高温胁迫导致水稻光温敏核不育系开颖与雌蕊受精障碍的研究进展
2022-09-13周宇娇张伟杨杨建昌
周宇娇 张伟杨 杨建昌
(1江苏省作物遗传生理重点实验室/江苏省作物栽培生理重点实验室/扬州大学农学院,225009,江苏扬州;2江苏省粮食作物现代产业技术协同创新中心/扬州大学,225009,江苏扬州)
“两系法”和“三系法”杂交水稻是利用杂种优势的2条重要途径[1-3]。与以核质互作雄性不育为技术核心的“三系法”杂交稻相比,“两系法”杂交稻(简称两系杂交稻)是利用光温敏核不育系与恢复系杂交制种,具有一系两用、无需保持系、育种流程简化和杂种优势强等优点,是继“三系法”杂交稻后杂种优势利用的重大科技创新[1-3]。近年来,两系杂交水稻在我国的年种植面积已超过500万hm2,成为水稻杂种优势利用的主要途径,为保障我国粮食安全发挥了至关重要的作用[3-4]。但光温敏核不育系的育性转换受光温条件影响:在杂交制种期间如遇低温(≤23℃),不育系可自交结实导致杂交种子纯度不高;若遭受到高温胁迫(≥35℃)会导致开颖障碍和雌性器官(雌蕊)不能正常受精,使制种产量严重下降,造成巨大的经济损失[4-9]。
随着全球工业化的不断发展,温室效应增强,全球气温随之增高,预计到21世纪末可能上升4℃~5℃。水稻生长季夜间温度每升高1℃,水稻产量可能减少10%[7]。高温胁迫已成为包括“两系法”杂交稻制种在内的水稻生产的一个主要危害[3,7-9]。以我国一个重要的两系法杂交制种水稻基地江苏省盐城市为例,2010至2020年间,有5年(2010、2013、2016、2018和2020年)在杂交制种期间遇到不同程度的高温危害,导致制种产量大幅下降,尤其是2013年的高温导致两系杂交稻较正常年景减产80%~90%,产量和经济损失巨大[10]。因此,了解水稻光温敏核不育系响应高温胁迫的机理,掌握减轻其高温危害的调控途径与技术,对于提高两系杂交稻制种产量和指导水稻抗高温育种、栽培具有重要意义。为此,本文综述了高温胁迫对水稻光温敏核不育系开颖、雌蕊受精以及影响抽穗的生理机制研究进展,讨论了减轻高温危害的调控措施,并对今后研究提出了建议。
1 高温胁迫导致水稻光温敏核不育系开颖障碍的生理机制
水稻开花包括颖壳张开(主要是外颖张开)、花丝伸长、花药开裂、花粉散落在柱头上、花粉萌发和花粉管向子房伸长等过程。正常开颖(外颖张开)是水稻完成受精的先决条件[11]。水稻外颖基部的1对浆片吸水膨大,变厚脱开内外颖的沟槽,再变宽把外颖推开,即颖花开颖;当浆片失水时,外颖在小穗轴弹性力的作用下会向内颖靠拢,完成闭颖过程。浆片渗透势降低是其吸水膨大的重要条件;浆片中淀粉通过淀粉酶水解成为可溶性糖,以及外部水分和养分的输入是浆片渗透势降低(或浆片细胞压力势增大)的主要原因[11-13]。此外,钾离子(K+)也是植物极为重要的渗透调节物质,在百合、玉米和黑麦等植物浆片渗透势变化中起关键作用[14]。但水稻开花前与开花时浆片中的K+浓度并没有明显变化[13]。由此推测,对于浆片中不含造粉体的植物(如百合、玉米和黑麦等),K+等无机离子浓度变化在浆片渗透势变化中起主导作用;而对于浆片中具有造粉体的水稻等作物,浆片中渗透势变化则主要取决于可溶性糖含量的变化[14]。王忠等[15]进一步观察到,适度增温有利于加强呼吸,增加颖花中二氧化碳浓度,使浆片细胞壁松弛,从而诱导浆片吸水膨大,促进开颖。Chen等[6]观察到,开花期高温胁迫会显著降低水稻光温敏核不育系浆片中的淀粉和可溶性糖浓度,导致浆片渗透势显著提高、含水量显著降低,进而导致开颖率与颖花受精率显著降低。Yang等[9]也观察到非常类似的结果。这些结果表明,浆片可溶性糖含量的降低是高温胁迫导致水稻光温敏核不育系开颖障碍的一个重要原因。
植物激素作为一类植物自身合成的、具有调节作用的痕量小分子物质,对植物生长发育具有重要的调控作用,目前尚未有五大类经典激素,如细胞分裂素(CTK)、生长素(IAA)、赤霉素(GAs)、脱落酸(ABA)和乙烯直接诱导水稻开颖的证据[16]。近年来,有学者[17-21]观察到,新型植物激素茉莉酸类(JAs),如茉莉酸(JA)和茉莉酸甲酯(MeJA),不仅在应对逆境胁迫和调控植物生长发育方面发挥重要作用,还可以诱导水稻和小麦等多种禾本科植物颖花开放。有学者[16]观察到,雄性可育的常规水稻在自然开放过程中,颖花中JA生物合成途径中一些关键基因的表达与内源JA水平变化均表现为开颖时急剧上升,闭颖后随即下降。细胞质雄性不育系水稻在缺乏JA时会通过延缓浆片内渗透调节物质和水分的积累来抑制浆片膨大,导致颖花开颖困难,开花时间分散[18]。开花期高温胁迫会导致水稻光温敏核不育系浆片中JA和MeJA含量显著下降,抑制了浆片中可溶性糖的积累,浆片内渗透势增加,吸水困难,致使颖花开颖受阻[9]。喷施外源JAs,一方面可以增强叶片和浆片的抗氧化能力,及时清除过量积累的活性氧,有效地减轻高温胁迫对细胞质雄性不育和光温敏核不育水稻的伤害[6-7,9-10];另一方面,水稻植株内JAs与浆片基部细胞中的JAs受体结合,通过一系列泛素化反应释放以MYC2(JA信号转导的核心转录因子)为主的转录因子,诱导糖转运体基因的表达,将糖和K+转运进入浆片细胞[16,22],同时JAs也能促进浆片细胞内淀粉水解,从而增加浆片中可溶性糖和K+等渗透调节物质含量,使浆片吸水膨胀,促进开颖[6,10,18]。此外,MYC2也是ABA信号传导的激活剂[23],喷施JAs增加了植物细胞中ABA含量,从而刺激气孔关闭,保存植株水分[24],提高耐热性。据此,有学者[6-7,9]认为,提高浆片中的JAs水平是缓解高温胁迫下开花期光温敏核不育系开颖障碍的一条重要调控途径(图1)。
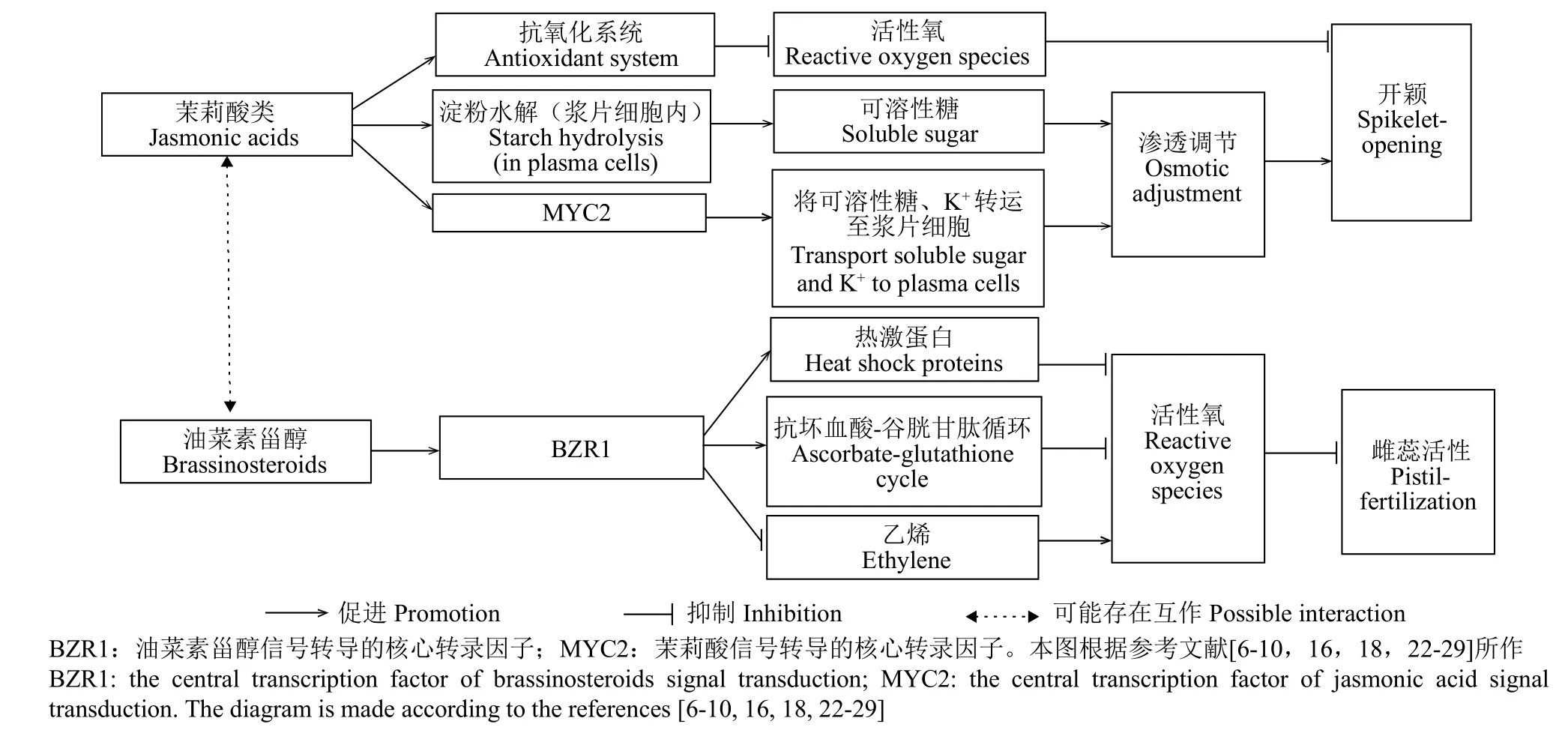
图1 茉莉酸类和油菜素甾醇缓解高温胁迫下水稻光温敏核不育系开颖和受精障碍的调控网络图Fig.1 Schematic diagram of jasmonates and brassinosteroids in alleviating heat stress to photo-thermo-sensitive genic male sterile(PTSGMS)rice line
植物激素间通常存在相互作用与平衡,共同调节植物的生长发育与逆境胁迫响应[7,30]。Yang等[7]观察到,喷施低浓度的促生长型激素如玉米素核苷(ZR,CTK的一种高活性形式)、IAA、GAs、24-表油菜素内酯(24-EBL)或ABA(10μmol/L),对水稻光温敏核不育系开颖无显著影响,但喷施高浓度的ABA或1-氨基环丙烷1-羧酸(ACC,乙烯的生物合成前体)使水稻光温敏核不育系开颖率显著降低。这进一步说明JAs可能对缓解开花期高温胁迫下水稻光温敏核不育系的开颖障碍有独特的作用;同时也表明JAs、ABA和乙烯对光温敏核不育系开颖的调节存在拮抗作用,它们之间的互作机制值得进一步深入探究。
2 高温胁迫对雌蕊形态与活性影响的生理机制
高温胁迫下,水稻雌蕊(由柱头、花柱及子房构成)比雄蕊更具有耐热性,但长时间的高温胁迫也会对雌蕊造成严重的损伤[31]。目前高温胁迫对雄蕊发育影响的研究[32-34]较多,对雌蕊的研究[35]较少。当温度高于23℃或24℃时,水稻光温敏核不育系表现为完全雄性不育[6-7]。雄性器官发育正常的恢复系(父本),其花粉粒被光温敏核不育系柱头接收后从柱头细胞中获取水分[10],萌发成花粉管并进一步完成受精过程[36],该过程与水稻光温敏核不育系雌蕊的形态与生理活性均密切相关。因此,水稻光温敏核不育系雌蕊的形态恶化与生理活性降低是开花期高温胁迫对其伤害的另外一个重要原因。
开花期高温胁迫会造成柱头萎蔫、灼伤,导致柱头面积降低[37-38],不育系获得恢复系(父本)花粉粒的能力明显降低,最终导致受精率降低,杂交制种产量下降[39-40]。高温胁迫不仅影响水稻柱头面积,还会影响柱头外露率。柱头外露的颖花能接受更多的花粉粒,因此其受精能力远高于柱头不外露的颖花[41-42]。柱头外露率与柱头活性有关,高温胁迫使不育系柱头活性显著下降,柱头外露率随之下降[6],导致不育系受精率降低。高温使水稻柱头生理状态发生改变,导致其表面分泌物减少、乳突细胞膜脂过氧化,致使花粉粒不易附着在柱头上,影响花粉的萌发,直接影响水稻的受精与结实[10,22,43]。高温胁迫会显著降低雌蕊细胞中过氧化氢酶(CAT)活性和抗坏血酸(AsA)含量等抗氧化物质水平,破坏其抗氧化系统,导致雌蕊细胞中活性氧(ROS)过量积累,造成细胞膜系统损伤,雌蕊细胞程序性死亡和生理活性显著降低[44-46]。此外,高温胁迫会破坏雌蕊细胞中Ca2+梯度,使得花粉管无法向指定方向生长[47-49];还会显著降低雌蕊细胞中可溶性糖和腺嘌呤核苷三磷酸(ATP)等的含量,导致雌蕊引导花粉管向胚囊延伸过程中的能量供应不足,降低受精率[50-52]。
雌蕊活性与其中的激素种类和含量水平密切相关。Zhang等[45]发现高温胁迫显著降低了日本晴及其高温敏感型突变体雌蕊中IAA的含量与合成水平,进而致使雌蕊活性降低,最终导致水稻受精结实率下降。油菜素甾醇(BRs)是继IAA、GAs、CTK、ABA和乙烯之后发现的第六大类植物激素,普遍存在于植物界,对植物的生长发育过程与抗逆胁迫具有重要的调节作用[53-55]。在高温胁迫下,BRs信号由细胞质膜上的BRs受体感知,通过一系列磷酸化与去磷酸化反应解除对核心转录因子BRASSINAZOLE-RESISTANT1(BZR1)的抑制作用,使其进入细胞核调控BRs响应基因的转录,将BRs信号向下传导,进而提高植株的耐热性[7,25]。高温胁迫会导致植物体内不耐热的蛋白质失活,并诱导植物细胞程序性死亡,从而导致热害的发生[52,56]。喷施BRs可以增加水稻[26]和西红柿[27]等植物体的热激蛋白含量,从而防止蛋白质变性并促进受损蛋白质重新折叠,使蛋白质免受不可逆的热害损伤,增强植物在高温胁迫下的抗逆性。Yang等[7]和Chen等[8]观察到,雌蕊中具有较高BRs含量的水稻光温敏核不育系在开花期高温胁迫下所受的损伤较小,雌蕊受精能力更强;喷施外源BRs可显著提高水稻光温敏核不育系雌蕊中参与抗坏血酸-谷胱甘肽(AsA-GSH)循环酶的活性以及编码这些酶的基因的表达[28-29],显著降低其雌蕊中ACC和H2O2含量[8],增强其雌蕊抗氧化能力,进而有效减轻开花期高温胁迫对水稻光温敏核不育系雌蕊活性的负面影响(图1)。
BRs与JAs、ABA及乙烯之间在对高温胁迫下水稻光温敏核不育系雌蕊活性的调控上也可能存在交互作用。对开花期高温胁迫下的水稻光温敏核不育系同时喷施MeJA和BRs比单一喷施两者的某一种更能增强雌蕊活性(AsA含量更高,H2O2含量更低),受精率与杂交制种产量也更高[7-8]。在甘蓝型油菜叶片上施用BRs可显著增加其内源ABA浓度并增强油菜植株对高温胁迫的耐受力[57]。在拟南芥和玉米植株上施用高浓度的ABA则显著降低其内源BRs水平,致使其对高温胁迫的耐受力变差[58-59]。此外,BRs和乙烯在对高温胁迫下水稻光温敏核不育系雌蕊活性的调控上存在明显的拮抗作用,即喷施外源BRs后可显著降低水稻光温敏核不育系雌蕊中ACC含量,导致乙烯生物合成受阻,进而显著降低其雌蕊中的ROS水平,有效减轻高温胁迫对水稻光温敏核不育系雌蕊受精的不利影响[7-8](图1)。这些结果表明,提高水稻光温敏核不育系雌蕊中内源BRs的生物合成能力或降低乙烯的水平是增强高温胁迫下雌蕊活性与受精能力的一个重要途径。
3 高温胁迫对抽穗包颈的影响
对于常规水稻而言,在减数分裂期遭遇高温胁迫主要通过降低花药开裂率和花粉育性进而降低其结实率和产量[32],但水稻光温敏核不育系在温度高于24℃时,雄性器官(花粉)即表现不育。因此,对于水稻光温敏核不育系而言,抽穗前遭遇高温胁迫主要的危害是加剧了不育系的抽穗障碍(如包颈现象)。水稻细胞质雄性不育系和光温敏核不育系在抽穗时均普遍存在不同程度的包颈现象(稻穗不能完全抽出,穗基部颖花包裹在剑叶叶鞘中,无法接受外来花粉而受精结实)[60-62]。水稻不育系抽穗包颈特性直接影响其异交受精率,是制约杂交种子生产的最大因素,也是评价不育系实用价值和经济价值的重要指标之一[63-64]。水稻不育系包穗是由于穗下倒1节间薄壁细胞(尤其是中部区段细胞)无法正常伸长与扩张,进而使穗下倒1节间缩短或退化缺失所致[65-66]。减数分裂期前后高温胁迫会进一步加剧水稻不育系抽穗包颈现象,进而严重阻碍其接受父本的花粉,显著降低不育系制种产量。但目前关于水稻不育系抽穗包颈性状(尤其在高温胁迫下)的机制研究[10]较少。
多种植物激素均可一定程度上影响水稻节间伸长,其中以GAs对水稻雄性不育系抽穗包颈的缓解作用最为明显[67-69]。此外,BRs在促进茎鞘纵向伸长方面也发挥着至关重要的作用[10,56]。喷施外源BRs可不同程度地解除或缓解由于减数分裂期前后高温胁迫导致的雄性不育系水稻抽穗包颈现象[10]。但不同雄性不育水稻材料本身的抽穗包颈程度及其对激素的敏感性有较大差异,如中国育成的首个实用光温敏核不育系培矮64S不仅包颈严重,且对GAs钝感,制种田一般需要喷施450~750g/hm2GAs才能解除其包颈性状,种子生产成本增加,且容易造成环境污染[70-74]。近年来,隐性长节间基因eui(elongated upper most-internode)被发掘和鉴定[75-78]。eui种质能促进穗下倒1节间伸长,可在不影响不育系水稻花粉育性、半矮秆表型及杂种优势的前提下有效解除雄性不育系水稻的抽穗包颈性状,并能提高不育系的部分农艺性状(如剑叶会变长变宽,柱头外露率也有所提高等)[79]。因此,培育能够解除抽穗包颈性状的雄性不育系水稻是降低杂交稻制种成本和减轻高温胁迫的重要途径之一。
4 减轻水稻光温敏核不育系高温危害的调控措施
4.1 氮肥
氮素是影响水稻产量和品质的重要因素,适当增施氮肥可以降低植株冠层及穗部温度,提高水稻抗热害能力,使水稻结实率和千粒重增加[80-81]。高温胁迫下增施氮肥可提高水稻根系氧化活性和叶片光合作用,增强叶片氮代谢相关酶和籽粒蔗糖-淀粉转化相关酶的活性,减少植株体内活性氧的产生,增强植株活性[82-83]。在开花期增施氮肥通过降低水稻光温敏核不育系已开颖颖花温度和雌蕊活性氧水平,提高促生长类激素含量,增强碳氮代谢关键酶等生理活性,有效缓解开花期高温胁迫对水稻光温敏核不育系颖花开放和雌蕊活性的负面影响,从而提高不育系与恢复系杂交后的受精率、结实率和制种量[10]。因此,当开花期稻株含氮量低于1.8%时,适当增施氮肥是缓解开花期高温胁迫导致水稻光温敏核不育系颖花开放和雌蕊受精障碍的一个重要栽培措施[10]。
4.2 灌溉方式
传统观点[84-85]认为,水稻抽穗开花期遇高温,需要对稻田进行深层灌水以降低冠层温度,减轻高温对水稻的伤害。但有学者[86-87]观察到,高温对水稻的危害与水稻冠层相对湿度密切相关;在同一温度下,相对湿度越高,高温伤害越严重。轻干湿交替灌溉可以显著降低稻田的相对湿度,减轻高温对水稻的伤害,从而显著提高水稻的结实率、千粒重和产量。此外,轻干湿交替灌溉还可以增加水稻叶片中抗氧化物质和CTK含量,改善根系形态,提高根系代谢功能,协调地上部生长,使水稻在高温胁迫下保持正常的生理功能[86-88]。Zhang等[89]观察到,稻穗发育期采用适度落干可显著提高水稻幼穗中BRs的生物合成,促进颖花分化并减少颖花退化,增加有效颖花数。这些结果表明,采用轻干湿交替灌溉可有效减轻高温胁迫对光温敏核不育系水稻的伤害,但其机理有待深入研究。
4.3 播期
调整播期可以避免或减少水稻生长发育中的极端环境气候(如开花期高温),提高水稻的产量和品质[90-94]。推迟播期可以使中籼稻和晚粳稻在各生育阶段内积温和累计日照时数降低,营养生长期缩短,生育进程加快[94-95]。近年来,华中地区高温危害频发,可推迟水稻播种期,使抽穗后前20d的日均温在24.9℃~26.4℃,可以显著提高产量和资源利用效率[91]。调整播期或是有效减轻水稻光温敏核不育系高温危害的另一个重要栽培措施。但是,推迟播期会缩短生育期,在开花期不发生高温的年景(如2021年)有可能降低水稻产量;在多熟制地区还会影响下茬作物的正常播种。
4.4 喷施激素
激素在植物应答高温胁迫等逆境响应过程中发挥着重要作用[16]。已有研究[6-10,16,18,22-29]证明,JAs对光温敏核不育系水稻开颖具有独特的正向调控作用,BRs分别对光温敏核不育系水稻雌蕊活性和缓解抽穗包颈均具有重要的正向调控作用。因此,在生产实践中可通过适当施用JAs和BRs来有效缓解高温胁迫对水稻光温敏核不育系造成的危害。但JAs和BRs相关制品的购买成本较高,且可能会造成环境污染,如何协调这一矛盾还需进一步探讨。
4.5 分子育种
通过传统育种方法或基因工程技术提高水稻光温敏核不育系浆片中JAs或雌蕊等器官中BRs的生物合成与信号传导,进而培育出耐热性强的光温敏核不育系水稻品种,是缓解或解决高温胁迫对水稻光温敏核不育系造成危害的另一个有效措施。随着分子生物学与基因工程的飞速发展,水稻中已经克隆出JAs的生物合成关键基因[16](如OsDAD1-3、OsLOX-RCI1、OsAOS1、OsAOC 和OsOPR7)、信号转导关键基因[96](如OsJAR1、OsCOI1b以及13个OsJAZs)、BRs生物合成关键基因[93](如D11、D2和BRD2)和信号转导关键基因[97](如OsBZR1),并对这些基因的功能已有较为深入全面的认识。因此,上述基因均可直接加以利用,培育出耐热性强的光温敏核不育系水稻品种。
5 展望
目前,人们对高温胁迫导致水稻光温敏核不育系开颖与受精障碍的机理认识较少,仅有的研究或侧重于某个独立器官的性状变化,或侧重于某个单独激素的响应与作用,而缺乏对其系统的认识。因此,建议今后可从以下5个方面深入研究水稻光温敏核不育系开颖与雌蕊受精的高温危害。
(1)研究高温胁迫下水稻光温敏核不育系包颈特性的分子机制,阐明高温加剧不育系包颈程度的内在机理;重点研究GAs和BRs缓解或解除水稻光温敏核不育系包颈的机理,探明缓解高温下水稻光温敏核不育系包颈加剧的有效调控措施。
(2)研究高温胁迫下水稻光温敏核不育系根系形态生理特征与根系化学信号传递、植株地上部(茎、叶与穗)激素含量或比例的变化以及碳氮代谢特性等与光温敏核不育系水稻开颖与受精的内在联系。分别从植株、器官与细胞水平等不同层次上阐明高温胁迫下水稻光温敏核不育系开颖与雌蕊受精障碍的内在机理。
(3)结合采用激素相关水稻突变体或转基因材料,利用现代分子生物学的技术手段重点研究JAs和BRs的生物合成、信号转导及其与其他激素(特别是ABA与乙烯)调控高温胁迫下水稻光温敏核不育系开颖与雌蕊受精障碍的互作机制。
(4)研究氮、磷、钾等矿质营养元素以及水分灌溉和播期等栽培措施对缓解水稻光温敏核不育系高温危害的作用及其机理,并探明缓解高温对水稻光温敏核不育系危害的环境条件指标,提出缓解水稻光温敏核不育系高温危害的栽培调控途径和关键技术。
(5)使用常规育种方法、生物信息分析、高通量全基因组测序和蛋白质组学等技术挖掘调节水稻光温敏核不育系包穗、开颖和雌蕊活性的有利基因和分子标记,培育出耐热性强的光温敏核不育系水稻品种。
