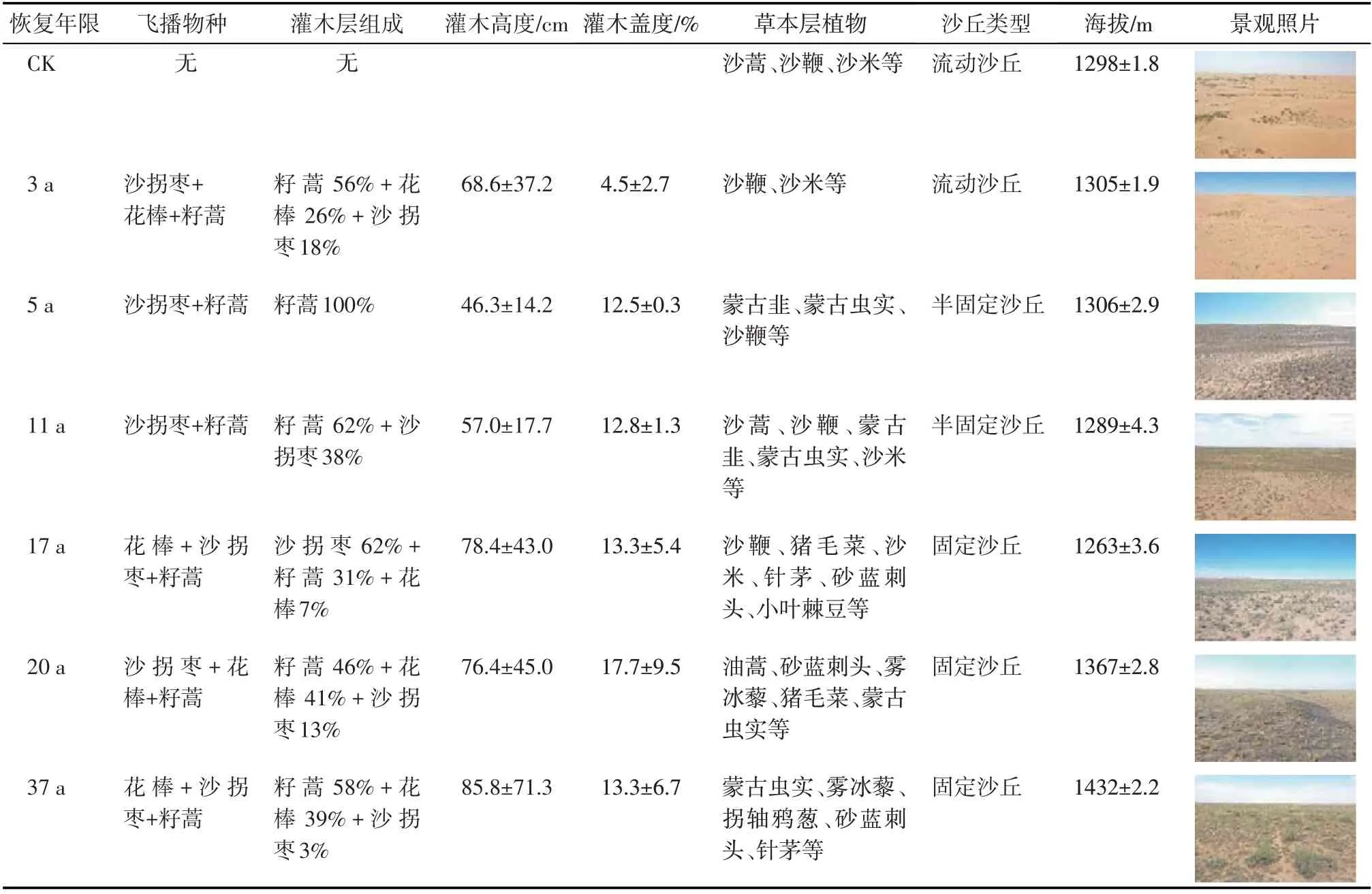
表1 研究区调查样地的植被组成及地表形态特征Tab.1 Vegetation composition and surface morphological characteristics of sample site

图1 研究区位置及调查样地布设Fig.1 Location of the study area and investigation plot setting
每个调查样方内,人工开挖1 m 深度土壤取样坑,按照0~10 cm、10~20 cm、20~40 cm、40~60 cm、60~80 cm 和80~100 cm 分6层取样。每层采用体积为100 cm3的环刀取原状土以测定土壤饱和含水量、土壤干容重和饱和入渗率;采用体积为50 cm3的铝盒取新鲜土样用0.01 g精度电子秤现场测定土壤鲜重。同时,采用土壤取样袋取新鲜土样约500 g带回实验室以测定土壤理化性质。
1.3 土壤物理性质测定
所有土壤鲜土样品带回实验室后,将同一样地内3份样品采用4分法各取1/4组成混合样,置于室内经自然风干后,过2 mm土壤筛,除去树根、叶片等杂物,取10 g左右用于土壤粒径组成的测定。土壤粒径(以体积分数计)采用激光粒度仪测定(Mastersizer2000)。土壤粒径组成采用美国农业部分类系统标准分为7级:黏粒(<2 μm)、粉粒(2~50 μm)、极细砂粒(50~100 μm)、细砂粒(100~250 μm)、中砂粒(250~500 μm)、粗砂粒(500~1000 μm)和极粗砂粒(1000~2000 μm),后5个混合为砂粒(50~2000 μm)[16]。
土壤铝盒样品带回实验室在105 ℃下烘至恒重,测定干重和铝盒重量后计算土壤含水量,即土壤质量含水量。土壤干容重采用环刀法测定,即土壤干容重等于原状土烘干至恒重时的质量除以环刀的体积。将含土环刀用于土壤饱和含水量的测定。首先,将含土环刀带孔一侧用吸水滤纸包裹后浸泡12 h,然后从水中取出静置至环刀内无水溢出,即认为土壤完全饱和,测定鲜重;在105 ℃下烘至恒重,测定土壤干重和环刀重,而后计算得到饱和含水量。
土壤饱和入渗率采用DIK-4012入渗仪测定[17]。首先,将含土柱环刀带孔一侧用吸水滤纸包裹后,将环刀的1/2浸泡在水里,期间不断取出称重,直至环刀土重量达到恒重。将环刀放置在入渗仪上记录样品的入渗量(Q,mL)和入渗时间(t,s),饱和入渗率(K,cm·s-1)使用达西公式计算:
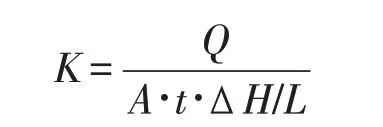
式中:A为土柱横截面积(19.6 cm2);ΔH为水头差(取6.8 cm,水槽液面水位);L为土柱高度(5.1 cm)。
1.4 数据分析
数值采用均值±标准误(Mean±S.E.)表示。首先,采用单因素方差分析(One-Way ANOVA)和Tukey真实差异显著性检验(Tukey’HSD),比较不同恢复年限土壤物理性质组间的差异(n=5)。然后,采用一般线性模型(GLM)分析不同恢复年限土壤物理参数变化趋势,并采用t检验确定变化趋势的显著性。最后,采用Spearman相关系数计算各指标间相关性,采用双尾t检验显著性(P=0.05)。以上所有分析均在SPSS 19.0 统计软件中完成,所有绘图在Sigplot 14.0和R语言软件中完成。
2 结果与分析
2.1 土壤粒径组成
土壤质地改良是荒漠化土地生态修复的第一目标,而土壤颗粒组成变化又是土壤形成、发育及进化的标志[11]。与干旱区其他造林方式相比,飞播造林显著增加了土壤黏粒和粉粒(图2a)。相较于CK,1 m深度内黏粒20 a后从0增加至0.2%,但年际差异不显著(P=0.234);而粉粒20 a 后显著增加至2.5%,37 a 与20 a 无显著差异。这一结果与已有研究一致,但黏粒和粉粒增加的幅度要低于其他飞播沙区或人工造林区[9-12,14-15,18]。例如,腾格里沙漠东南缘(沙坡头)草方格+人工造林30 a后土壤黏粒和粉粒相较于对照组分别增加了1.3%和22.7%[11];乌兰布和沙漠东缘人工造林28 a 后土壤黏粒由0.2%增加到3.0%,粉粒由11.3%增加到40.7%[18];半干旱区毛乌素沙地飞播造林17 a后0~40 cm黏粒增加了5.3%,其中,表层增加尤为明显(11.1%)[15]。由此可见,人工植被修复对于土壤质地的改良存在区域差异,研究区低的黏粒和粉粒可能与该区降水少、沙丘形态起伏大、风沙活动强烈有关。
随着黏粒和粉粒的增加,飞播区土壤中的砂粒明显减少,但各粒径的砂粒变化存在迥异的趋势(图2b)。研究区土壤砂粒主要以细砂粒和中砂粒为主,约占总粒径组成的80%,而极细砂粒、粗砂粒和极粗砂粒不足10%。随着植被恢复年限的增加,极细砂粒呈先增加后减少的趋势,20 a 播区显著高于CK 和其他播区,37 a 和5 a、11 a 和17 a 播区无显著差异;细砂粒随植被恢复年限的增加而减少,37 a播区最低,显著低于其他播区,5 a、11 a和20 a播区无显著差异,但显著高于17 a播区。中砂粒呈先减少后增加的趋势,3 a 播区显著高于其他播区,37 a与对照组无显著差异,11 a和20 a播区最低,显著低于其他播区。粗砂粒呈波动变化趋势,37 a 播区最高,显著高于其他播区,5 a、11 a和20 a播区无显著差异,CK最低。极粗砂粒变化呈先增加后减少的趋势,5 a和11 a最高,17 a、20 a和37 a相对较高,无显著差异,3 a 和CK 无显著差异,接近于0。综上所述,飞播造林减少了沙区土壤粒径占比最高的细砂粒,与CK相比,37 a的播区细砂粒减少了27.1%,极细砂粒和中砂粒分别增加了7.6%和1.3%。这一结果与腾格里沙漠东南缘草方格+人工固沙植被细砂粒的变化不同,该区细砂粒最初是增加的,30 a 后才减少了15%;而中砂粒持续减少,30 a 后减少了约30%[9]。由此可见,不同造林方式对土壤质地的改良效果存在巨大的差异,短期内飞播群落内土壤黏粒和粉粒的增加是有限的,但对土壤中占比最高的细砂粒的消减作用是显著的。
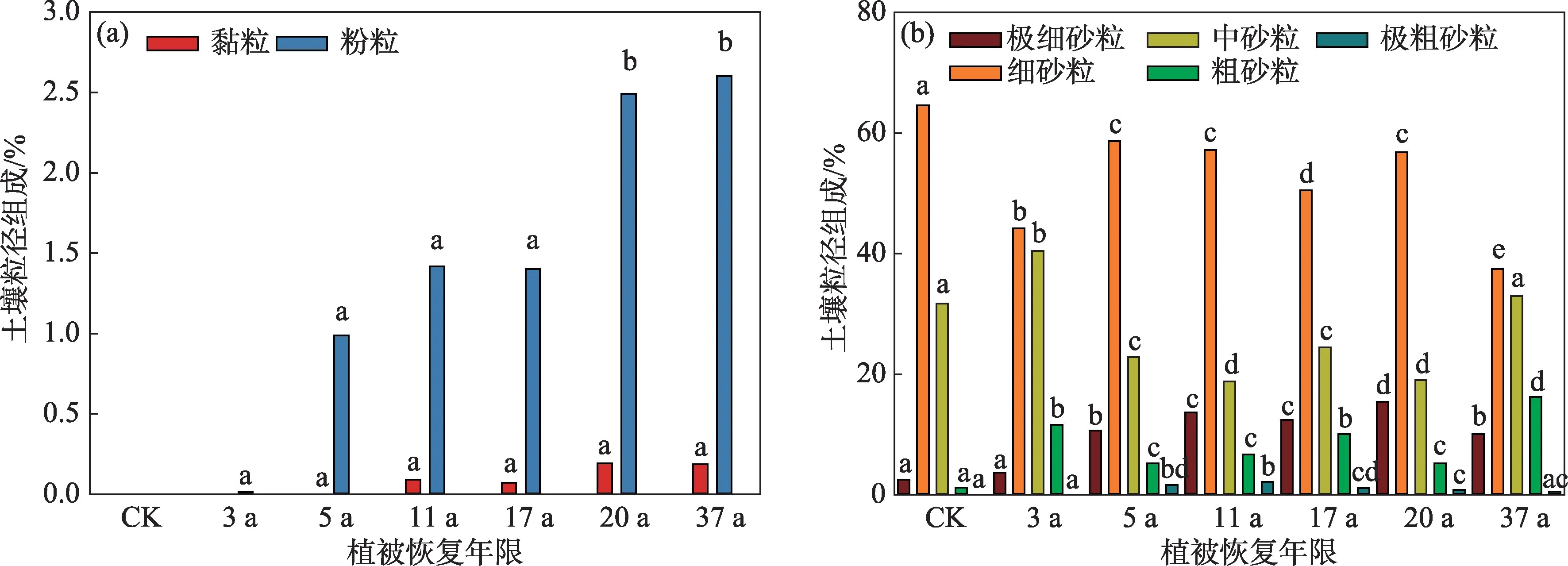
图2 土壤粒径组成随植被恢复年限的变化Fig.2 Change of soil particle size composition with vegetation restoration ages
不同深度的土壤粒径组成以细砂粒和中砂粒为主(图3)。随着植被恢复年限的增加,不同深度的细砂粒均有所减少,粒径较小的黏粒、粉粒和极细砂粒及粒径较粗的粗砂粒和极粗砂粒均有不同程度的增加,中砂粒先减小后增加。相较于对照组(CK),37 a播区的细砂粒0~10 cm土层下降的最多,从65.9%下降至35.7%,下降了32.4%,其次是深层80~100 cm 和40~60 cm,分 别 下 降 了28.2%和27.3%,而10~20 cm 下降最少,仅23.5%。中砂粒含量随着造林年份的增加,不同深度表现出分异的趋势:10~20 cm 和60~80 cm 呈增加趋势,其余各层均下降,其中80~100 cm下降最大,为4.3%。不同深度土壤粒径组成另一个主要的变化是黏粒(除10~20 cm 外)和粉粒均有所增加,其中,深层60~80 cm 和80~100 cm 黏粒和粉粒增加最为明显,黏粒分别增加了0.4%和0.6%,粉粒分别增加了4.7%和4.9%。
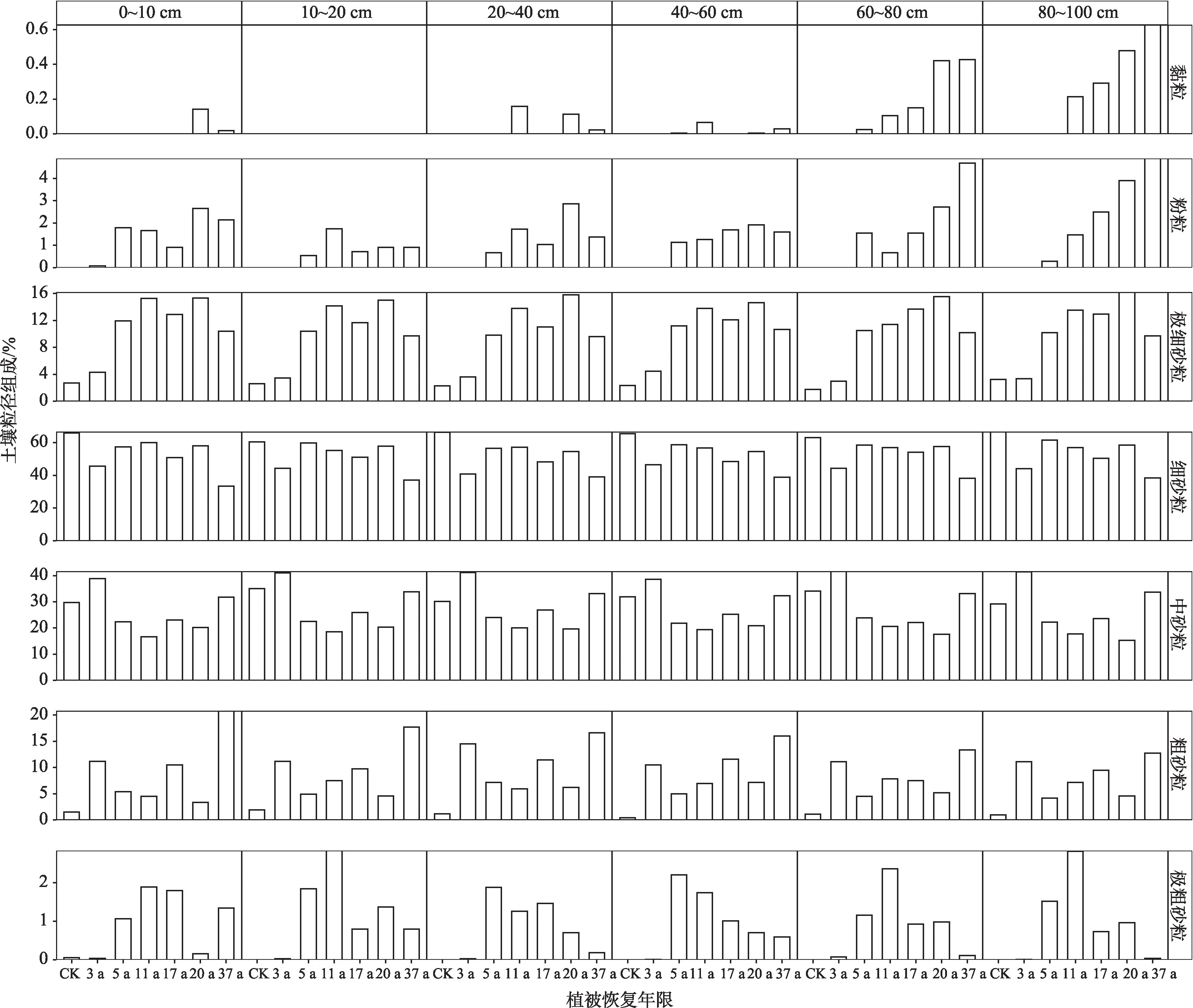
图3 不同深度土壤粒径组成随植被恢复年限的变化Fig.3 Soil particle size composition at different depths changed with vegetation restoration ages
不同深度的粗砂粒均呈明显的增加趋势,其中,表层0~10 cm 增加最多,为19.2%,10~60 cm 大致相当,平均为15.6%,深层60~100 cm增加最少,平均为12.0%;同样,极粗砂粒含量也有类似的趋势,表层0~10 cm 增加了1.3%,10~60 cm 增加了0.5%,深层60~100 cm几乎不变。这一趋势与细砂粒含量变化相反,说明粗砂粒的增加可能是引起细砂粒减少的相对变化。GLM 分析结果表明(表2):随着植被恢复年限的增加,60~80 cm 和80~100 cm 的黏粒和粉粒含量显著增加;极细砂粒增加不显著,细砂粒减少也不显著,但粗砂粒在40~60 cm 处显著增加,其余各层增加不显著;极粗砂粒在表层0~40 cm增加,在40~100 cm减少,均不显著(P>0.05)。

表2 不同深度飞播群落土壤粒径组成随植被恢复年限变化的GLM分析Tab.2 GLM analysis of soil particle size composition at different depths changed with vegetation restoration ages for aerial-seeding biocoenosis
2.2 土壤其他物理性质变化
受土壤粒径组成变化的影响,不同恢复年限的土壤质量含水量、饱和含水量、干容重、饱和入渗率也存在显著差异(图4)。随着植被恢复年限的增加,土壤1 m深度内质量含水量先减少后增加,20 a和37 a的土壤质量含水量相对较高,稳定在1.6%左右(图4a),这与2 个播区黏粒和粉粒含量较高的结果一致(图2a)。与之不同的是,土壤饱和含水量呈显著减少趋势(斜率为-1.49,P<0.05),这与土壤中砂粒含量的变化趋势一致,20 a 播区饱和含水量最高,17 a 播区最低。土壤干容重变化趋势(图4c)与饱和含水量相反,呈逐年增加趋势,20 a 的播区最低,显著低于其他年份。饱和入渗率存在明显的分异特征(图4d),对照组和3 a 的播区显著高于其他年份,这可能与两地土壤近乎由砂粒(约为100%)组成有关。同样,20 a播区饱和入渗率最低,显著低于其他年份,但除了对照组和3 a播区外,其他年份差异不显著。
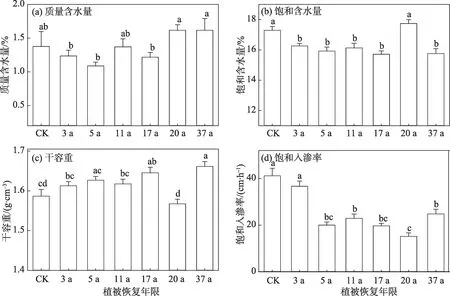
图4 土壤物理性质随植被恢复年限的变化Fig.4 Soil hydraulic characteristics changed with vegetation restoration ages
土壤干容重的变化趋势与已有研究结果不一致[12,14],原因可能是:随着飞播区植被盖度和高度的增加(表2),地表粗糙度增加,表层土壤中较细的颗粒被风吹蚀带走,粗砂粒被保留下来,这可以从37 a播区中表层粗砂粒含量最大得到证实;另外,对照组和3 a播区不同深度极粗砂粒极低,不足0.1%,但在最初的10 a 表层0~20 cm 是明显增加的,而后才开始下降(图2)。从不同恢复年限的景观图和实际调查来看,最初的播区是很少见到草本的,也没有类似人工栽种区草方格的阻挡,地表裸沙明显,较细的颗粒被风吹走(表2)。另外,据不同粒径土壤组分风蚀的风洞实验[19]结果显示:随着风速的增加,中砂粒、细砂粒和极细砂粒的风蚀量迅速增加,而研究区多年平均风速为7.1 m·s-1,为风蚀创造了条件。因此,笔者认为,土壤干容重随着植被恢复年限的增加而升高是因粗砂含量增加引起的,而粗砂含量的增加是风蚀作用的结果。
不同造林年份土壤水力特征参数变化存在明显的垂直分异(表3):表层0~20 cm土壤质量含水量在不同年份存在极显著差异,且呈增加趋势,深层20~100 cm 质量含水量均呈减小趋势,平均减少了0.6%,这与灌木根系的吸水深度大致相当,说明土壤含水量的减少是由植被耗水引起的,这对于土壤含水量极低的沙区影响显著。相较于对照组,20 a后0~10 cm 和10~20 cm 土壤质量含水量分别增加0.8%和0.7%,平均增加了0.7%,这对于浅根系的草本植物生长具有重要意义。不同深度饱和含水量有增有减,但趋势不显著;不同深度土壤干容重几乎无显著变化,而饱和入渗率呈减少趋势。
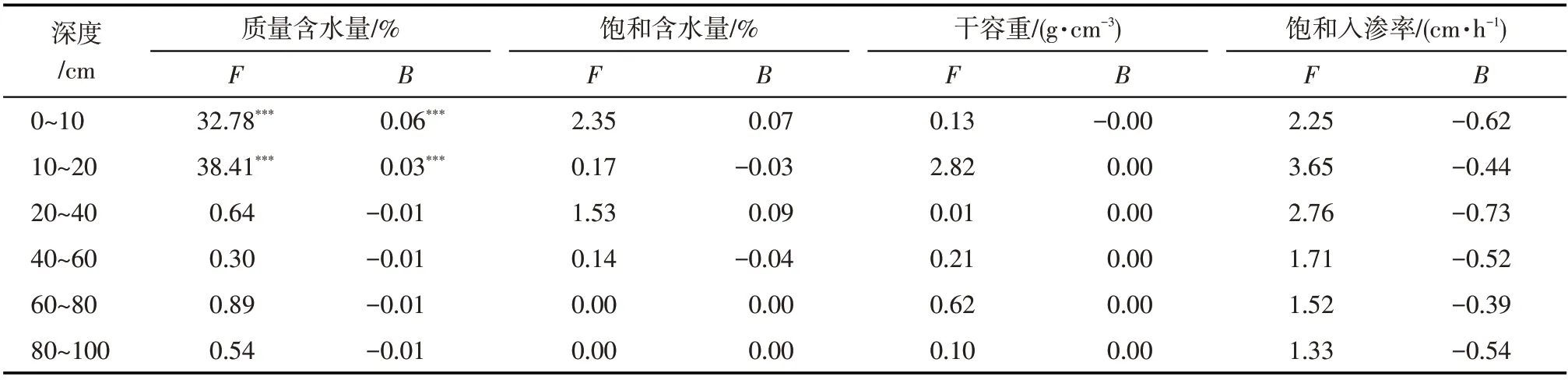
表3 不同深度土壤水力特性随飞播年份变化的One-Way ANOVA和GLM分析Tab.3 One-Way ANOVA and GLM analysis of soil hydraulic characteristics changed with the aerial-seeding ages for different depths
2.3 土壤物理性质的关联
综上所述,飞播造林显著影响着干旱沙区的土壤颗粒组成和物理性质。Spearman 相关分析表明(图5):中砂粒和饱和入渗率、粗砂粒、干容重三者之间显著正相关(P<0.01),而与极细砂粒、极粗沙粒、黏粒、粉粒及细砂粒之间呈显著负相关(P<0.001)。土壤质量含水量和饱和含水量与中砂粒无显著相关,而与干容重呈显著负相关(P<0.01)。土壤干容重与饱和含水量、细砂粒、土壤质量含水量呈显著负相关(P<0.01),而与粗砂粒和中砂粒含量呈显著正相关(P<0.01)。由此可见,土壤干容重的增加是细砂粒含量减少引起的,土壤干容重是反映土壤结构、透气透水性能、土壤养分状况等的重要指标,含矿物质多而结构差的土壤(如砂土)一般在1.4~1.7之间,本研究中平均土壤干容重为1.63 g·cm-3,这与陈闻等[20]对毛乌素沙地花棒群落的测定结果一致。另外,干容重与粗砂粒含量呈显著的正相关进一步证明了干容重增加是由粗砂粒含量增加引起的。随着土壤中细颗粒成分的增加、土壤干容重增加,导致土壤饱和含水量显著下降(图5)。土壤质量含水量除受到土壤自身的物理特性(干容重)的影响外,还与植被生长密切相关。随着植被生长、枯枝落叶增加,输入土壤的有机质增加,土壤表层含水量将增加[21]。

图5 飞播群落土壤物理性质之间的关系Fig.5 Relationship among soil physical characteristics of aerial-seeding community
植被与土壤是一个互馈的过程。随着植被恢复年限的增加,土壤表层大气降尘、枯落物增加,生物过程(细菌、放线菌和真菌的繁殖)加强,促进了表层土壤的成土过程[22]。反过来,表层土壤生物化学过程的演变也影响着植被的演替。一方面,随着表层土壤黏粒、粉粒、有机质含量等的增加,土壤表层形成生物结皮,提高了表层土壤的持水能力和含水量[19];研究区20 a播区的表层土壤饱和含水量高于流动沙地(CK)约2.7%也证明了这一点。随着表层持水能力的增加,隐花植物和浅根系的草本植物构成的草本层片逐步形成[21]。另一方面,随着表层土壤的改良,有效降水很难入渗补给深层土壤,深层土壤含水量随着植被恢复年限的增长而减少,深根系的灌木对水分利用的竞争加剧,使得群落内飞播的深根系灌木物种减少甚至退出。本研究中土壤含水量的变化存在垂直分异,20 a 播区深层(40~100 cm)含水量减少0.7%,浅层含水量(0~40 cm)增加0.6%,这可能导致群落从单一的灌木层片结构演替为灌木层、草本层和藻类、苔藓等隐花植物结皮层组成的复杂结构[23]。
3 结论
荒漠化和土地沙化是一个全球性的环境问题,人工植被恢复是目前防治土地沙化的重要方式之一,深刻理解人工植被恢复过程中土壤理化性质的变化是认识生态恢复与治理成效的重要方面。飞播造林因具有速度快、范围广、省劳力、投入少、能深入偏远地区等优势,成为我国干旱、半干旱区一项快速而有效的生态恢复措施。随着飞播区植被恢复年限的增长,沙地土壤质地得到一定改善,黏粒、粉粒和极细砂粒增加,但这一增加量与其地飞播区(如毛乌素沙地)或其他造林方式(草方格+植苗造林)相比明显偏低,说明飞播造林对沙地土壤质地的改良作用在短期内是有限的。与人工栽植乔木或灌木不同,飞播结合围栏封育的植被恢复更接近于半自然状态,因此对土壤理化性质的影响相对迟缓。另外,土壤含水量的变化存在垂向分异特征,深层含水量减少,浅层含水量增加,这一差异可能导致植物群落由灌木向草本的演替。综上所述,飞播造林对干旱沙区土壤改良具有一定的正效应,但这种正效应在短期内是有限的,土壤质地的变化导致土壤水分状况发生垂直分异,土壤干容重增加,饱和含水量和饱和入渗率下降,提高了土壤的保水能力,增加了土壤水分的有效性。
