盐处理及套袋处理对‘摩尔多瓦’葡萄果实品质及花色苷合成相关基因的影响
2022-09-03田淑芬热汗古丽艾海提王超霞
孙 洋,田淑芬,热汗古丽·艾海提,王超霞,王 荣,刘 丹
(1.天津农学院园艺园林学院,天津 3003842;2.天津市尖峰天然产物研究开发有限公司,天津 300192)
花色苷作为类黄酮途径的次级代谢产物,其生物合成由结构基因和转录因子协同调控。花色苷合成途径中的苯丙氨酸解氨酶(Phenylala nineammonialyase,PAL)、类 黄 酮-3’-羟 基 化 酶(Flavonoid 3’-hydroxylase,F3’H)、类黄酮-3’-5’-羟基化酶(Flavonoid 3’-5’-hydroxylase,F3’5’H)、无色花色素还原酶(Leucoanthocyanidin Reductase,LAR)、无色花色素双加氧酶(Leucoanthocyanidin Dioxygenase,LDOX)、类黄酮3-O-葡萄糖基转移酶(UDP-glucose Flavonoid Glucosyltransferase,UFGT)等都直接催化花色苷的合成。除此之外,转录因子通过与靶基因启动子上的特定元件结合,调节靶基因的转录水平,从而间接调控花色苷的合成。
葡萄果实中花色苷的合成,与品种特性有关,由品种基因型决定,也易受外界光照、土壤、温度等栽培环境的影响。天津位于环渤海产区的中心,土壤盐渍化问题严重,研究表明,适度的盐胁迫处理可以改善葡萄果实品质。50 mmol·L的NaCl 浓度提高了果实发育后期的固酸比和可溶性固形物含量。根系的局部盐处理提高了果实风味及果实中的香气物质的种类和浓度。盐溶液喷施叶面可以提高‘ 巨峰’葡萄的成熟果实的花色苷及可溶性固形物。对酿酒葡萄‘ 赤霞珠’果粒喷施过高浓度盐溶液会延缓花色苷的积累。
近几年来,套袋技术在葡萄生产栽培中广泛应用。套袋处理能有效地降低果实表面的农药残留,调节果实色泽的形成,不同程度地影响了光照条件、果实着色面积、果面光洁度及果品安全性。套袋可提高葡萄果实的可溶性糖、可滴定酸、VC 等内含物质的含量。高献亭等试验证明,套袋果实第二发育期时间较长,依据果实发育规律,从肥水、负载量、病虫防治几个方面加强管理,可促进果粒增大。倪志婧等研究证明,葡萄果实中花色苷的质量比随着遮光率增加而降低;马宗桓等研究发现,与透光率50%,15%,5%,0%对比下,不套袋的对照组花色苷含量最高,随着‘ 马瑟兰’的成熟,差异逐渐显著。紫外线照射可以促进合成花色苷相关基因UFGT 的表达,从而使花色苷含量增加。Ubi 等发现,紫外线可以促进基因的表达,而UFGT 是花色苷合成的主要基因之一;Zhang研究发现,UV-C 照射葡萄果实可以诱导莽草酸代谢途径中 VvDAHPS -1 和VvDAHPS-2 基因的表达,从而导致花色苷的含量增加。
‘ 摩尔多瓦’葡萄耐寒耐旱,适于在四季分明、干旱多风、土壤盐渍化的天津地区进行种植和栽培。本研究通过浇灌盐溶液、套袋等措施改变土壤盐分和光照等栽培环境,研究盐溶液、套袋处理对花色苷合成的分子调控机制的影响,为培育高含量花色苷葡萄新品种及提高葡萄附加值提供理论基础。
1 材料和方法
1.1 试验材料
本研究于2020 年7 月5 日—9 月25 日在天津市武清区金锅农业生态园进行(E117.13°,N39.38°),该园区为葡萄种植基地核心示范区,农户已连续种植葡萄10 余年,管理经验丰富。所用的‘ 摩尔多瓦’葡萄(Moldova)为多年生自根苗,采用户外棚架栽培,定植株距1.5 m,棚架高4 m。每株葡萄结果主蔓1 个,每个结果枝保留1 个果穗,整株葡萄果穗保持40 穗。
自7 月5 日果实转色前1 周开始处理,试验设置3 个处理组,具体处理方法见表1。每个处理10株树,每3 株作为1 个重复,共3 个重复。每10 d 处理1 次,共处理5 次。在第3 次处理后,于转色后期(8 月5 日)、第2 次膨大期(8 月25 日)及成熟期(9月25 日)采样,共采样3 次。采样时随机挑选同一结果部位、无病虫害及机械损伤的果穗,从果穗上、中、下采集果实混合。其中用于花色苷及其相关基因表达转录水平检测的样品葡萄剥皮后,将果皮用锡箔纸包裹好放入液氮速冻,并带回实验室保存在-80 ℃冰箱备用。

表1 处理方法
1.2 检测指标及测定方法
1.2.1 可溶性固形物的测定 按照不同处理不同采样时间随机取出30 个完整果粒用于可溶性固形物的检测。用数显折光仪(陆恒生物)测定可溶性固形物的含量。
1.2.2 总酚含量的测定‘摩尔多瓦’葡萄果皮中总酚含量的测定采用可见光分光光度计(Mapada,p6)来测定:将存储在-80 ℃冰箱里的葡萄果皮用总酚提取试剂盒(购于南京建成生物工程研究所)进行测定。每个处理重复3 次。
1.2.3 总花色苷的测定 葡萄果皮中的总花色苷含量的测定采用pH 示差法来测定:将存储在-80 ℃冰箱里的葡萄果皮用花色苷提取试剂盒(购于北京盒子生工科技有限公司)进行测定。每个处理重复3 次。1.2.4 总RNA 的提取及定量方法 选取葡萄花色苷合成代谢途径中6 个相关酶基因:PAL、F3’H、F3’5’H、LAR、LDOX、UFGT。根据NCBI 查找的参考基因序列,采用Primer5 软件设计试验中的基因引物序列(表2)。
用RNAprep Pure Plant Plus Kit 试剂盒(DP441,天根)提取葡萄果皮总RNA,反转录后得到cDNA。按照SuperReal PreMix Plus(SYBR Green)的操作指导,在Illumina-Eco 荧光定量PCR 仪(美国ABI)上测定基因的表达量。
以延伸因子EF-1α 为内参基因进行荧光定量PCR,采用Primer5 软件设计相关引物(表2)。10 μL反应体系:2×SuperReal PreMix Plus 5 μL、上下游引物 混 合 物(10 mM)0.3 μL、cDNA 模 板0.4 μL 和RNase-free 水4.3 μL。反应条件:95 ℃15 min;然后,95 ℃10 s,56 ℃20 s,72 ℃30 s,共40 个循环;最后,进行溶解曲线测定。荧光定量结果通过相对定量法(2)进行分析,结果以3 次生物学重复的平均值±标准差表示,采用Excel 软件绘图。

表2 引物信息
1.3 数据整理
数据方差和差异显著性(P<0.05)分析利用Excel 和SPSS 软件。绘图采用Excel 和Origin 软件。
2 结果与分析
2.1 不同处理对‘摩尔多瓦’葡萄果实品质的影响
2.1.1 盐和套袋处理对‘摩尔多瓦’葡萄可溶性固形物的影响 葡萄从转色期到成熟期间,B 处理和C处理对摩尔多瓦葡萄可溶性固形物含量变化情况如图1 所示。由图1 可见,与采样初期相比,随着‘ 摩尔多瓦’葡萄的成熟,A、B、C 3 个处理组果实的可溶性固形物含量呈上升趋势,9 月25 日最后1 次采样的果实可溶性固形物含量显著高于8 月5 日采样初期。尤其是从8 月25 日开始,各组可溶性固形物含量迅速上升,成熟期B 处理和C 处理的可溶性固形物含量均显著高于A 处理(对照),B 处理可溶性固形物含量最高。结果表明,B 处理和C 处理有利于‘ 摩尔多瓦’葡萄可溶性固形物的积累。
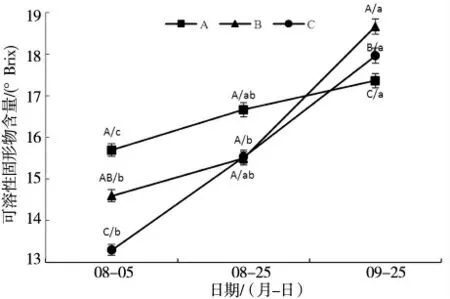
图1 盐和套袋处理在不同时期对‘摩尔多瓦’葡萄可溶性固形物的影响
2.1.2 盐和套袋处理对‘摩尔多瓦’葡萄可滴定酸的影响 随着‘ 摩尔多瓦’葡萄果实的成熟,不同处理果实可滴定酸含量变化情况如图2 所示。从转色期到成熟期,A、B、C 处理的果实可滴定酸含量整体呈现持续下降的趋势。结合葡萄物候期分析,8 月25日之后,葡萄进入成熟期,开始增糖降酸,B 处理组果实可滴定酸含量下降速度高于A 处理组,到果实成熟后期,B 处理组可滴定酸含量显著低于A、C 处理组。结果表明,套袋处理及盐处理有利于可滴定酸含量的降低。

图2 盐和套袋处理在不同时间对‘摩尔多瓦’葡萄可滴定酸的影响
2.1.3 盐和套袋处理对‘摩尔多瓦’葡萄总酚含量的影响 图3 可知,随着果实成熟处理C 总酚含量升高,处理A、B 总酚含量显著降低,处理B 下降速度最快,且成熟期总酚含量最低,显著低于对照。这说明套袋处理及盐处理有利于葡萄果实中总酚含量的降低,推测与花色苷含量下降有关。
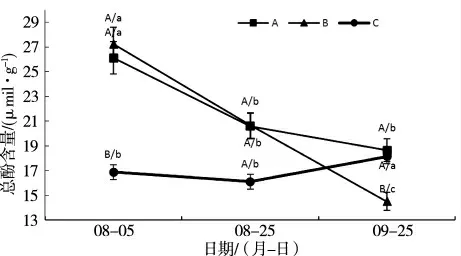
图3 盐和套袋处理对‘摩尔多瓦’葡萄果皮中总酚的影响
2.2 不同处理对‘摩尔多瓦’葡萄果皮中花色苷合成的影响
2.2.1 盐和套袋处理对‘摩尔多瓦’葡萄花色苷含量的影响 由图4 可以看出,随着‘ 摩尔多瓦’葡萄的成熟,各组果皮花色苷的含量逐渐升高,成熟期不同处理果实中总花色苷含量差异不显著。结果表明,B和C 处理不利于花色苷含量积累。由此可见,光照对花色苷积累的重要性。

图4 盐和套袋处理对‘摩尔多瓦’葡萄果皮中总花色苷的影响
2.2.2 ‘摩尔多瓦’葡萄中花色苷合成相关基因的相对表达量 PAL、F3’H、DFR、F3’5’H、LDOX 和UFGT这6 种结构酶基因是花色苷生物合成过程中的关键基因。图5 是本试验B 和C 处理对‘ 摩尔多瓦’葡萄果皮中花色苷相关基因转录表达水平的影响研究结果。
从图5 可以看出,与第1 次采样相比,随着果实的成熟,C 处理组F3’H、DFR 和F3’5’H 基因的相对表达量上调,UFGT 基因的相对表达量下调,PAL和LDOX 的相对表达量没有明显的变化。B 处理组的PAL、DFR、F3’5’H 和UFGT 基因的相对表达量均明显下调,F3’H 和LDOX 基因的相对表达量没有明显的变化。A 处理组的LDOX 和UFGT 基因的相对表达量先上调后下调,PAL、F3’H、DFR 和F3’5’H基因的相对表达量均下调。结果表明,B 处理和C 处理抑制花色苷合成相关基因LDOX 和UFGT 的表达,不利于无色花青素向有色花色素的转变以及有色花色素与各种单糖结合形成花色苷。

图5 盐和套袋处理对‘摩尔多瓦’葡萄果皮中花色苷相关基因转录表达水平的影响
3 结论与讨论
植物在生长发育的过程中经常会受到一些非生物的胁迫,如:温度、盐、紫外线、干旱等,盐就是其中的一类。盐不仅影响着植物的生产过程,同时大大影响着产品的丰产与稳定。用盐处理葡萄会影响它的渗透性,使得葡萄体内失水,减弱光合作用和呼吸作用的能力,抑制葡萄的正常生长发育与成熟。研究表明,只有低盐处理可以使得果树果实品质下降,说明盐处理对果实品质的影响具有浓度效应。本研究盐处理组‘ 摩尔多瓦’葡萄可溶性固形物和总花色苷含量的增长速度缓慢,可滴定酸含量下降不明显,总酚含量明显低于对照组,从而证实了3‰盐处理通过影响葡萄的可滴定酸、可溶性固形物、总酚、花色苷的含量来影响葡萄果实的品质,不利于果实品质的提高。
盐当中的金属离子对花色苷有一定的辅色效果。Na的大量积累可以提高果实的着色。Galli 等研究发现,在草莓果实中,Na含量增加3 倍使得UFGT 等着色相关基因的大幅度上调,而当Na增加6 倍时对葡萄果实的品质和着色相关基因没有显著影响或显著下调了UFGT 的表达,暗示了不同盐水平对葡萄品质形成和基因调控具有不同的效应。孙红等研究表明,低盐处理对提高花色苷有显著影响,而高盐处理则不利于着色。本研究盐处理的‘ 摩尔多瓦’ 葡萄从转色期到成熟期花色苷的含量虽然也在不断上升,但一直低于对照组,且合成速度较慢。这说明3‰盐浓度还是较低,对花色苷合成的影响不显著。
光照是决定果实品质的主要因素之一,它通过光合作用,影响库源关系来实现对葡萄代谢的影响。前人研究紫外线对果实品质影响的结果表明,在葡萄果实的生长发育过程中,紫外线照射并没有影响果实的外在品质、可溶性总糖和有机酸,但显著影响了果实的内在品质,且在成熟期降低了VC 含量。张珍珍等试验证明100%转色至完全成熟期间采用50%遮光的遮阳网处理,显著提高了样品中甲基化花色苷含量(甲基化花色苷的稳定性较强)。本试验中随着葡萄的成熟,套袋处理的可溶性糖的含量上升较快,而花色苷含量低于对照组,可滴定酸含量和总酚的含量下降趋势都比对照组慢。套袋减少了葡萄果实接受光照的强度,减少空气流通等带来的水分蒸发,改变了果穗周围的温湿度等微气候环境,但不利于花色苷的积累。
花色苷不仅是类黄酮代谢途径的重要产物,也是葡萄中重要的一类酚类物质。它的积累过程中会受到逆境胁迫的影响,光照是其中的影响因素之一。前任研究发现,紫外线和红外线对植物生长影响较大。紫外线可根据波长分为UV-A(315~390 nm)、UV-B(280~315 nm)、UV-C(小于280 nm)。UV 福射对类黄酮合成的调节主要是在其基因转录水平上进行的。Ubi等研究发现,紫外线能促进CHS、F3H、DFR、ANS、UFGT 基因的表达,进而增加花色苷的积累。Matus 等研究发现,在葡萄转色期增加果穗的受光面积能够影响花色苷的合成。由此可见,光主要诱导原花色素的合成并影响其组成,而紫外线主要诱导黄酮醇的生物合成。本研究结果表明,套袋处理不利于‘ 摩尔多瓦’葡萄花色苷含量的积累,间接表明紫外线等光照条件对花色苷合成的重要性。
影响花色苷合成的因素除光照和盐胁迫外还有温度和水分胁迫。Pastore 等研究表明,低温处理会在一定程度上促进PAL 表达,进而促进花色苷的含量,经验证,低温条件下的葡萄总花色苷含量是高温处理的2 倍。Yang 等通过转录组和代谢组学分析,表明水分胁迫可以促进CHS、F3H、3GT 等一般生物合成基因的表达,进而使花色苷和合成花色苷的物质含量升高。
此外,现阶段关于花色苷的研究很多,有着广泛的应用前景。花色苷的种类多,稳定性差,影响其合成的因子及其调控机制复杂,花色苷合成决定的酶学与分子机制还不够完善。花色苷的生理活性功能广受人们关注,由于国内合成工艺的限制,花色苷的纯度较低,无法满足人们的需求。因此,研究葡萄中花色苷合成调控机制,可为培育高含量花色苷新品种提供理论基础。
本研究通过对‘ 摩尔多瓦’葡萄果实果皮中花色苷合成相关基因的转录表达水平、可滴定酸、可溶性固形物、总花色苷含量以及总酚含量等一系列生理指标的检测结果表明,套袋处理有利于葡萄果实可溶性固形物含量的增加,不利于果皮花色苷的合成和积累。盐处理不利于葡萄果实可溶性固形物含量的增加,以及果皮花色苷的合成和积累。
