基于单位补充量模型的浙江南部海域蓝圆鲹资源评价
2022-09-21崔明远田思泉麻秋云范青松
崔明远,田思泉,2,3,4,麻秋云,2,3,4,范青松
( 1.上海海洋大学 海洋科学学院,上海 201306;2.国家远洋渔业工程技术研究中心,上海 201306;3.大洋渔业资源可持续开发教育部重点实验室,上海 201306;4.农业农村部大洋渔业开发重点实验室,上海 201306;5.浙江省海洋水产养殖研究所,浙江 温州 325000 )
蓝圆鲹(Decapterusmaruadsi)为中上层鱼类,在我国主要分布在东海、黄海和南海,是我国近海重要的渔业资源,主要捕捞方式是拖网和灯光围网。20世纪70年代以来我国蓝圆鲹的产量逐年提升[1],2017年全国渔获量已经达到5.3×105t;在浙江海域蓝圆鲹渔获量仅次于小黄鱼(Larimichthyspolyactis),2017年浙江省渔获量约为6.0×104t[2]。作为我国的重要经济鱼类,蓝圆鲹的渔业资源状况也因此颇受关注,其资源评价的研究多为单位补充量模型,陈国宝等[3-5]均用单位补充量模型研究其资源状态。单位补充量模型将生长、死亡、补充3个生物本身的因素结合起来,是常用的渔业资源评估模型[6]。单位补充量渔获量模型可以根据该群体当前的捕捞水平和开捕年龄来分析其资源利用是否合理[3]。单位补充量亲体生物量模型是防止补充型过度捕捞模型,运用补充亲体量评估渔业资源现状[7]。单位补充量模型对数据要求简单,能够反映当前种群的状态并提供管理参考点,在渔业资源评估中应用广泛[8]。徐旭才等[9]通过Beverton-Holt模型分析了闽南—台湾渔场蓝圆鲹的资源现状,研究发现,补充年龄为0.6龄,捕捞死亡系数为1.4时,持续产量最大;陈国宝等[3]根据1997—1999年南海北部陆架区底拖网调查数据研究蓝圆鲹资源状况。有关蓝圆鲹资源状态的研究较少应用单位补充量亲体生物量模型,但亲体补充量在渔业管理中具有重要作用[10]。冯波等[11]运用单位补充量亲体生物量模型研究了北部湾的带鱼(Trichiuruslepturus)的资源现状,研究发现,在2007—2010年间带鱼资源处于过度利用状态;李跃飞等[12]运用单位补充量亲体生物量模型研究了南海三角鲂(Megalobramaterminalis)的渔业资源状态;李策等[13]运用单位补充量亲体生物量模型研究了南海赤眼鳟(Squaliobarbuscurriculus)的渔业资源状况。以上研究结果均能够为相关渔业资源提供基础知识和管理建议。单位补充量渔获量模型仅从渔业生产角度考量,未涉及种群产卵群体,增加了评估结果的不确定性[11],因此同时考虑单位补充量亲体生物量模型对资源评估的影响将更加合理。
笔者基于2015—2018年在浙江南部近海底拖网调查数据,研究蓝圆鲹生活史特征、单位补充量渔获量和单位补充量亲体生物量,并计算其生物学参考点,评估其资源开发状态,为蓝圆鲹渔业管理提供科学建议。
1 材料与方法
1.1 材料来源
蓝圆鲹样本来源于浙江南部海域(E 120.5°~123.5°,N 27°~29°)的渔业资源专项调查,共设置42个监测站位(图1)。调查船为近海大型底拖网作业船,总吨位约360 t,主机马力403 kW,平均船速为3 kn。在2015年11月—2018年11月期间进行的13次季节调查中,共获得蓝圆鲹样本1134尾(表1)。渔获物冷冻保存,并根据GB/T 12763.6—2007《海洋调查规范》,在实验室内测定样本的叉长、总质量等生物学参数(长度和质量分别精确至1.0 mm和0.1 g)。

表1 浙江南部海域蓝圆鲹不同季节不同年份的样本数量Tab.1 Sample size of Japanese scad in offshore waters of southern Zhejiang in different seasons and years

图1 浙江南部海域渔业资源专项调查的站位设置Fig.1 Survey stations of fisheries resources survey in offshore waters of southern Zhejiang
1.2 生活史特征
浙江南部海域蓝圆鲹体长、体质量关系采用幂指数拟合公式[14]:
m=aLb
(1)
式中,L为体长(叉长代体长)(cm),m为相对应的体质量(g),a为条件因子,b为幂指数系数[15]。通过分析浙江南部海域蓝圆鲹叉长体质量关系,求得参数a为7.4×10-3,参数b为3.17。

图2 浙江南部海域体长时间序列Fig.2 Size time series of Japanese scad in offshore waters of southern Zhejiang柱状图背后颜色的深浅度代表数量的多寡.The dark color in the background in the histogram represents the number.
采用体长频率分析(ELEFAN Ⅰ)法估算生长参数,采用von Bertalanffy生长方程[16]拟合蓝圆鲹的年龄与叉长生长关系:
Lt=L∞[1-e-k(t-t0)]
(2)
式中,Lt为t龄时的叉长(cm),L∞为渐近叉长(cm),t0为理论上体长和体质量等于0时的年龄,k为生长曲线的平均曲率(表示趋近渐近体长的相对速度)(1/a)。
根据Pauly经验公式[17]估算浙江南部海域蓝圆鲹的理论初始年龄(t0):
ln(-t0)=-0.3922-0.2752lnL∞-1.038k
(3)
临界年龄(Tc)是在未捕捞前提下,一个世代的鱼类种群生物量达到最大时的年龄,求解公式[18]如下:
(4)
式中,M为自然死亡系数。
体质量生长加速度为0时对应的年龄为鱼类拐点年龄(Tp),计算公式[6,19]如下:
(5)
运用R语言软件中的TropFishR程序包[20],通过长度转换渔获物曲线法[21-23]计算浙江南部海域蓝圆鲹的总死亡系数(Z)。根据渔获曲线拟合方程中各点相对应的期望值ln (N/dt),计算各点的观测值与期望值之比,进而求得捕捞选择性(S)的相关系数[24-25],其中每个叉长组的数量占总样品数量的比例为N,之后分别除以各叉长组上下限的生长时间(dt)[3]。根据逻辑斯蒂曲线拟合捕捞选择性与年龄的关系[25]:
(6)
式中,St和t为对应的捕捞选择性和年龄,t50为开捕年龄,q为常数。当St=0.5时对应的年龄为开捕年龄(t50)。
根据Pauly经验公式[26]估算自然死亡系数(M):
lnM=-0.0066-0.279lnL∞+0.6543lnk+0.4634lnT
(7)
式中,T为蓝圆鲹生长的水环境温度(℃),L∞为渐近全长(cm),通过全长=1.19475叉长-0.03209(r2=0.996)关系转换。
蓝圆鲹为中上层鱼类,因此取浙江南部近海的中上层水温,根据资源专项调查中的环境因素调查,中上层年平均水温为23 ℃。捕捞死亡系数(F)通过总死亡系数与自然死亡系数的差值(F=Z-M)计算得出。开发率(E)是捕捞死亡系数与总死亡系数的比值(E=F/Z)。
1.3 单位补充量模型
单位补充量渔获量(YPR)模型和单位补充量亲体生物量(SSBR)模型在资源评估中应用广泛[6]。单位补充量渔获量计算公式如下:
(8)
式中,Y为渔获量,R为补充量,S为捕捞选择性,tλ为最大年龄,tR为补充年龄。
经文献查询蓝圆鲹最大年龄为9龄[27]。根据全部样本中第1优势叉长组的中值,通过生长方程估算补充年龄为0.51龄[15],而众多研究均将小型鱼类的补充年龄设为0或1龄[12,13,25],因此本研究中蓝圆鲹的补充年龄为1龄。
将雌性性腺成熟度Ⅲ期及以上视为成熟个体[28],采用Logistic曲线拟合性成熟概率与叉长的关系:
(9)
式中,At为t龄时性成熟概率,L为对应的叉长,r为瞬时性成熟速度,L50%为初次性成熟时的叉长。
单位补充量亲体生物量(SSBR)计算公式如下:
(10)
为评价当前浙江南部海域蓝圆鲹的资源状况,选取了YPR增长率为最大值0.1倍(YPR0.1)时的捕捞死亡系数(F0.1)、YPR最大时(YPRmax)的F(Fmax)、SSBR为未开发状态下20%和40%(SSBR20%和SSBR40%)时的F(F20%和F40%)4个生物学参考点[29-30],用于评价资源状态和提供管理建议。
1.4 敏感性分析
单位补充量模型运行需要众多生长参数,参数的不确定性严重影响模型的准确性和精确性,笔者通过敏感性分析探讨捕捞选择性和自然死亡系数对单位补充量渔获量模型和单位补充量亲体生物量模型估算的影响。通过修改现阶段捕捞选择性的Logistic曲线的斜率得到不同捕捞选择性,将现有斜率(q=-2.451)±0.5和±1.0,其开捕年龄(叉长)也将对应发生变化,最终获得新的选择性,进而得到不同捕捞选择性下单位补充量渔获量模型和单位补充量亲体生物量模型估算结果的变化。查阅文献[31-33]可知,蓝圆鲹自然死亡系数为0.36~0.86,因此探讨6个策略(M=0.35、0.45、0.55、0.65、0.75、0.85)下单位补充量渔获量和单位补充量亲体生物量的变化趋势。
2 结 果
2.1 生活史参数估算
用于蓝圆鲹生长的研究样本实测叉长为4.50~24.70 cm,平均为12.60 cm;实测体质量为0.7~206.6 g,平均为29.05 g。以幂指数形式求得参数a为7.4×10-3,参数b为3.17(图3)。蓝圆鲹的渐近叉长L∞为26.00 cm,生长速率k的估计值为0.19/a,体长体质量理论值为零时的年龄t0估计值为-1.54。计算求得蓝圆鲹体质量生长的拐点年龄估计值为4.53龄,对应的拐点叉长17.79 cm,拐点体质量为67.97 g。临界年龄为2.35龄,对应的临界叉长为13.58 cm,临界体质量为28.88 g。性成熟叉长为20.74 cm。

图3 浙江南部海域蓝圆鲹叉长体质量关系Fig.3 The length-weight relationship of Japanese scad D. maruadsi in the offshore waters of southern Zhejiang
采集的蓝圆鲹样本,叉长为4.50~24.70 cm,以0.50 cm为间隔分为34体长组,通过体长频率求得的渔获曲线(图4)得出蓝圆鲹总死亡系数为1.41(±0.06);求得浙江南部海域蓝圆鲹的捕捞选择性(图5、表2),现阶段其开捕叉长为12.76 cm,对应开捕年龄为2.01龄。估算浙江南部海域蓝圆鲹自然死亡系数为0.55。根据总死亡系数和自然死亡系数,求得捕捞死亡系数为0.86,开发率为61%。
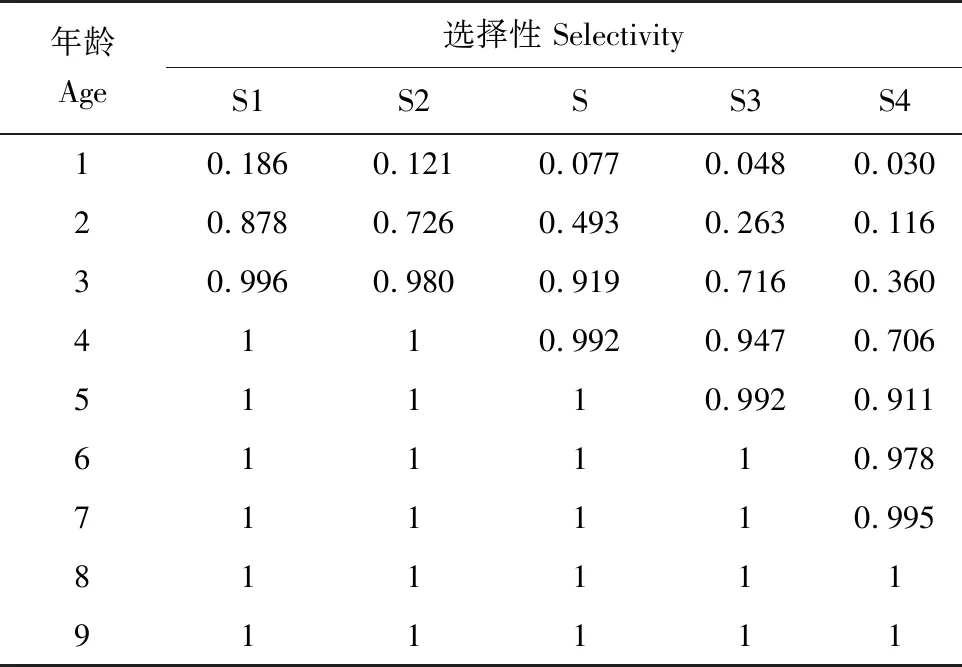
表2 浙江南部海域蓝圆鲹不同捕捞选择性Tab.2 Different catch selectivities of Japanese scad D. maruadsi in the offshore waters of southern Zhejiang

图4 浙江南部海域蓝圆鲹叉长转化的渔获曲线Fig.4 The fork length-converted catch curve of Japanese scad D. maruadsi in the offshore waters of southern Zhejiang每个叉长组的数量占总样品数量的比例为N,之后分别除以各叉长组上下限的生长时间dt,采用ln (N/dt)与其对应的年龄拟合渔获曲线[3].The ratio of the number of each fork-length group to the total number of samples is described as N,which is then divided by the growth time dt of the upper and lower limits of each fork-length group,and the catch curve is fitted with ln (N/dt) and its corresponding age.

图5 浙江南部海域蓝圆鲹捕捞选择性曲线Fig.5 The catch selectivity curve of Japanese scad D. maruadsi in the offshore waters of southern ZhejiangS为当前捕捞选择性,S1~S4为修改捕捞选择性Logistic曲线的斜率得到的不同捕捞选择性;下同.S is the current fishing selectivity,S1—S4 are different fishing selectivities obtained by modifying the slope of the Logistic curve of fishing selectivity;et sequentia.
2.2 单位补充量模型评估
单位补充量渔获量随捕捞死亡系数的变化先快速增加,到达最大值后逐渐减少,并趋于稳定。在当前捕捞死亡系数下,单位补充量渔获量随自然死亡系数值的增大而减小(图6),当前单位补充量渔获量由15.417减至5.130。在不同开捕叉长下,当前单位补充量渔获量由9.828减至8.129,单位补充量渔获量随捕捞死亡系数增大而变化复杂:当捕捞死亡系数在0~1时,单位补充量渔获量随开捕叉长的增大而减小;当捕捞死亡系数在1~9时,S3对应的单位补充量渔获量随捕捞死亡系数的增加逐步增大为最大值,其余捕捞选择性对应的单位补充量渔获量随开捕叉长的减少而减少;当捕捞死亡系数大于9时,单位补充量渔获量随着开捕叉长的增大而增大(图7)。

图6 基于不同自然死亡系数的浙江南部海域蓝圆鲹单位补充量渔获量与捕捞死亡系数的拟合曲线Fig.6 The fitting curve between yield per recruitment and fishing mortality coefficient of Japanese scad D. maruadsi based on different natural mortality in the offshore waters of southern ZhejiangFmax和F0.1为生物学参考点,下同.Fmax and F0.1 are biological reference points,et sequentia.

图7 基于不同选择性的浙江南部海域蓝圆鲹单位补充量渔获量与捕捞死亡系数的拟合曲线Fig.7 The fitting curve between yield per recruitment and fishing mortality coefficient of Japanese scad D. maruadsi based on different selectivities in the inshore waters of southern Zhejiang
在当前开捕年龄(2.01龄)和自然死亡系数(0.55)下,对应的F0.1估计值为0.596,此时的YPR0.1为8.812,对应的Fmax估计值为3.602,YPRmax为10.593。当开捕年龄不变,自然死亡系数由0.35增至0.85时,对应的F0.1和Fmax分别由0.369、0.846增至1.205、15.000,对应的YPR0.1和YPRmax分别由14.015、15.418减至5.767、8.218(表3)。当自然死亡系数不变,开捕年龄由1.43增至3.40时,对应的F0.1和Fmax分别由0.515、2.079增至0.842、7.533,YPR0.1由8.874先增至8.887后减至8.080,YPRmax由10.321增至10.600后减至10.490(表4)。
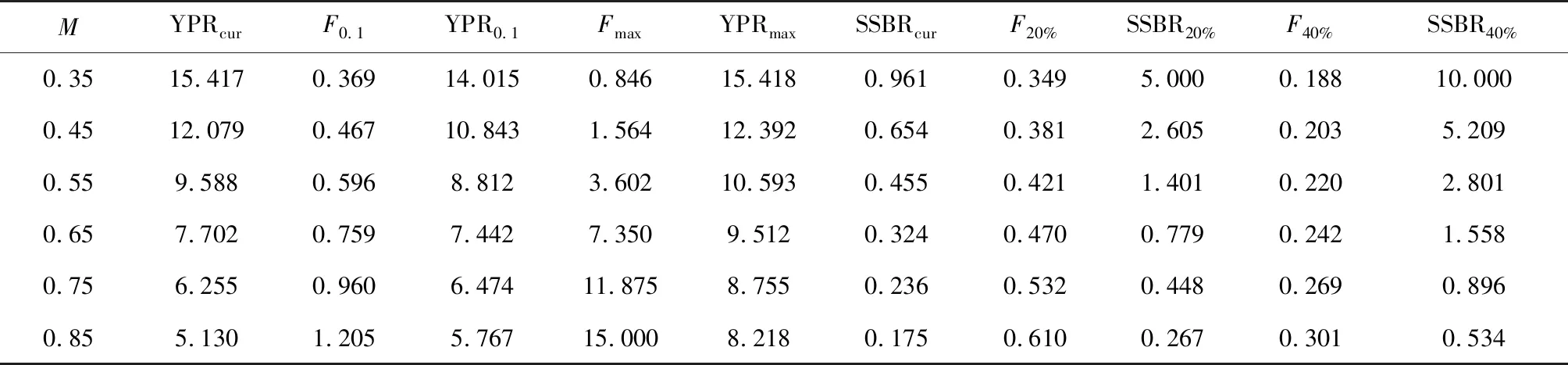
表3 浙江南部海域蓝圆鲹基于不同自然死亡系数的生物学参考点Tab.3 Biological reference points of Japanese scad D. maruadsi based on different mortalities in the offshore waters of southern Zhejiang

表4 浙江南部海域蓝圆鲹基于不同选择性的生物学参考点Tab.4 Biological reference points of Japanese scad D. maruadsi based on different selectivity coefficient in the offshore waters of southern Zhejiang
在当前捕捞强度和开捕叉长下,相对于未开发状态,参考点F20%和F40%分别为0.421和0.220。当前的单位补充量亲体生物量为0.455,相当于初始亲体量(当F=0时对应的单位补充量亲体生物量)的6.5%,远低于防止补充性过度捕捞的警戒线20%。在当前开捕叉长下,单位补充量亲体生物量随自然死亡系数的增大而减小(图8)。当自然死亡系数由0.35增至0.85时,对应的当前单位补充量亲体生物量由0.961降至0.175,对应的F20%和F40%分别由0.349、0.188增至0.610、0.301,SSBR20%和SSBR40%分别由5.000、10.000降至0.267、0.534。在当前捕捞死亡系数下,单位补充量亲体生物量随开捕叉长的增加而增加,当开捕年龄由1.43龄增至3.40龄时,对应的当前单位补充量亲体生物量由0.291增至1.102,对应的F20%和F40%分别由0.358、0.192增至0.695、0.334,对应的SSBR20%和SSBR40%分别为1.401、2.801(表4、图9)。
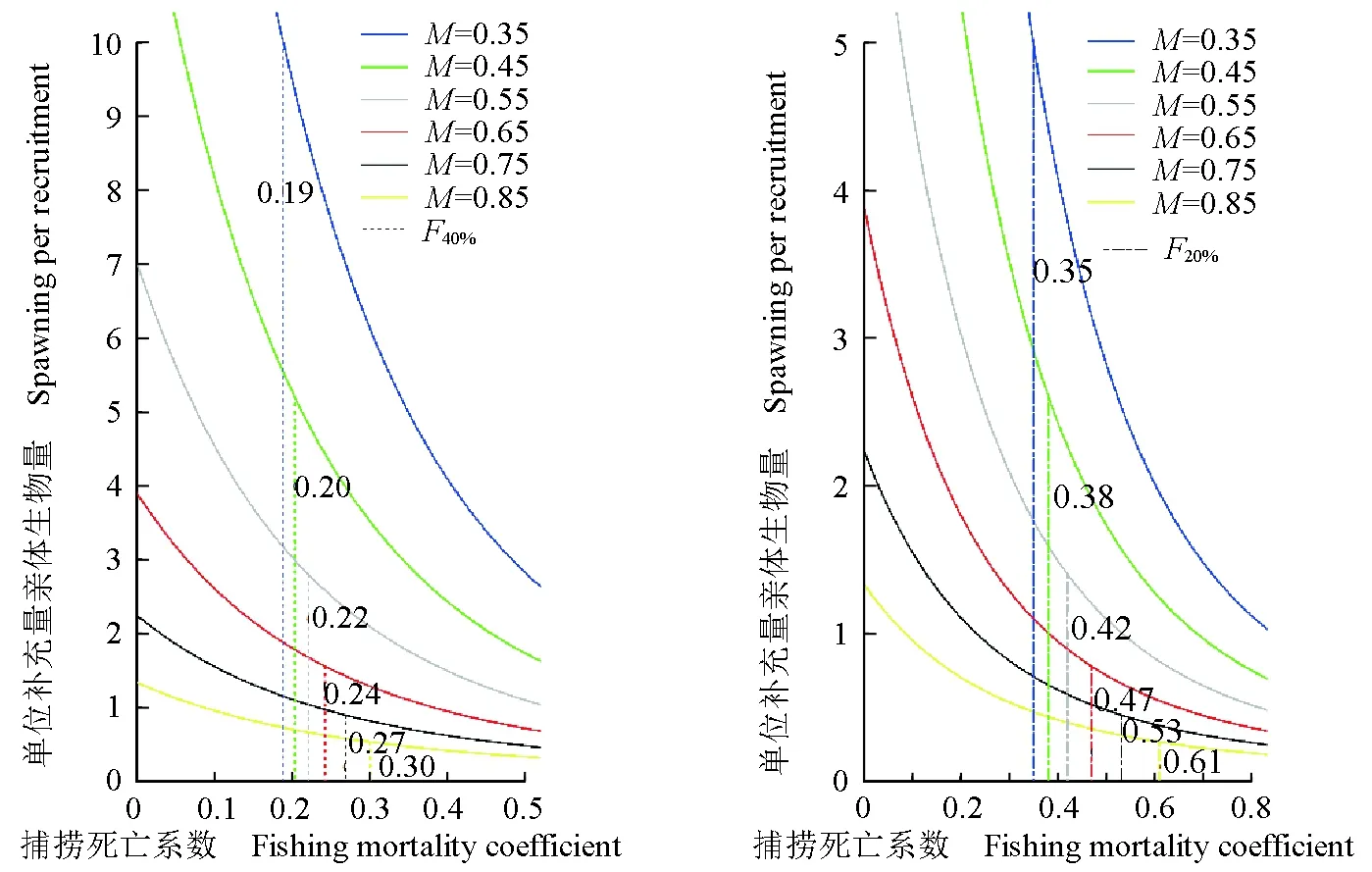
图8 基于不同自然死亡系数的浙江南部海域蓝圆鲹单位补充量亲体生物量与捕捞死亡系数的拟合曲线Fig.8 The fitting curve between spawning stock biomass per recruitment and fishing mortality coefficient of Japanese scad D. maruadsi based on different natural mortalities in the offshore waters of southern ZhejiangF40%和F20%为生物学参考点;下同.F40% and F20% are biological reference points;et sequentia.

图9 基于不同选择性的浙江南部海域蓝圆鲹单位补充量亲体生物量与捕捞死亡系数的拟合曲线Fig.9 The fitting curve between spawning stock biomass per recruitment and fishing mortality coefficient of Japanese scad D. maruadsi based on different selectivities in the offshore waters of southern Zhejiang
3 讨 论
在本研究中,初步了解浙江南部海域蓝圆鲹的生活史特征,蓝圆鲹叉长体质量关系中生长因子a为0.0074,异速生长系数b为3.17,渐近叉长L∞估计值为26.00 cm,生长速率k的估计值为0.19/a,体长体质量理论值为零时的年龄t0估计值为-1.54。通过Pauly经验公式求得蓝圆鲹自然死亡系数为0.55,体长转换渔获曲线求得总死亡系数为1.41(±0.06),当前捕捞死亡系数为0.86,开发率为61%。单位补充量渔获量模型和单位补充量亲体生物量模型分析表明,浙江南部海域蓝圆鲹的渔业资源已处于补充型过度捕捞状态,且不同捕捞选择性和自然死亡系数对单位补充量亲体生物量模型和单位补充量渔获量模型有显著影响。
3.1 生长参数求解方法的适用性
体长频率分析法拟合生长方程时需要按照鱼类体长频率的时间序列对样本的体长频率重组,找出建立在波峰波谷上的Score指数的最大值[34]。笔者将叉长数据以0.50 cm为组距分组并统计不同年份不同季节的每个分组的叉长频率,进而以Score指数为判别标准[35],运用软件FiSAT Ⅱ中的体长频率分析法估算生长参数(L∞和k)。Score指数最大值对应的点为最佳点,即所求L∞和k,但所求生长参数必须能够在生物学上有实际意义。体长频率分析法在数据的时间跨度上没有严格的假设[36],有学者同样基于多年数据运用体长频率法求解生长参数,如王雪辉等[37]根据1997—1999年多年的数据,运用体长频率分析法研究南海北部金线鱼(Nemipterusvirgatus)的生长参数。同时,笔者按照上述标准计算每年的生长参数,发现差异并不显著,因此,笔者运用4年的数据求解生长参数。此方法虽然应用普遍,但是其适用性还有待进一步探讨。叶婷等[38]认为,影响样本体长频率结构的原因较多,如渔具选择性等,这会对研究结果产生一定影响。本研究中,因数据有限而未能采用耳石、鳞片、脊椎骨等更加合适的材料进行年龄鉴定,如梁正其等[39]选取耳石、脊椎骨、鳃盖骨、胸鳍棘作为耳石年龄的对比材料,进而选取年龄鉴定的最佳材料,得到比较可靠的年龄数据。在今后的研究中,应加大力度采集鱼类的相关硬组织,鉴定年龄后进行生活史参数估算和资源评估,其结果将更加准确可靠。
3.2 浙江南部海域蓝圆鲹资源状况
通过在单位补充量渔获量模型中估算出的生物学参考点F0.1和Fmax与当前捕捞死亡系数比较,可以评估种群目前的利用状态[40]。F0.1、Fmax通常作为预防性参考点,在预防生长型过度捕捞中有较大意义[10]。本研究结果显示,捕捞死亡系数为0.86,蓝圆鲹当前捕捞强度没有达到单位补充量渔获量模型的最大捕捞强度,表明其渔业资源未达到生长型过度捕捞程度[30]。根据单位补充量亲体生物量与捕捞死亡系数的分析结果,蓝圆鲹生物学参考点F20%、F40%分别为0.421、0.220,当前捕捞死亡系数(0.86)远大于参考点,单位补充量亲体生物量仅为未开发状态的6.5%,远低于防止补充型过度捕捞警戒线(20%),已处于补充型过度捕捞状态。在当前开捕叉长下,如果追求利益最大化,加大开发力度,可能会造成单位补充量亲体生物量下降,从而造成资源衰退,不利于渔业可持续发展。
根据单位补充量渔获量模型和单位补充量亲体生物量模型的分析结果,仅用单位补充量渔获量模型分析鱼类资源状态可能存在不确定性,同时考虑单位补充量亲体生物量模型能够更全面地了解当前鱼类资源开发状态。在预防性参考点上,较多研究者认为F0.1是更稳健的参考点,但也有研究者认为,F0.1不是建立在维持产卵亲体生物量上,是有可能导致补充型过度捕捞的,因此F0.1和F40%常被一起用来评价鱼类资源的状态[41,12]。然而较多蓝圆鲹资源状态的研究仅用单位补充量渔获量模型,如:宣立强[5]运用Berverton-Holt模型对南海北部底拖网蓝圆鲹资源进行评估,结果显示,20世纪60年代中期开发不足,蓝圆鲹等渔获量曲线中最大单位补充量渔获量超过22.20 g,开捕年龄0~2龄,捕捞死亡系数0.50~1.00比较合适;陈国宝等[3]根据1997—1999年南海北部陆架区底拖网调查数据对蓝圆鲹研究发现,在法定捕捞叉长12 cm下,最大单位补充量渔获量约为26,同时增加捕捞死亡系数与开捕长度时,单位补充量渔获量也增加,但是单位补充量渔获量增加速度会减慢。以上研究仅考虑生长型过度捕捞状态,未考虑补充型过度捕捞是否发生。在大多数渔业种群中,补充型过度捕捞更容易导致种群崩溃,且难以修复[30]。在当前捕捞选择性和捕捞死亡系数下,单位补充量亲体生物量为0.455,相当于初始亲体量的6.5%,远低于防止补充型过度捕捞的警戒线(20%),表明浙江南部海域蓝圆鲹渔业资源处于补充型过度捕捞状态。当捕捞死亡系数减至0.421时,单位补充量亲体生物量为1.401,约增加了当前单位补充量亲体生物量的两倍,因此,建议适当降低捕捞强度,促进渔业资源恢复。
捕捞选择性对单位补充量渔获量和单位补充量亲体生物量有重要影响,当前捕捞强度和选择性下,开捕叉长为12.76 cm(2.01龄),随着开捕年龄增加,单位补充量渔获量相应减小,单位补充量亲体生物量相应增加。蓝圆鲹拐点叉长17.79 cm,临界叉长为13.58 cm;林龙山等[42]对1997—2000年东海区底拖网渔业资源调查数据研究发现,东海区蓝圆鲹初次性成熟叉长为16.00~21.00 cm,现阶段开捕叉长(12.76 cm)小于临界叉长和拐点叉长,也远小于初次性成熟叉长,表明当前捕捞压力过大;朱江峰等[43]认为,从资源增殖的方面考虑,应使所有性成熟个体至少有一次产卵机会。因此,建议适当减小捕捞死亡系数,同时提高开捕叉长至21 cm以上,从而保障蓝圆鲹渔业资源补充。
3.3 不同自然死亡系数和捕捞选择性的敏感性分析
捕捞选择性和自然死亡系数是估算种群动力学的重要参数,捕捞选择性的改变会使开捕体长发生改变,能够直接影响到渔获量的增加或降低,从而使资源群体结构发生改变[6]。而自然死亡系数计算方法较多,但以间接方法(经验公式)为主,而且计算结果差异较大,不确定性较高[32]。探讨自然死亡系数和捕捞选择性的误差和波动对评估结果的影响,能够有效提高对模型结果的理解和应用。在渔业管理中,设置捕捞选择性是主要的管理措施之一[6],同时为了评估渔业管理措施的效果,也需要了解捕捞选择性变化后种群动力学和资源评估结果的变化。
笔者通过敏感性分析发现,捕捞选择性和自然死亡系数对模型的影响较大。自然死亡系数由0.35增至0.85时,对应的当前单位补充量渔获量和当前单位补充量亲体生物量分别减小10.29和0.79,变化幅度较大。自然死亡系数在决定种群潜在生产力上发挥关键作用,显著影响资源评估的结果[25]。当自然死亡系数为0.35时,对应的捕捞死亡系数为1.06,对应的开发率约为75%;当自然死亡系数为0.85时,对应的捕捞死亡系数为0.56,对应的开发率约为40%。这会造成单位补充量模型的不确定性,可能对当前渔业资源状态评价造成严重影响。刘逸文等[25]对2016—2017年山东近海底拖网数据分析发现,自然死亡系数的不确定性对口虾蛄(Oratosquillaoratoria)资源当前开发状况的估计影响较大。开捕年龄由1.43龄增至3.40龄时,对应的当前单位补充量亲体生物量增加0.81,对应的当前单位补充量渔获量下降1.70,变化明显,而开捕年龄(叉长)是评价当前资源状态和制定相关策略的关键因素[13]。综上所述,该模型的结果对开捕年龄和自然死亡系数比较敏感,参数的不确定性可能严重影响渔业资源状态评价。
单位补充量渔获量模型和单位补充量亲体生物量模型的求解需要生长参数、死亡系数等众多种群特征参数,若这些参数估算值的精确性不足,可能会造成模型分析结果的不确定性,这些种群特征参数的确定性可能决定了模型的精确性。崔明远等[32]研究发现,蓝圆鲹自然死亡系数差异较大(0.36~1.41),本研究中的自然死亡系数估计值为0.55,在上述范围之中。笔者仅根据Pauly经验公式计算自然死亡系数,其生长参数众多,容易造成计算结果差异。因此,参数的确定性应作为今后研究的重点。笔者仅从保护产卵亲体生物量方向,建议蓝圆鲹的开捕叉长至少为21 cm,尚未从生产角度考虑。在今后的研究中应寻找合适的开捕叉长,既保证渔业生产,又能够保障蓝圆鲹资源恢复。另外,笔者仅对自然死亡系数和捕捞选择性两个参数进行敏感性分析,其他参数尚未考虑。如本研究求得的生长系数为0.19/a,陈国宝等[3]求得1997—1999年南海蓝圆鲹的生长系数为0.36/a,张杰等[31]计算出的1982—1983年东海蓝圆鲹生长系数为0.26/a,生长系数差异明显,在今后的研究中可以考虑生长系数差异对模型的敏感性影响。
4 结 论
笔者研究了浙江南部海域蓝圆鲹生长特征参数,并采用单位补充量模型分析其资源状态,浙江南部海域蓝圆鲹已处于补充型过度捕捞状态。如果继续加大开发力度,浙江南部海域蓝圆鲹群体可能面临崩溃的情况,建议降低捕捞强度,提高开捕叉长,以保护产卵亲体量。单位补充量模型的求解结果对生活史参数和渔业参数比较敏感,提高相关参数的精确性对模型准确估计至关重要。
