我国8个水电站15种过鱼对象游泳能力研究*
2022-09-05柯森繁金志军李志敏雷青松王永猛石小涛涂志英
柯森繁,金志军,李志敏,雷青松,王永猛,石小涛,涂志英
(1:三峡大学水利与环境学院,宜昌 443002) (2:湖北省鱼类过坝技术国际科技合作基地,宜昌 443002) (3:中国电建集团贵阳勘测设计研究院有限公司,贵阳 550081) (4:上海勘测设计研究院有限公司,上海 200434) (5:广州市天河区水务设施建设中心,广州 510655) (6:三峡大学三峡库区生态环境教育部工程研究中心,宜昌 443002)
我国是水电资源相对丰富的国家,截至2013年,中国的水电站数量已达到46758座,居世界首位[1]. 高坝大库的水电站建设造福国计民生的同时,却对河流连通性产生了不利影响,阻隔了鱼类的洄游通道,使整个河流生态系统的结构和生态系统的完整性遭到破坏. 过鱼设施作为筑坝河流上下游的连接通道,旨在协助鱼类洄游以及促进上下游物质、能量与基因交流,最大限度解决水电站修建带来的生境破碎问题. 截至2020年8月底,中国现有和在建鱼道已达93座[2]. 但由于长期以来对水生生物的基础研究薄弱,鱼类生态习性、游泳能力等资料严重不足,我国过鱼设施的过鱼效果难以达到预期效果[3]. 为更好地满足鱼类溯河洄游的需求,有必要开展过鱼设施主要过鱼对象的行为学研究,如鱼类游泳能力、趋流特性、底质偏好等.
不同流域上建有过鱼设施的水利工程过鱼目标不同. 课题组通过对布置有电站的长江、珠江、黄河以及雅鲁藏布江现场调查与环评报告等资料收集,发现四大家鱼、裂腹鱼(Schizothorax)和鳅(Cobitidae)已成为上述典型流域的过鱼目标物种[2]. 过鱼设施根据过鱼目标不同生命阶段的过坝需求,分为上行过鱼设施和下行过鱼设施. 上行过鱼设施(如鱼道、升鱼机等)设计主要考虑成鱼需要上行过坝产卵的需求[4],而下行过鱼设施(旁路通道、溢洪道等)设计则主要考虑幼鱼需要下行至坝下肥育的需求[5]. 过鱼对象的游泳能力是过鱼设施结构和水力学设计的关键,其中感应流速(induction velocity,Uind)是指鱼能感应到的最低流速,也是上行过鱼设施内的最小设计流速,低于此流速,成鱼在上行过鱼设施内会迷失方向[6];爆发游泳速度(burst swimming speed,Uburst)是过鱼设施设计的另一重要参数[7],鱼在穿过障碍物、高流速区及漩涡时,常常采用爆发游泳速度[8-10],也称为鱼的最大游泳速度[11]. 上行过鱼设施内高流速区的流速不应大于过鱼对象的爆发游泳速度[12]. 对于下行洄游的幼鱼,虽能主动适应水流的主体流动,但幼鱼并非被动漂向下游,而是利用游泳能力、趋流能力和适应性行为响应水动力变化,以优化洄游时间[13-14]. 当下行过鱼设施的入口流速过大,顺流而下的幼鱼可能会增加摆尾频率以保持位置,或者将趋流方向由负变正,再利用爆发游泳速度向上游逃逸[15]. 我国自1990s初就有鱼类游泳能力的研究[16]. 迄今为止,游泳能力的研究对象主要是四大家鱼和裂腹鱼[2],鳅的游泳能力研究较少. 游泳能力的研究主要集中于临界游泳速度及爆发游泳速度[2],研究结果仍不能满足过鱼设施工程设计的需要. 此外,不同研究人员采用的游泳能力测试装置、测试环境及测试方法等差异较大,这将导致同一种鱼测得的游泳能力结果不同. 多元回归分析是评价鱼类游泳能力的方法之一. Lynda等利用多元线性回归模型评估了鳙的游泳能力、水温、方法和叉长之间的关系[17]. 而国内采用此方法对本土鱼类游泳能力的评估较少.
本研究选取8个电站下游15种鱼为研究对象,采用统一的游泳能力测定方法,测试了每种鱼的感应流速和爆发游泳速度. 将15种鱼分成3类目标鱼(四大家鱼、裂腹鱼和鳅),采用多元线性回归定量分析了水温(T)和鱼体长(BL)对感应流速和爆发游泳速度的影响,建立了游泳能力预测模型,为我国的过鱼设施设计提供建议与参考.
1 材料与方法
1.1 实验材料
实验鱼捕获自我国8个水电站下游. 8个水电站分别为:錾高水电站、木扎提河三级水电站、藏木水电站、果多水电站、两河口水电站、东庄水利枢纽工程、葛洲坝水电站和松新水电站(图1),选择了15种优势种为研究目标,并将捕获的15种鱼分为3组进行研究,分别为Ⅰ裂腹鱼组:厚唇裂腹鱼(RacomalabrosusWang (RL))、斑重唇鱼(DiptychusmaculatusSteindachner (DS))、异齿裂腹鱼(Schizothoraxoconnori(SO))、拉萨裸裂尻鱼(Schizopygopsisyounghusbandi(SY))、长丝裂腹鱼(Schizothoraxdolichonema(SD))、齐口裂腹鱼(Schizothoraxprenanti(SP))、短须裂腹鱼(Schizothoraxwangchiachii(SW));Ⅱ四大家鱼组:青鱼(Mylopharyngodonpiceus(MP))、草鱼(Ctenopharyngodonidellus(CI))、鲢(Hypophthalmichthysmolitrix(HM))和鳙(Hypophthalmichthysnobilis(HN));Ⅲ鳅组:红尾副鳅(Paracobitisvariegatus(PV))、叶尔羌高原鳅(Triplophysayarkandensis(TY))、长身高原鳅(Triplophysatenuis(TT))和斯氏高原鳅(Triplophysastoliczkae(TS)). 15种鱼均为野外捕获,渔获物的体长范围具有随机性,分别集中分布在不同的区间. 各组实验鱼的体长范围及分布,水温范围及样本量见表1、图2和图3. 由于实验装置尺寸的限制,四大家鱼组的实验对象均为幼鱼. 捕获后的鱼置于装有新鲜河水且持续通氧的水槽中暂养(饥饿)48 h.

图1 8个水电站地理位置分布 (该图基于自然资源部标准地图服务网站下载的审图号为GS(2019)4345号的标准地图制作,底图无修改)Fig.1 Geographical distribution of eight hydropower stations in China
1.2 实验装置
游泳能力测定装置为Loligo水槽(Loligo System),水槽有两种不同规格,游泳区体积分别为5 L(型号SY10700)和30 L(型号SY10800). 游泳区流速采用螺旋桨流速计(LS300-A/LS1206B, China)测定. 游泳能力测试水槽用水来源于实验现场的河水,利用潜水泵(型号1WZB50-0.75D, China)将河道中的水抽至蓄水池,经过滤后再引入游泳能力水槽,水槽来水通过尾水管最后排至河道. 过鱼对象的测试水温与河道水温基本一致. 由于测试对象捕获水域不同,且测试时间跨度较大,实验水温的变化与自然水温的变化一致. 水槽上方安装一个海康威视DC-2CD3T35D.I3摄像头(25帧/s)对实验鱼的游泳行为进行录像,观察实验鱼的游泳行为,设施安装在野外环境.
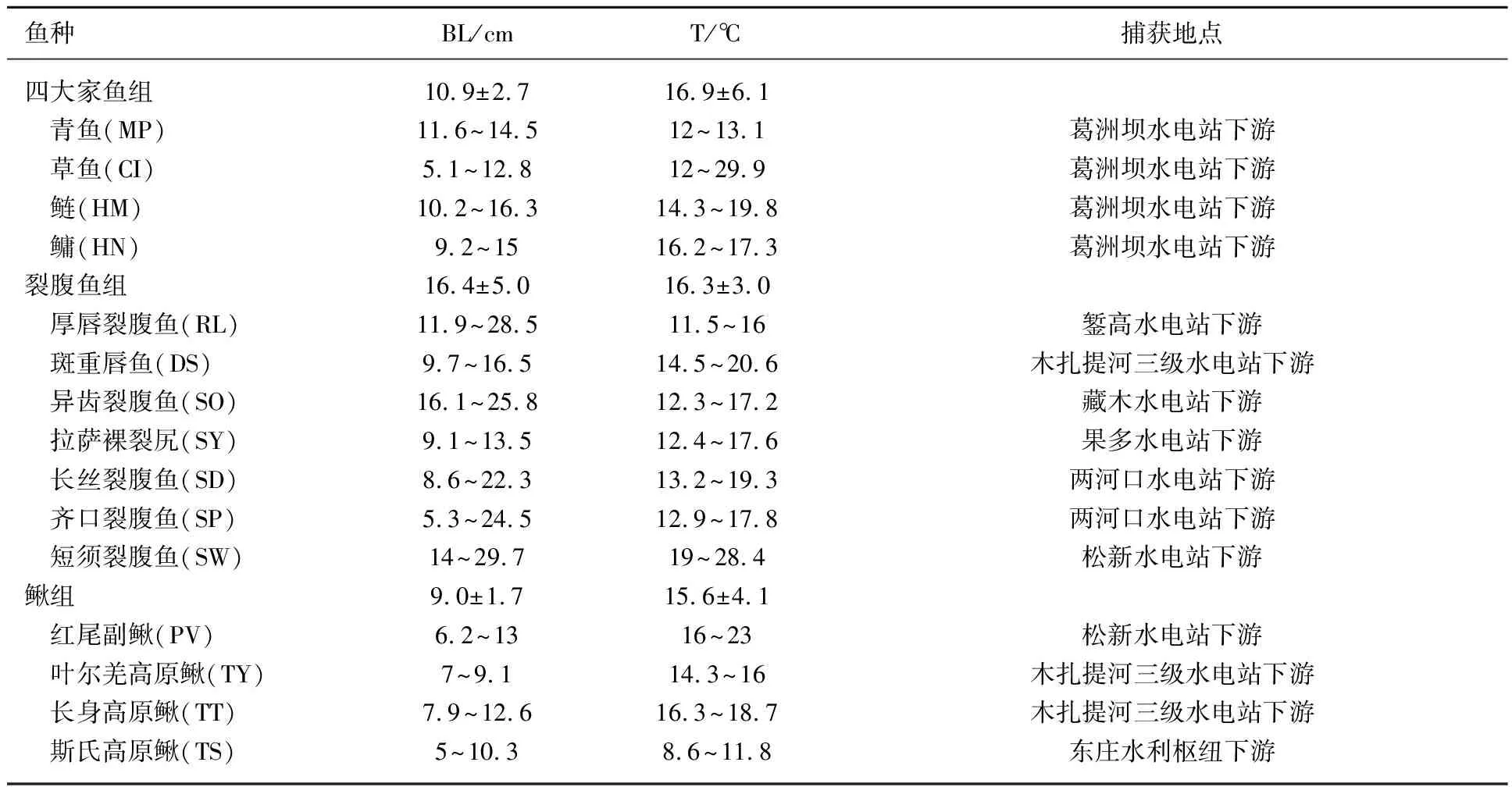
表1 四大家鱼、裂腹鱼和鳅组各鱼种感应流速和爆发游泳速度测试的体长和水温
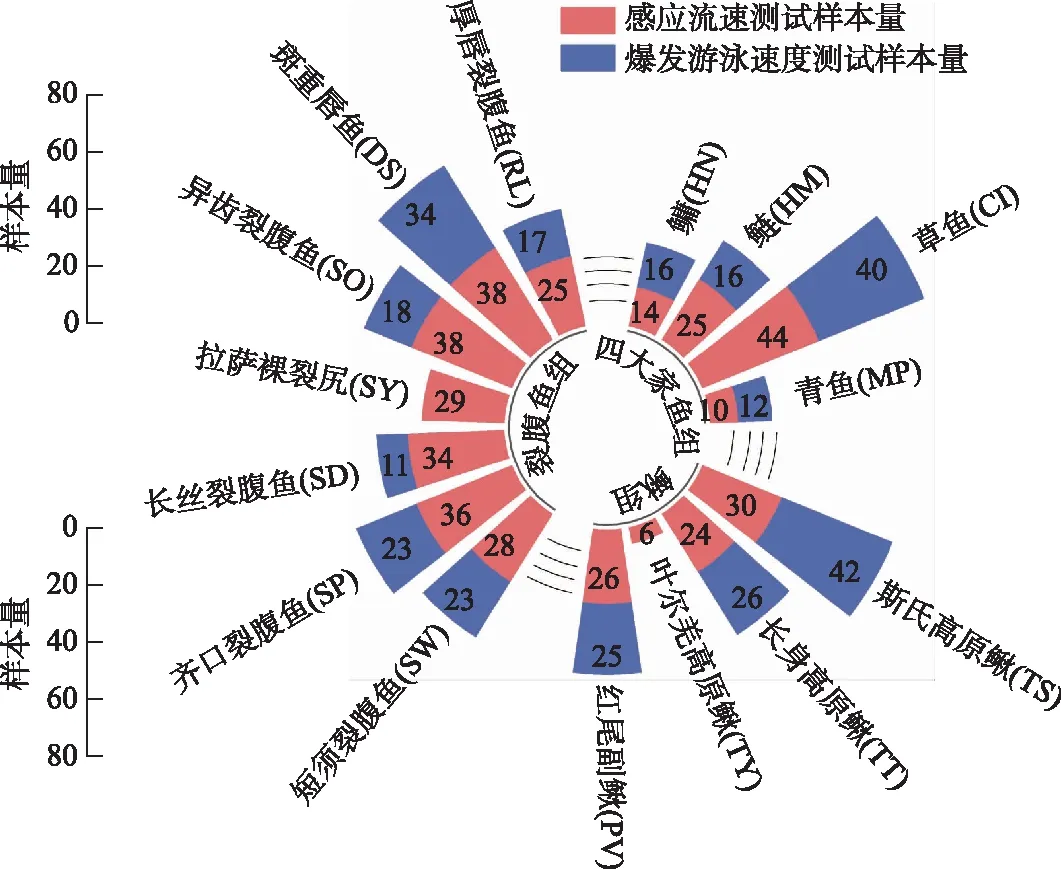
图2 四大家鱼组、裂腹鱼组和鳅组样本量Fig.2 The sample sizes of the four major Chinese carp, Schizothorax fish species and Cobitidae fish species groups
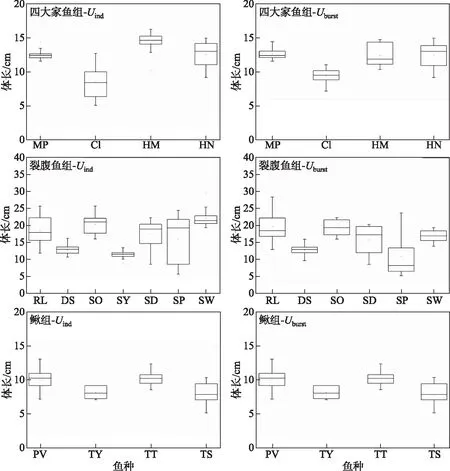
图3 四大家鱼、裂腹鱼和鳅组感应流速和爆发游泳速度测试鱼种体长分布Fig.3 The body length distribution of the four major Chinese carp, Schizothorax fish species and Cobitidae fish species groups tested for Uind and Uburst
1.3 实验方法
1.3.1 感应流速(Uind) 测试前先估测实验鱼的体长和体重,然后将其放入游泳能力测定装置的游泳区中,在静水下适应1 h. 鱼在静水中基本不游动,且头朝向来流的方向. 适应结束后,每隔5 s流速增加0.01 m/s,当实验鱼开始主动朝着来流方向游动,此流速即为该实验鱼的感应流速.
1.3.2 爆发游泳速度(Uburst) 测试前先估测实验鱼的体长,然后将其放入游泳能力测定装置的游泳区中,在0.5 BL/s的流速下适应1 h. 适应结束后,每隔20 s流速增加0.5 BL/s直至实验鱼疲劳. 当实验鱼停靠在下游拦网上时,轻拍下游壁面20 s,鱼仍不重新游动,视为疲劳. 爆发游泳速度的计算方法为[18]:
(1)
式中,Umax表示实验鱼能够完成持续时间Δt的最大游速(BL/s),Δt为流速递增的时间间隔(20 s),t为实验鱼在最高流速下的游泳时间(s),ΔU为流速增量(BL/s).
1.4 数据分析
本研究构建了以体长和水温为协变量,四大家鱼、裂腹鱼和鳅组感应流速和爆发游泳速度为因变量的多元线性回归预测模型. 函数模型表达式为:
(Uind|Uburst)=a0+a1·T+a2·BL
(2)
式中,a0为截距,a1、a2为回归系数,T为水温(℃),BL为体长(cm),Uind为感应流速(BL/s),Uburst为爆发游泳速度(BL/s). 为避免模型中自变量(T和BL)可能存在多重共线性的问题,提高模型精度,从所有可能的模型中采用逐步回归法,选择赤池信息准则(Akaike information criterion,AIC)筛选出最优模型,最优模型的判定依据为模型AIC值最小[19].
利用过鱼对象感应流速和爆发游泳速度值绘制生存分析曲线(Kaplan-Meier曲线),获取不同流速下鱼可感知水流方向后顶流游泳的数量百分比以及鱼在爆发游泳时未发生疲劳的数量百分比. 分别计算90%的鱼可感知该流速并顶流运动所对应的水流速度值和90%的鱼在某流速下爆发游泳未产生疲劳所对应的水流速度值. 本研究统计学数据均采用R软件(R 3.6.0)进行分析,并用Origin 2018软件绘图.
2 结果
2.1 感应流速
对四大家鱼组(幼鱼)、裂腹鱼组(幼鱼或成鱼)和鳅组(幼鱼或成鱼)的感应流速多元线性回归模型进行了筛选,得到的最优模型中协变量均包括体长和水温(表2). 通过分析各鱼组最优模型的回归系数,发现四大家鱼、裂腹鱼和鳅组的感应流速均与体长和水温呈负相关(表3). 以水温为16℃为例,3组鱼感应流速随着体长的增加而减小(图4).
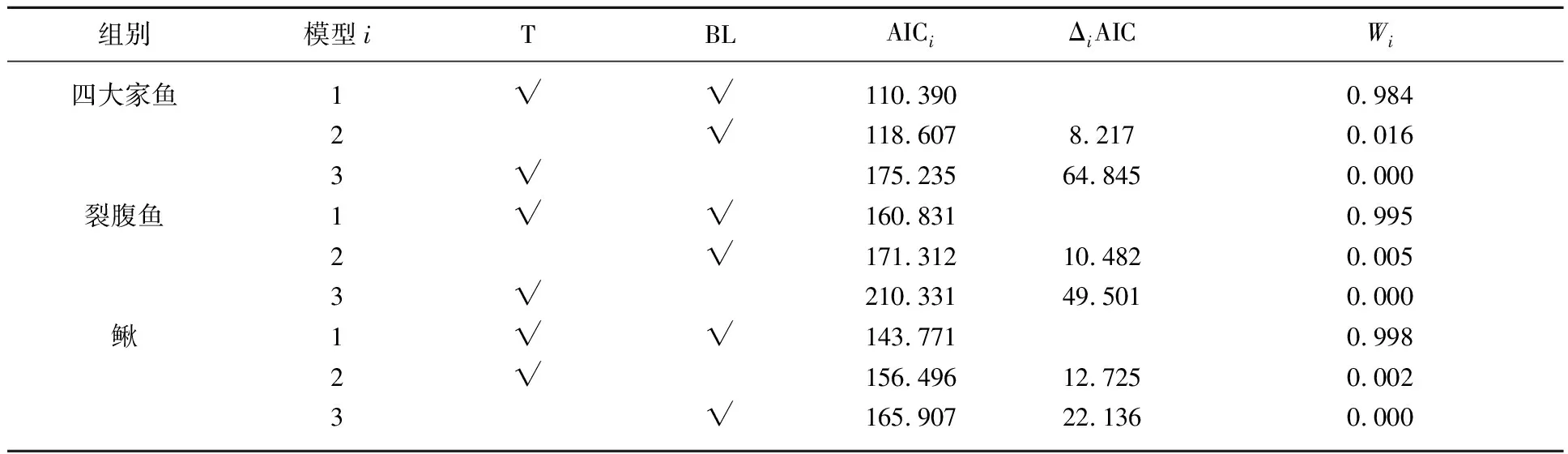
表2 基于赤池信息准则(AIC)的四大家鱼、裂腹鱼和鳅组感应流速多元线性回归模型选择*

表3 四大家鱼、裂腹鱼和鳅组感应流速最优回归模型各协变量回归系数*
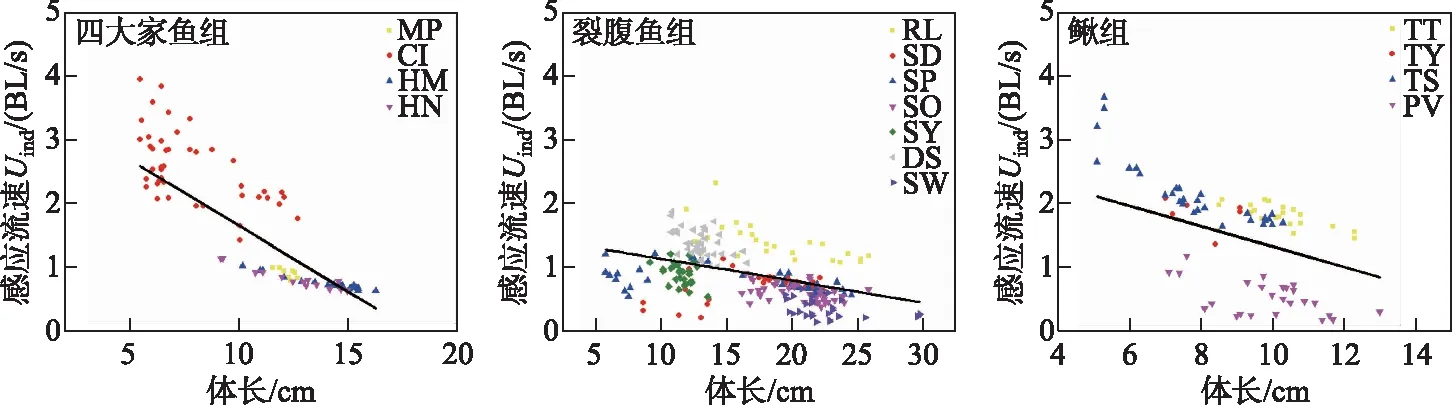
图4 体长对四大家鱼、裂腹鱼和鳅组感应流速的影响(T=16℃) (黑色实线为水温16℃时的感应速度线性回归模型预测线;16℃为各组鱼的测试水温平均值)Fig.4 Effects of body length on Uind of the four major Chinese carp, Schizothorax fish species and Cobitidae fish species groups (T=16℃) (The black solid lines are the prediction lines of the linear regression models of Uind at 16℃; the average test water temperature of each group was 16℃)
2.2 爆发游泳速度
对四大家鱼组、裂腹鱼组和鳅组的爆发流速多元线性回归模型进行了筛选,四大家鱼和裂腹鱼组的最优模型中协变量均包括体长和水温,而鳅组最优模型协变量只有体长(表4). 通过分析各鱼组最优模型的回归系数,发现四大家鱼和裂腹鱼组的爆发游泳速度与体长呈负相关,而与水温呈正相关(表5). 鳅组爆发游泳速度与体长呈负相关(表5). 以水温为16℃为例,3组鱼爆发游泳速度也随体长的增加而减小(图5).
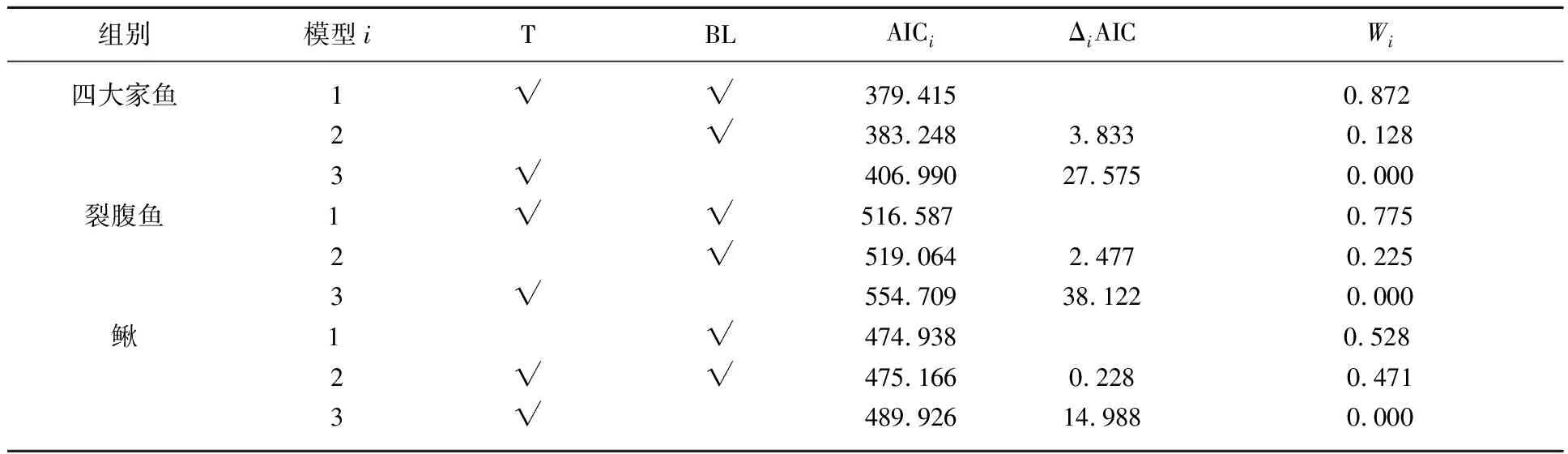
表4 基于赤池信息准则(AIC)的多元线性回归模型选择*
2.3 感应水流百分比及爆发未疲劳百分比与流速的关系
根据实验鱼体长范围,将其幼鱼和成鱼[20-24]分别进行游泳能力统计分析. 通过绘制鱼感应水流和爆发未疲劳的数量百分比的Kaplan-Meier曲线,发现当百分比为90%时,四大家鱼、裂腹鱼和鳅组幼鱼感应流速分别为3.00、1.67和3.22 BL/s(图6),裂腹鱼和鳅组成鱼感应流速分别为1.27和2.09 BL/s(图6);四大家鱼、裂腹鱼和鳅组幼鱼爆发未疲劳的游泳速度分别为8.13 、9.27和9.61 BL/s(图7),裂腹鱼和鳅组成鱼爆发未疲劳的游泳速度分别为7.64和7.77 BL/s(图7).
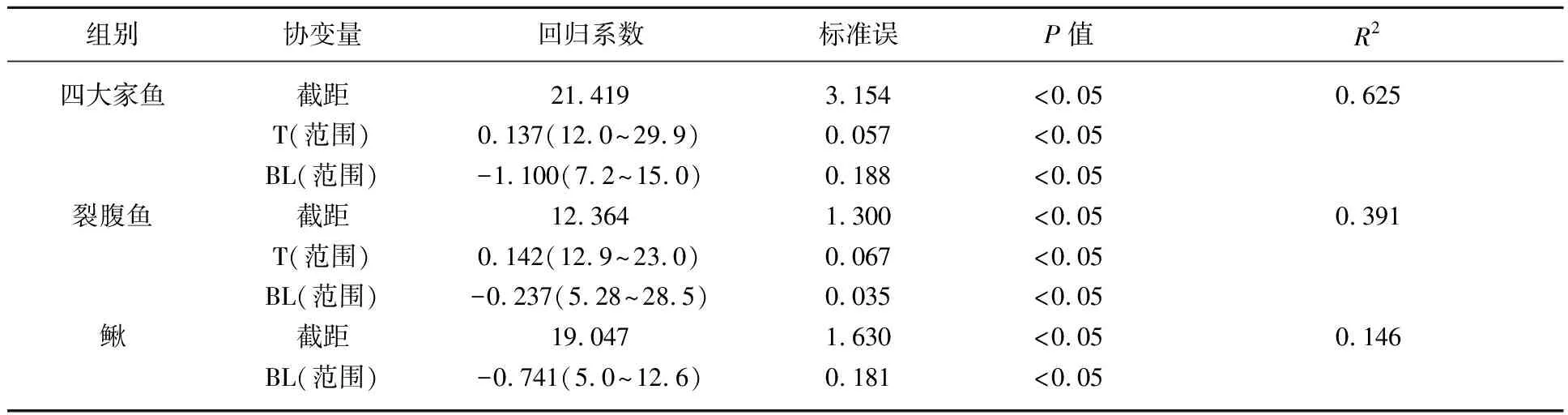
表5 四大家鱼、裂腹鱼和鳅组爆发游泳速度最优回归模型各协变量回归系数*

图5 体长对四大家鱼、裂腹鱼和鳅组爆发游泳速度的影响(T=16℃) (黑色实线为水温16℃时的爆发游泳速度线性回归模型预测线;16℃为各组鱼的测试水温平均值)Fig.5 Effects of body length on Uburst of the four major Chinese carp, Schizothorax fish species and Cobitidae fish species groups (T=16℃) (The black solid lines are the prediction lines of the linear regression models of Uburst at 16℃; the average test water temperature of each group was 16℃)

图6 四大家鱼、裂腹鱼和鳅组幼鱼和成鱼感应流速百分比Fig.6 The percentage of juvenile and adult fish induced to swim (Uind) of the four major Chinese carp, Schizothorax fish species and Cobitidae fish species groups
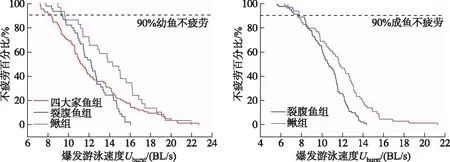
图7 四大家鱼、裂腹鱼和鳅组幼鱼和成鱼爆发游泳时不疲劳百分比Fig.7 The percentage of juvenile and adult fish that were not exhausted (Uburst) of the four major Chinese carp, Schizothorax fish species and Cobitidae fish species groups
3 讨论
3.1 水温及体长对游泳能力的影响
鱼体长是影响鱼类游泳能力的重要因素之一. 本实验3组鱼的感应流速与爆发游泳速度均随体长的增加而下降(表3,表5). 红河马堵山水电站3种过鱼对象(暗色唇鲮、华南鲤和红魾)和汉江旬阳坝8种过鱼对象的感应流速和爆发游泳速度与体长表现出负相关性[18,25]. 此现象应与游泳能力随体长变化为异数生长关系有关,即随着鱼体尺寸的增大,鱼体的表面积和肌肉重量的相对比例减小,导致随鱼体长的增加,鱼的有氧及无氧游泳能力反而下降[26]. 本研究中四大家鱼组未包括成鱼,但实验对象的感应流速与体长也呈负相关,与白艳勤等[27]关于草鱼幼鱼和鲢幼鱼相对感应流速与体长呈负相关的研究结果一致.
水温是影响游泳能力的另一个重要环境因子. 研究结果表明水温与3组鱼的感应流速均为负相关. 可能是由于水温下降导致了个体的活跃性降低. 如当水温从28℃下降至22℃时,表征大斑马鱼(Devariomalabaricus)个体活跃性的摆尾频率显著降低[28]. 在较低水温下,肌肉组织中三磷酸腺苷和葡萄糖均减少,当水温从15℃下降至5℃时,底鳉鱼(Fundulusheteroclitus)红肌中的三磷酸腺苷从1.72 μmol/g下降至1.38 μmol/g,葡萄糖从1.93 μmol/g下降至1.34 μmol/g,临界游泳速度显著降低[29]. 温度下降还会使有氧肌肉纤维中酶活性降低,如罗非鱼(Oreochromisniloticus)的肌酸磷酸激酶从20.5℃时的176 mmol/(g·min)升至26.5℃ 时的202 mmol/(g·min)[30],这都将导致低温下鱼的感应流速增加.
爆发式游泳是利用无氧代谢产生的高能量完成冲刺、快速启动等过程,因此运动时间较短,运动后迅速疲劳[31-32]. 四大家鱼与裂腹鱼组的爆发游速与温度为正相关关系(表5). 较多研究表明,水温对有氧运动有影响,但对无氧运动影响不大[33-34]. 本研究中水温升高使鱼的爆发游泳速度增大,与此结论不符. 以18 cm体长的裂腹鱼组为例,根据裂腹鱼的爆发游泳速度预测模型(Uburst=12.364+0.142T-0.237BL)可得,水温从12℃增加至17℃时,爆发游速由18.33 BL/s增加至19.04 BL/s. 这可能与前期的有氧运动有关或者由其他因素影响造成,还需进一步研究. 然而,鳅组的爆发游速与水温无显著相关性(P>0.05). 视频观察发现,多数鳅科鱼类在高流速下利用腹部紧贴水槽底部或侧面,节省游泳能量消耗,抵抗更高的水流速度. 可见,这种腹部吸附行为基本不受温度的影响,仅在高流速才表现. 腹部吸附行为也显著提高了鳅的爆发游泳能力,根据3组鱼各自的爆发游泳速度预测模型(表5)可知,在相同水温(16℃)下,平均体长为10 cm的鳅科鱼的爆发游速为26.46 BL/s,相近体长的四大家鱼为12.61 BL/s,裂腹鱼组为12.27 BL/s. 鳅组的爆发游泳速度数据较另外两组分散,预测模型确定系数偏低,这是因为有部分鳅在高流速下未采用腹部吸附行为抵抗高速水流,导致部分鳅爆发游泳速度偏低.
3.2 过鱼设施内流速设计建议
过鱼对象的游泳能力或行为数据可为过鱼设施进口、池室内和出口的设计提供重要信息. 对于上行过鱼设施设计,若入口处的设计流速太低,不足以对鱼产生方向性刺激[14],则鱼将找不到入口;但若入口流速超过了爆发游泳速度,又将成为鱼进入过鱼设施的阻碍. 为了吸引距离更远的鱼能感应到过鱼设施入口的水流,入口流速应该尽量高些,但上限不应超过爆发游泳速度. 过鱼设施池室内的流速应高于感应流速,以避免鱼在上溯过程中迷失方向. 根据感应流速生存曲线(图6),为保证90%的过鱼对象感知水流且朝正确方向洄游上溯,当以裂腹鱼成鱼为过鱼对象时,建议池室内流速大于1.27 BL/s,以鳅科成鱼为过鱼对象时,池室内流速大于2.09 BL/s. 鱼道内流速不应超过过鱼对象的爆发游泳速度[35]. 根据爆发游泳速度的生存曲线,为保证90%的过鱼对象不疲劳(图7),当以裂腹鱼成鱼为过鱼对象时,建议池室内最大流速不能超过7.64 BL/s,以鳅科成鱼为过鱼对象时,建议池室内最大流速不能超过7.77 BL/s. 上行过鱼设施的出口通常设置在远离坝址的上游处,若出口流速过小,低于感应流速,则会导致鱼迷失方向甚至返回过鱼设施内,导致过鱼设施通过效率下降. 因此,上行过鱼设施出口处的主流流速建议大于过鱼对象的感应流速.
下行过鱼设施主要考虑幼鱼下行过坝需求,幼鱼下行主要采用水流引导的方式使鱼主动进入下行过鱼设施入口,若入口流速超过鱼的爆发游泳速度,幼鱼的游泳方向将改变为与水流方向相反,逆流向上游逃逸,若因水流的速度过快,造成被动下行,则会导致身体损伤[15]. 当以四大家鱼幼鱼为下行过鱼设施的过鱼对象时,根据爆发游泳速度的生存曲线,建议过鱼设施的入口最大流速不能超过8.13 BL/s. 当以裂腹鱼幼鱼为过鱼对象时,建议下行过鱼设施的入口最大流速不能超过9.27 BL/s. 以鳅科成鱼为过鱼对象时,建议下行过鱼设施的入口最大流速不能超过9.61 BL/s.
由于野外实验条件的限制,所选择的实验对象并未涵盖整个生命周期,如四大家鱼,因此过鱼设施流速建议值有一定的局限性,将其应用于过鱼设施设计时,需在原型过鱼设施中进行校正及优化. 此外,游泳能力测试的流场环境是均匀且单一的,真实的过鱼设施流场环境具有复杂性. 过鱼对象游泳能力并不能完全作为评判过鱼设施设计的唯一标准. 过鱼设施的局部区域(竖缝或孔口)的设计流速大小与游泳能力有关,然而鱼在过鱼设施内部的游泳选择往往是对多个水力因子(紊动能、紊动强度、雷诺切应力、耗散率和涡)耦合作用的行为反馈[36-37]. 因此亟需开展鱼在复杂流场中的游泳行为研究.
