不同轮作模式下旱地番茄土壤理化性质及细菌群落组成特征
2022-09-07李万星李小霞靳鲲鹏韩文清刘永忠黄学芳苏秀敏曹晋军
李万星, 李小霞, 李 丹, 靳鲲鹏, 韩文清, 刘永忠, 黄学芳, 苏秀敏, 王 佼, 曹晋军
(1.山西农业大学谷子研究所,山西 长治 046000; 2.山西农业大学山西有机旱作农业研究院,山西 太原 030000)
番茄广泛种植于世界各地,既可作为蔬菜也可作为水果,喜食之人甚多,以温室栽培为主。相对于温室番茄,旱地番茄种植于旱季,靠自然降雨生长,多种植于山西省东南部,尤以长治市壶关县生产的旱地番茄闻名。旱地番茄具有含糖量高、酸度低、番茄红素含量高等特点,且沙瓤细腻、果形圆润鲜亮、口感佳,因而深受各地人们喜爱[1]。但为追求经济价值,当地百姓采用多年连作不换茬的种植方式,连作障碍现象严重,导致旱地番茄病害加重,土壤质量下降,严重影响旱地番茄品质。
在同一土地连续种植同种或同科作物,会使土壤酶活性下降和土壤微生物结构恶化,引起土传病害和产量降低,产生连作障碍。轮作不同作物可以改善土壤物理性质[2],均衡土壤养分[3],改变土壤酶活性[4],改善作物根际土壤微生物结构[5],减少土传病害的发生。杨尚东等[6]研究发现番茄连作降低了土壤细菌的丰富度和多样性,土壤菌群失衡,而轮作黄瓜、白菜和菜豆等可以提高土壤微生物数量和土壤酶活性。细菌是土壤中最活跃的生物成分,在土壤中比例最大[7],其在土壤中的丰富度和群落结构会因前茬作物的不同而不同。本研究以旱地番茄为研究对象,通过轮作不同作物,研究土壤容重、团聚体结构、土壤养分及土壤酶活性,并应用高通量测序技术研究土壤细菌多样性及群落结构组成,探索旱地番茄连作障碍的形成机制,以期为通过轮作方式缓解旱地番茄连作障碍提供理论基础和实践依据。
1 材料与方法
1.1 试验地概况
试验地位于山西省长治市壶关县东关壁村(113°22′E,36°19′N)。2018年至2020年旱地番茄生育期内平均降水量为367.1 mm,降水主要集中在6、7、8月份,冬季、春季降雨量少,为典型的雨养农业区。年蒸发量2 063 mm,年平均气温9.4 ℃,一月最低气温-13.6 ℃,七月最高气温35.3 ℃,无霜期151 d,年日照时数2 619 h。
1.2 供试材料
供试番茄为日润1号,糯玉米为晋糯10号,西葫芦为东葫1号,秋葵为绿盐,黄瓜为中农18号,花生为长治县裕盛丰农业专业合作社提供,豆角和葱为当地种植品种。
1.3 试验设计
试验期为2018年3月-2020年11月,共7个处理。试验地前茬为旱地番茄,2018年轮作豆角,2019年轮作6种作物,2020年所有处理种植旱地番茄,试验设计见表1。每处理设置3个重复,随机区组排列,小区面积52 m2(长8.0 m,宽6.5 m),起垄铺膜,每垄1.3 m,每小区5垄。播前统一撒施发酵过的鸡粪,施肥量为54 000 kg/hm2,整个生长过程中不追肥,各小区管理方式一致。

表1 试验的处理设置
1.4 土样采集
2020年旱地番茄拉秧前于每个小区采集土壤,采用五点法取样,土钻钻取0~20.0 cm表层土层(内径为7 cm),将土样混合,自然风干,捡去石块、根等杂物,按照鲍士旦方法[8]过不同孔径的筛子得到较细腻的土样用于测定土壤养分。拉秧前挖取旱地番茄的完整根部,将大块土壤抖落至铁盒,并将附着于根毛的土壤刷下,用筛子(1 mm)筛至自封袋内用于测定土壤酶活性。用铁锹挖取旱地番茄(三穗果时期)完整根部,将土壤抖落至无菌袋中,并将附着在根毛上的土壤轻轻刷下,置于冰盒带回实验室,保存于-80 ℃冰箱用于测定土壤细菌组成。
1.5 项目测定
1.5.1 土壤容重和团聚体结构测定 于2020年拉秧前取0~20.0 cm土层测土壤容重(采用环刀法)。分别取0~10.0 cm和10.1~20.0 cm土层的土壤,采用筛分法测定土壤水稳性团聚体。
1.5.2 土壤养分测定 土壤各养分的测定参考鲍士旦的方法[8],pH采用pH计测定,电导率采用P4多功能测定仪测定。
1.5.3 酶活性测定 土壤过氧化氢酶、碱性磷酸酶、蔗糖酶、脲酶和纤维素酶活性的测定参考关松荫的方法[9],蛋白酶活性用从南京卡文思检测技术有限公司购买的试剂盒测定。
1.5.4 土壤微生物的测定 DNA提取和PCR扩增:将样品充分混匀,去除杂质,称取1.00 g鲜样。土壤细菌总DNA提取按照DNeasy PowerSoil Kit(QIAGEN)试剂盒说明书进行,DNA浓度和纯度利用NanoDrop2000进行检测,对细菌DNA的16S rRNA V3-V4区段扩增,建库,并进行质检与定量。利用QIIME2软件对操作分类单元(Operational Taxonomic Units,OTU)在门和属水平上(97%相似度)进行聚类分析。根据样品OTU数量进行Alpha多样性(Chao1、Shannon、Shimpson指数[10-12])分析。
1.6 数据分析
采用Excel2007软件进行数据处理,使用SPSS17.0软件进行方差分析与多重比较(Duncan’s新复极差法)、Person相关分析检验和主成分分析(Principalcomponent analysis,PCA)。
2 结果与分析
2.1 轮作对土壤容重和团聚体结构的影响
轮作不同作物对土壤容重影响不同。由图1可知,与对照比较,LVZm、LVAh、LVAf、LVCs处理显著降低了土壤容重,降幅为5.43%~7.75%,而LVCp、LVAe处理土壤容重显著增加。根据土壤容重分级标准,容重在1.00~1.25 g/cm3范围内属容重适宜,LVZm处理、LVAh处理、LVAf处理和LVCs处理土壤容重适宜,而对照、LVCp处理和LVAe处理容重均大于1.25 g/cm3,土壤属于偏紧和紧实程度。

各处理见表1。不同字母表示处理间在0.05水平上差异显著。 图1 轮作对旱地番茄土壤容重的影响Fig.1 Effects of rotation on soil bulk density of dryland tomato
由表2可知,各处理≥0.25 mm的土壤团聚体含量在0~10.0 cm土层和10.1~20.0 cm土层有差异。与对照比较,LVAh、LVCs处理显著提高了0~10.0 cm土层≥0.25 mm的土壤团聚体含量,其他处理差异不显著;LVZm、LVAh处理显著提高了10.1~20.0 cm土层≥0.25mm的土壤团聚体含量。与对照比较,LVAe处理0~10.0 cm土层<0.25 mm的土壤团聚体差异不显著,其余各轮作处理显著降低了<0.25 mm的土壤团聚体含量,降幅为9.01%~18.90%;10.1~20.0 cm土层,LVZm、LVAe、LVCs处理显著降低了<0.25 mm的土壤团聚体含量,其余轮作处理与对照差异不显著。

表2 轮作对旱地番茄土壤团聚体结构的影响
2.2 轮作对旱地番茄土壤养分的影响
由表3可知,各处理土壤养分差异显著。与对照比较,LVAf、LVAe处理土壤有机质含量与对照差异不显著,其余各轮作处理有机质含量均显著增加。LVAh处理全氮含量显著增加,其余处理与对照差异不显著。LVCs处理全磷含量显著低于对照,其余各轮作处理均显著高于对照,其中LVAf处理全磷含量最高,比对照高36.63%。LVZm处理全钾含量显著高于对照,LVCs处理全钾含量显著低于对照,其余各处理与对照差异不显著。各轮作处理碱解氮含量显著高于对照,增幅为25.11%~102.64%。除LVAe处理有效磷含量显著低于对照外,其余各轮作处理有效磷含量均显著高于对照。LVAe处理速效钾含量与对照含量差异不显著,其余各轮作处理均显著高于对照。pH值各处理土壤均呈碱性,且对照最大为8.63,各轮作处理间差异不显著。对照土壤电导率为112.50 μs/cm,各轮作处理均显著降低了土壤电导率,降幅为18.13%~29.96%,说明各轮作处理可以降低旱地番茄土壤盐渍化程度。

表3 轮作对旱地番茄土壤养分的影响
2.3 轮作对土壤酶活性的影响
由表4可知,与对照相比,LVAh、LVCs处理分别显著提高了过氧化氢酶活性4.87%和3.68%, 而LVAf处理降低了过氧化氢酶活性,其余轮作处理与对照差异不显著。与对照相比,各轮作处理碱性磷酸酶活性均增加,增幅为1.85%~18.38%。与对照相比,LVCp、LVAf、LVAe与对照脲酶活性差异不显著,其余各轮作处理脲酶活性均高于对照,其中LVAh脲酶活性比对照高34.78%。各轮作处理与对照相比均可提高纤维素酶和蔗糖酶活性,增幅分别为22.14%~102.86%和21.43%~122.86%。与对照比较,LVCs、LVAh、LVCp处理显著增加了蛋白酶活性,增幅为39.13%~47.83%。
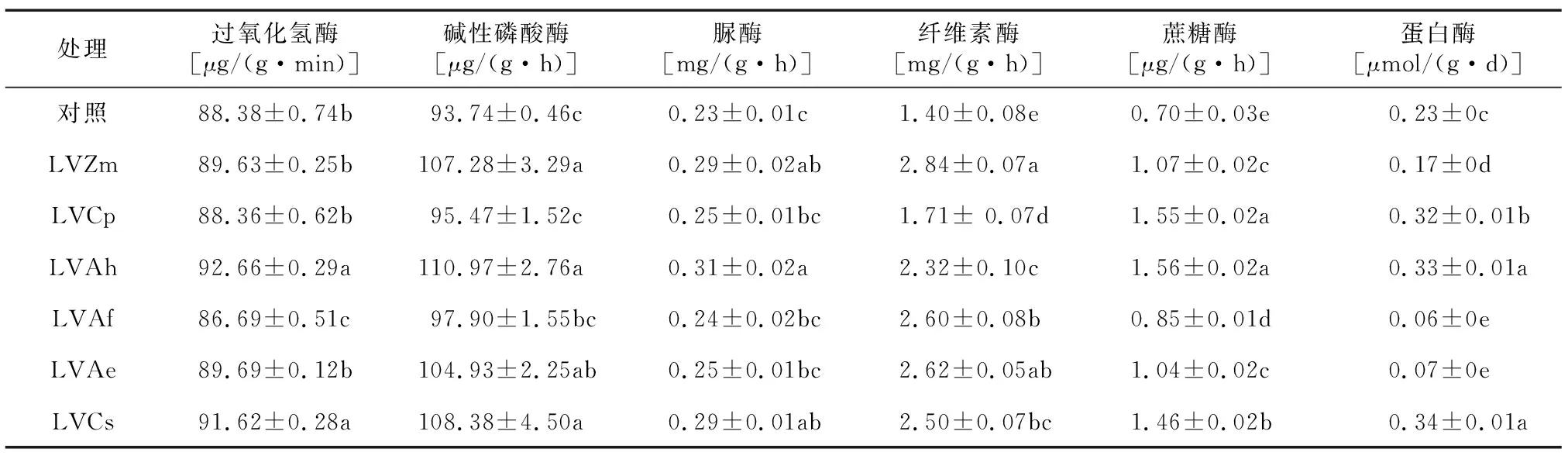
表4 轮作对旱地番茄土壤酶活性的影响
2.4 轮作对土壤细菌Alpha多样性的影响
各处理样品的覆盖度均大于90%,达到分析要求。由表5可知,与对照比较,LVCp处理显著降低了Chao1指数和Shannon指数,其余轮作处理Chao1指数和Shannon指数与对照差异不显著,说明轮作并没有显著增加土壤细菌丰富度和多样性。各轮作处理与对照Simpson指数差异不显著,均大于0.998,说明各处理细菌群落优势度高而多样性低。

表5 轮作对土壤细菌Alpha多样性的影响
2.5 轮作对土壤细菌群落组成的影响
由图2可知,7个处理细菌群落组成主要集中在10个细菌门类,分别是变形菌门(Proteobacteria)(34.97%~39.62%)、放线菌门(Actinobacteria)(23.80%~33.79%)、绿弯菌门(Chloroflexi)(7.35%~10.03%)、酸杆菌门(Acidobacteria)(5.90%~9.60%)、芽单胞菌门(Gemmatimonadetes)(5.80%~8.43%)、拟杆菌门(Bacteroidetes)(4.59%~6.69%)、髌骨细菌门(Patescibacteria)(0.83%~1.82%)、厚壁菌门(Firmicutes)(0.79%~2.02%)、疣微菌门(Verrucomicrobia)(0.80%~1.19%)及硝化螺旋菌门(Nitrospirae)(0.60%~0.86%),这10种菌占所有菌的97.67%~98.51%。由上可知,连作和轮作对土壤细菌门水平群落结构组成无影响,但细菌群落相对丰度有影响。
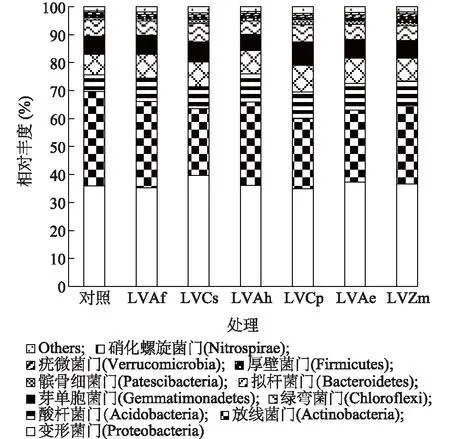
各处理见表1。图2 不同轮作模式土壤在门水平的细菌类群比较Fig.2 Comparison of bacteria groups at phylum level in soils under different rotation patterns
对丰富度排在前6的细菌门进行差异性分析,如表6所示,LVCs处理变形菌门丰富度显著高于对照,其余轮作处理与对照差异不显著;各轮作处理放线菌门丰富度显著降低(LVAf处理除外),绿弯菌门各轮作处理与对照差异不显著;各轮作处理酸杆菌门丰富度显著增加,LVCp处理芽单胞菌门丰富度显著高于对照,拟杆菌门丰富度变化规律不明显。
由于现代分子生物学检测技术的局限性,绝大多数的细菌分类受到限制,因此绝大多数的细菌被归为未知菌属。由图3可知,未知菌属占比约74.93%~77.85%,在已知菌属中丰富度较高的10个属分别为溶杆菌属(Lysobacter)、Subgroup_6、鞘氨醇单胞菌属(Sphingomonas)、藤黄单孢菌属(Luteimonas)、MND1、气微菌属(Aeromicrobium)、KD4-96、Gitt-GS-136、类诺卡氏菌属(Nocardioides)、节杆菌属(Arthrobacter)。不同处理对细菌丰富度有影响,各轮作处理与对照相比,Subgroup_6、藤黄单孢菌属(Luteimonas)、MND1和KD4-96丰富度增加,鞘氨醇单胞菌属(Sphingomonas)、类诺卡氏菌属(Nocardioides)和节杆菌属(Arthrobacter)丰富度降低。
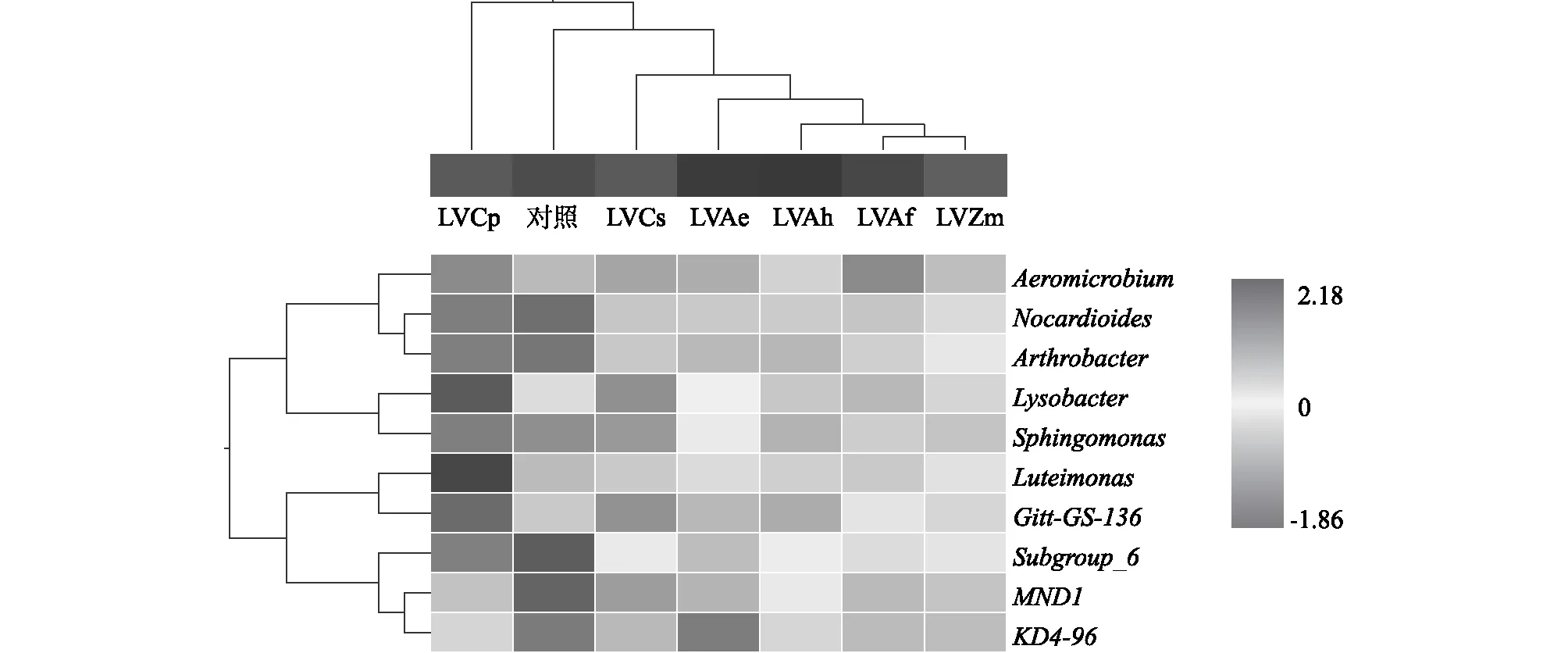
2.6 基于土壤细菌属组成的聚类分析
对相对丰度较高的10个细菌属构建分层聚类,分析7个处理细菌组成的相似性和多样性。由图4可知,7个处理的土壤样品较为明显地聚为6类。LVCp独成一支,说明其细菌属群落结构显著不同于其余处理。LVAf处理和LVZm处理聚为一支,说明二者土壤细菌群落结构相似,其余各轮作处理各为一支,连作(对照)为一支。因此,轮作可对属水平上的细菌群落结构产生显著影响。
2.7 土壤酶活性与土壤细菌优势属间的相关关系
由表7可知,土壤碱性磷酸酶、纤维素酶是影响土壤细菌群落最主要的因子。碱性磷酸酶活性与Gitt-GS-136呈显著负相关关系(P<0.05),纤维素酶活性与KD4-96呈显著正相关关系(P<0.05)。
2.8 不同轮作模式土壤理化性质的主成分分析

各处理见表1。图4 不同轮作模式土壤属水平的细菌类群比较Fig.4 Comparison of bacteria groups at genus level in the soil under different rotation patterns

表7 土壤酶活性与优势细菌属之间的相关性分析

表8 土壤理化因子主成分分析的特征值与方差分析贡献率

表9 土壤理化因子主成分得分与综合得分
3 讨 论
容重是土壤的重要物理性质,是反映土壤结构、评价土壤质量的一项重要指标。土壤容重与土壤紧实度呈正相关关系,适宜的土壤容重能够为作物提供良好的生长环境。张晓霞等[14]发现土壤有机质增加可以降低土壤容重。本研究中,LVZm处理、LVAh处理、LVAf处理和LVCs处理土壤有机质含量高于对照,同时这4个处理土壤容重低于对照。土壤团聚体结构大小可以反映土壤稳定性,相较于非稳定性团聚体,水稳定性团聚体更能体现土壤结构的稳定性[15], ≥0.25 mm水稳性团聚体的数量越多,土壤结构越稳定。各轮作处理(除LVAf处理外)0~10 cm土层 ≥0.25 mm水稳性团聚体含量均高于对照,但差异不显著,<0.25 mm的土壤团聚体含量均显著低于对照(除LVAe处理外),说明轮作在0~10 cm土壤层可以改善土壤结构,增加土壤稳定性,而10~20 cm土壤层变化不明显,可能是因为轮作年限短,各轮作处理在10~20 cm土壤层对团聚体结构的影响还没有显现,有待于进一步研究。
微生物的生命活动是土壤活力和功能形成的关键因子,细菌作为土壤中最活跃的成分,其多样性的变化可以反映土壤微环境的变化[16-19]。本研究中,轮作不同作物对旱地番茄土壤细菌多样性的影响不同。杜思瑶等[17]、张立成等[18]研究发现,轮作比连作土壤细菌群落多样性高,说明轮作可引起土壤细菌多样性的变化。本研究中,LVCp处理显著降低了土壤细菌丰富度和多样性,且其细菌属聚类独成一支,细菌群落多样性不同于连作和其他轮作模式,可能是因为西葫芦在生长周期内相较于其他作物需要施用较多农药,部分农药入土引起了土壤微环境的变化,导致土壤细菌丰富度、多样性降低和细菌群落结构发生变化。其余各轮作处理与对照土壤细菌丰富度和多样性差异不显著,可能是因为在种植前统一撒施了生物有机肥(鸡粪),而生物有机肥作为一种生物调节剂,可以将大量活的有益功能菌带入土壤,起到调控土壤微生物区系、改善土壤生物活性的作用[20],在本研究中轮作处理和对照因为撒施有机肥削弱了各土壤区系微生物的差异,降低了处理间微生物多样性的差异性。
土壤养分是影响土壤微生物群落结构的重要因子[21-23],许多研究结果表明土壤pH是影响微生物的重要因素[24-25]。刘佳等[26]发现,细菌群落结构改变受土壤pH的驱动,pH是决定优势菌相对丰富度及多样性指数的首要因素。本研究中轮作处理的pH值相较于对照pH值降低,酸杆菌门丰富度在轮作处理中高于对照,这是因为酸杆菌门丰富度可以反映土壤营养状况,其丰富度大小在一定程度上反映土壤酸性条件。有研究报道,酸杆菌门细菌能够降解复杂的化合物如纤维素[27],这与本研究中轮作处理纤维素酶含量增加的结果一致。Subgroup_6可以将复杂的有机物发酵成酸[28],这与本研究中轮作处理与对照比较Subgroup_6丰富度增加,同时轮作处理pH降低的结果相一致。本研究中发现轮作处理的土壤中有机质含量比连作(对照)有机质含量高,可能是因为轮作土壤中绿弯菌门细菌含量高于连作(对照),绿弯菌门细菌在良好的土壤状况下选择了光能自养营养方式,消耗土壤中的有机质较少。
轮作不同作物,植物根系与土壤微生物共同作用形成不同的土壤生态环境[29],植物根系、微生物分泌物和动植物残体分解的物质会引起土壤结构的变化。本研究中LVAh处理显著降低了土壤容重,有机质、全氮、碱解氮、速效钾含量显著高于对照,过氧化氢酶、碱性磷酸酶、脲酶、蔗糖酶和蛋白酶活性也显著高于对照和其他轮作处理,与范业庚等[30]的研究结果一致,说明旱地番茄-豆角-花生-旱地番茄轮作模式通过作物根系和残体以及土壤微生物互相作用形成了独特的土壤微环境。轮作豆科作物可显著提高土壤的碳矿化率和酶活性,进而提高土壤有机碳和全氮含量[31],这是因为豆科作物根部的根瘤菌可有效固定空气中的氮。本研究中,在旱地番茄-豆角-花生-旱地番茄轮作模式中,2年连续种植豆科作物,其根部的固氮菌通过强大的固氮作用,为后茬作物生长提供了足够的氮肥,这与该轮作模式中土壤全氮和碱解氮含量最高的结果一致。综上所述,LVAh轮作模式能够较好地改善土壤结构,平衡土壤各养分,调节酶活性,且通过主成分分析也得出了同样的结果。但LVAh轮作模式对土壤细菌影响小,这与吕毅等[32]的研究结果不一致,这可能与不同轮作模式有关,其原因有待进一步研究。
4 结 论
LVZm、LVAh、LVAf和LVCs轮作模式显著降低了土壤容重。轮作降低了0~10 cm土层<0.25 mm的土壤团聚体含量,增加了0~10 cm土层土壤稳定性。各轮作土壤有机质、全氮、全磷、全钾、碱解氮、速效钾、有效磷含量均有变化,轮作显著降低了土壤pH值和土壤盐渍化程度。各轮作处理碱性磷酸酶、脲酶、纤维素酶和蔗糖酶活性高于或显著高于对照。除LVCp轮作模式显著降低土壤细菌丰富度和多样性外,其余轮作模式与对照差异不显著。主成分分析结果表明,LVAh轮作模式优于其他轮作模式,可以作为改善旱地番茄连作障碍的主要模式在当地进行推广应用。
