玉米S1Fa-like 转录因子基因家族的鉴定与表达分析
2022-09-03赵东波高建胜郭建军李拥军郭良海
赵东波,高建胜,郭建军,李拥军,薛 鑫,郭良海
(1.德州市农业科学研究院,山东 德州 253015;2.德州学院,山东 德州 253023)
转录因子通过激活或者抑制相关基因的表达,提高植物的非生物胁迫耐受性。转录因子S1Fa 最早从菠菜中发现,是含有70 个氨基酸的小肽,负调控rps1 表达,其保守结构域包含核定位信号和特定的DNA 结合位点,在双子叶植物和单子叶植物之间高度保守,主要在根和黄化幼苗等组织中表达。S1Fa-like TFs(transcription factors)的数量在不同的植物种类中差异很大。大多数植物不超过5 个S1Fa-like 家族成员;然而花生基因组中含有126 个S1Fa-like 转录因子,且被鉴定为膜结合型转录因子。越来越多的研究发现,S1Fa-lik TFs 响应植物的非生物胁迫,如甘蓝型油菜(Brassica napus)中S1Fa-like响应盐胁迫刺激;棉花(Gossypium hirsutum)中S1FalikeTFs 大多数成员受非生物胁迫刺激表达量下调;杨树(Populus trichocarpa)中PtS1Fa1 和PtS1Fa2 的表达受到干旱和盐胁迫的抑制,且响应ABA、MeJA、SA等激素刺激,其中PtS1Fa2 通过增加抗氧化剂活性,减少活性氧(ROS)积累,进而增强杨树的耐旱性。
植物在生长发育过程中必须应对非生物胁迫,如土壤盐度、干旱和极端温度。然而,关于玉米S1Fa-like TFs 生物信息学、组织表达及胁迫调控表达模式的研究未见报道。本研究采用生物信息学方法,分析玉米S1Fa-like TFs 基因结构、染色体定位、蛋白理化性质、进化关系及组织表达和胁迫表达模式,通过qRT-PCR 技术检测玉米S1Fa-like TFs 受热胁迫时的表达量,为深入解析玉米S1Fa-like TFs在响应非生物胁迫应答中的生物学功能奠定基础。
1 材料和方法
1.1 基因家族的鉴定、蛋白质理化性质及亚细胞定位分析
从PlantTFDB(http://planttfdb.gao-lab.org/)查询S1Fa-like TFs 成员,并利用Pfam(http://pfam.xfam.org/)对得到的蛋白序列进行保守结构域验证。利用Expasy 中的ProtParam(https://web.expasy.org/protparam/)工具获取蛋白质理化性质信息,Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)进行蛋白亚细胞定位分析。
1.2 家族蛋白进化和二级结构预测分析
通过PlantTFDB 获取单子叶植物(大麦、玉米、籼稻、粳稻、高粱、小麦、乌拉尔小麦)和双子叶植物(猕猴桃、拟南芥、油菜、大豆、棉花、杨树、菠菜)共14 种作物的S1Fa-like TFs 蛋白序列,利用MEGA7.0软件中最大似然法(Maximum Likelihood,bootstrap=1000)构建系统发育树。利用在线网站SOMPA(http://www.prabi.fr/)预测蛋白二级结构。
1.3 保守基序、基因结构及启动子分析
利用在线工具MEME(http://memesuite.org/)对家族蛋白进行motif 分析,预测基因家族成员的保守结 构 域。使 用PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线预测ZmS1Fas启动子区域(ATG 上游2 000 bp)顺式调控元件。
1.4 基因组织表达和诱导表达模式分析
从MaizeGDB qTeller 数据库下载正常组织部位的表达数据(登录号PMC5808982),NCBI 数据库下载受干旱胁迫根部位转录组数据(登录号SRP052697)和不同玉米材料苗期受温度胁迫地上部位转录组数据(登录号SRP106663),利用TBtools软件中HeatMap 进行可视化处理。
1.5 qRT-PCR 检测基因家族成员热胁迫表达量
玉米自交系B73 和Oh43 材料,在16/8 h 光照培养室长日照条件下培养至3 叶期(14 d)。玉米苗被置于42 ℃培养箱中处理4 h,对照则正常条件培养,每个样本处理6 株幼苗。收集幼苗地上部分,液氮中研磨,提取RNA,反转录之后以ZmActin1 为内参,检测ZmS1Fas 胁迫处理前后的相对表达量,并采用GraphPad Prism 软件作图。所用引物序列见表1。
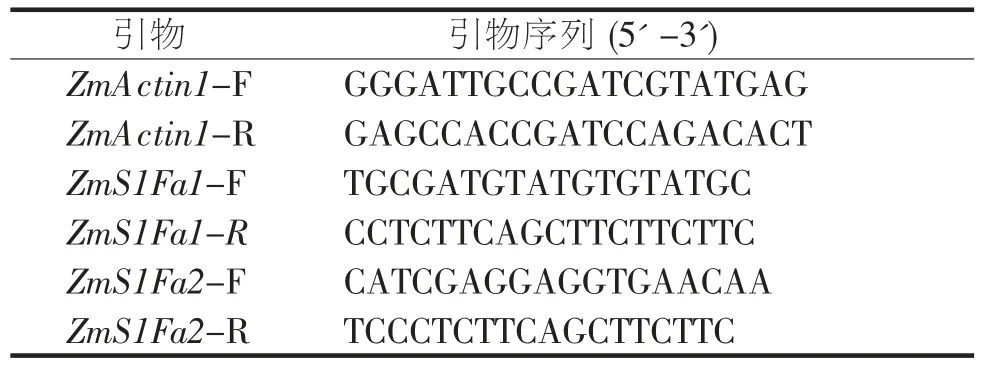
表1 荧光定量PCR 引物
2 结果与分析
2.1 ZmS1Fas 的鉴定、理化性质、亚细胞定位分析及蛋白二级结构预测
从PlantTFDB 获取S1Fa-like 家族成员蛋白序列,经Pfam 检验,结果显示(图1-C),家族成员均含有S1FA 或S1FA superfamily 保守结构域。根据染色体位置命名为ZmS1Fa1、ZmS1Fa2(图2),ZmS1Fa1位于2 号染色体、ZmS1Fa2 位于10 号染色体。ZmS1Fas 共编码5 个蛋白(4 种不同蛋白,表2),蛋白长度为56~80 个氨基酸;分子量在6 310.77~8 867.10 Da 之间;等电点在9.82~10.25 之间,均为碱性蛋白;不稳定系数在33.18~37.49 之间,均小于40,属于稳定蛋白;脂肪指数在101.00~105.27 之间,具有良好的热稳定性;亲水性系数在-0.049~0.070 之间,表明ZmS1Fas 家族成员亲水性氨基酸和疏水性氨基酸基本持平;亚细胞定位结果显示,ZmS1Fas 家族成员均定位在细胞核内。蛋白二级结构分析结果表明(表3),其α-螺旋、延长链、β-转角、无规则卷曲等蛋白质二级结构在ZmS1Fas 基因家族成员中均有分布且占比差别不大。
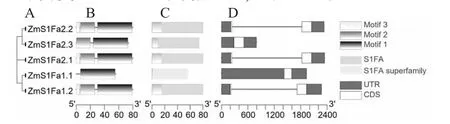
图1 ZmS1Fas 系统进化树(A)、保守基序(B)、保守结构域(C)和基因结构(D)
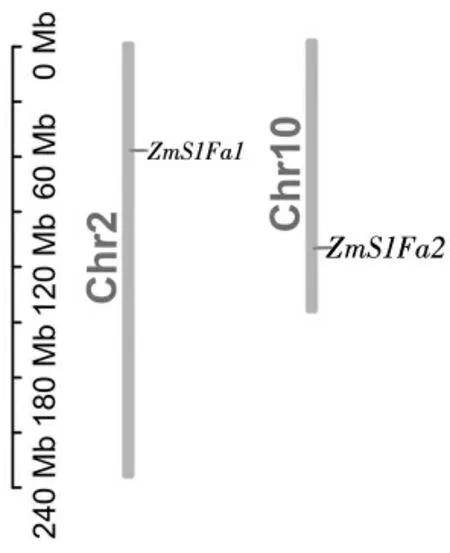
图2 ZmS1Fas 基因染色体定位
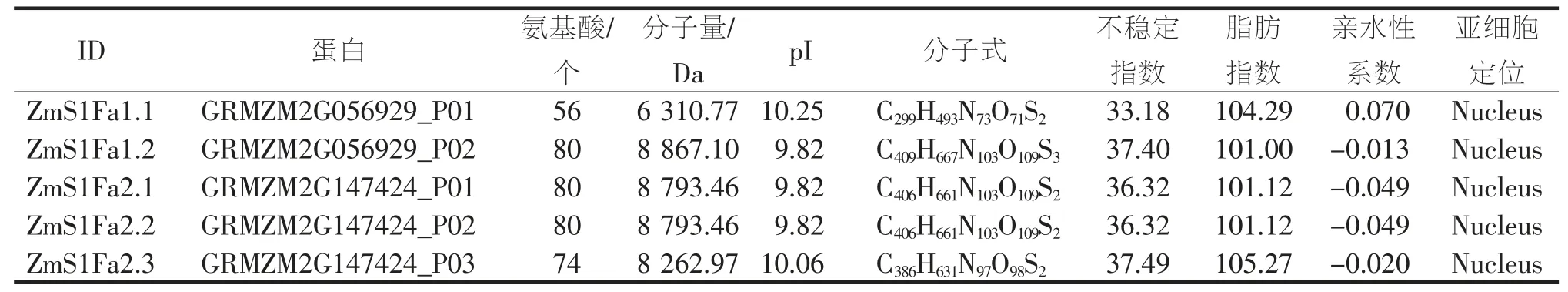
表2 ZmS1Fas 蛋白理化性质
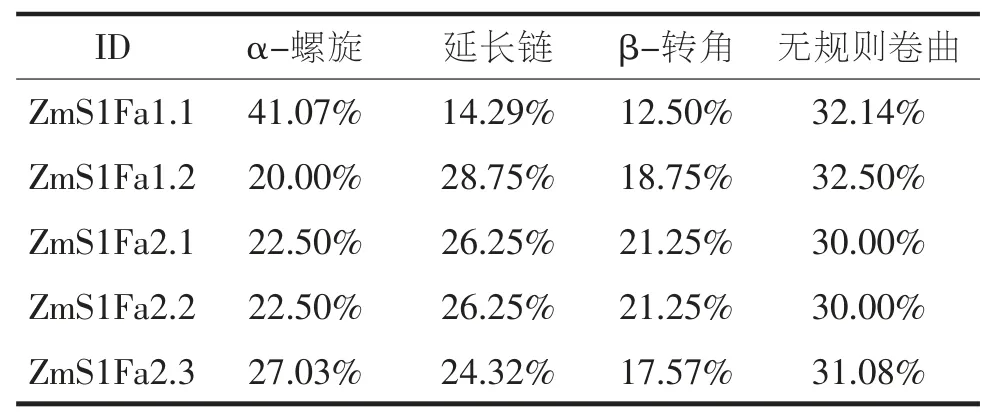
表3 ZmS1Fas 蛋白二级结构
2.2 ZmS1Fas 系统进化、保守基序及基因结构分析
将单子叶和双子叶植物共14 种作物的51 个S1Fa-like 蛋白氨基酸序列进行多重比对,并构建系统进化树(图3)。结果表明,单子叶植物与双子叶植物被分开,说明S1Fa-like 家族快速扩增可能发生在单子叶植物与双子叶植物分化之前。ZmS1Fas 和高粱S1Fa-like 家族成员具有较高的亲缘关系。对ZmS1Fas 蛋白进行motif 分析,结果(图1-B)显示:ZmS1Fas 保守基序较简单,均含有motif1(含50 个氨基酸),占ZmS1Fas 蛋白长度的62.5%~89.3%,推测其为ZmS1Fas 基因家族成员的核心功能区域。ZmS1Fas 的基因结构相对保守(图1-D),均含有1~2 段外显子,0~1 段内含子,且均具有5'UTR 和3'UTR。
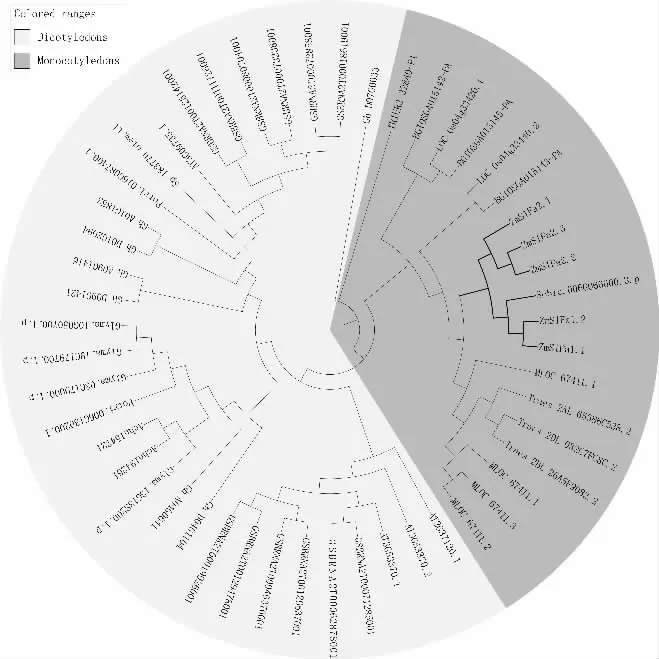
图3 S1Fa-like 基因家族系统进化树
2.3 ZmS1Fas 顺式作用元件分析
检索ZmS1Fas 启动子上已知的激素和胁迫相关的响应元件,结果显示(图4),ZmS1Fas 启动子区域含有丰富的激素(脱落酸、水杨酸、乙烯、茉莉酸、茉莉酸甲酯、生长素、赤霉素等)和胁迫(如高温、低温、干旱、厌氧、损伤等)相关的响应元件。尤其脱落酸响应元件,ZmS1Fas 成员平均含有13.2 个;其次是茉莉酸和茉莉酸甲酯响应元件,平均含有8.6 个。干旱响应元件也很丰富,在ZmS1Fa1.1、ZmS1Fa1.2和ZmS1Fa2.3 的启动子区均含有10 个以上。
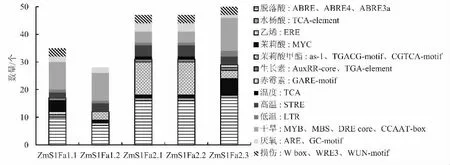
图4 ZmS1Fas 启动子区域激素和胁迫相关顺式作用元件的分布
2.4 ZmS1Fas 基因组织表达和胁迫表达分析
由图5 可知,正常条件下,除B73 成熟花粉(B73 Mature Pollen),各组织均有ZmS1Fas 基因表达,且多数组织中ZmS1Fa2 的表达量均比ZmS1Fa1的表达量高。授粉12 d 的胚乳(Endosperm 12 DAP)中ZmS1Fa 基因的表达量均高于其他组织部位中的表达量,B73 成熟花粉中表达量均低于其他部位的表达量。ZmS1Fa2 在根皮层、授粉27 d 的果皮、6~7节间、花丝等部位表达量也较高。

图5 正常条件下ZmS1Fas 表达模式分析
由组织表达模式可知,在根部位ZmS1Fas 表达量较高,选取根部位干旱胁迫表达数据,结果显示(图6),受干旱胁迫,其ZmS1Fas 转录水平并未达到显著上调或下调,推测ZmS1Fas 转录因子家族可能在蛋白质水平响应干旱胁迫。
图6 受干旱胁迫ZmS1Fas 在根部位的表达
由图7 可知,自交系Oh43 和杂交种Oh43×B73受高温胁迫,ZmS1Fas 均显著上调表达;自交系PH207 受低温胁迫,ZmS1Fas 均显著下调表达。不同玉米材料中,ZmS1Fas 响应温度胁迫模式不同,为选育耐高温或低温的玉米种质提供了理论支持,为玉米遗传改良提供潜在的基因资源。
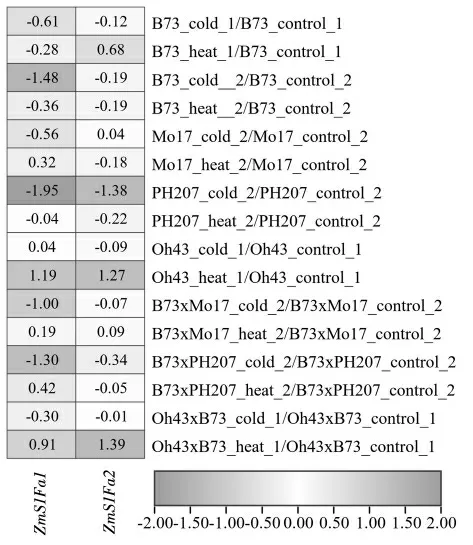
图7 不同玉米材料受温度胁迫ZmS1Fas 的表达
2.5 ZmS1Fas 热调控表达模式分析
为进一步明确ZmS1Fas 基因热胁迫表达模式,以ZmActin1 为内参,检测自交系B73 和Oh43 热胁迫表达量。设置正常条件下基因的相对表达量(CK)为1,结果(图8)表明,ZmS1Fa1 和ZmS1Fa2 在B73中均受热胁迫轻微下调表达;而在Oh43 中受热胁迫显著上调表达,受热胁迫处理材料中ZmS1Fa1 和ZmS1Fa2 表达量约是正常生长材料表达量的2 倍。与生物信息学预测结果基本一致,不同材料中ZmS1Fas 基因响应热胁迫模式不一致。笔者推测ZmS1Fas 在玉米响应非生物胁迫方面具有潜在的功能。
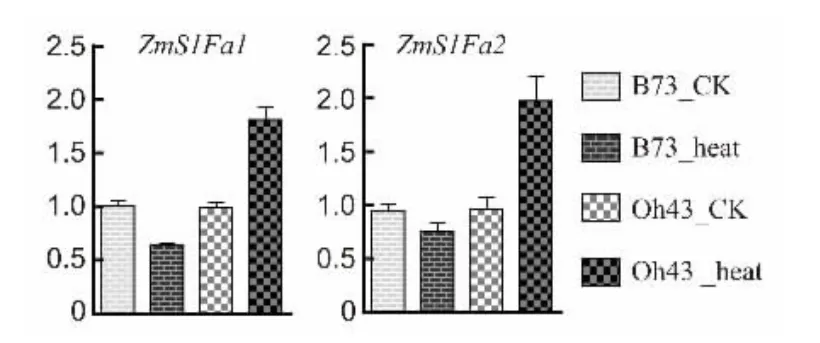
图8 热胁迫条件下ZmS1Fas 的相对表达量
3 结论与讨论
目前,玉米中许多逆境胁迫相关基因家族已经被鉴定和分析,如VOZ、GATA等基因,而关于S1Fa 基因的研究少之又少。本研究在玉米中共鉴定到2 个ZmS1Fas 基因,均含有S1FA 或S1FA superfamily 保守结构域,共编码5 个蛋白,长度为56~80 个氨基酸;分子量在6 310.77~8 867.10 Da 之间;均为碱性蛋白;不稳定系数均小于40,属于稳定蛋白,具有良好的热稳定性;α-螺旋、延长链、β-转角、无规则卷曲等蛋白质二级结构占比差别不大。ZmS1Fas 蛋白均定位在细胞核内。将大麦、玉米、籼稻、粳稻、高粱、小麦、乌拉尔小麦、猕猴桃、拟南芥、油菜、大豆、棉花、杨树、菠菜等14 种作物的51 个S1Fa-like 蛋白氨基酸序列进行多重比对,发现单子叶植物和双子叶植物的S1Fa-like 蛋白具有高度保守性。ZmS1Fas 保守序列较简单,motif1 是ZmS1Fas基因家族成员的核心功能区域。ZmS1Fas 均含有1~2 段外显子,0~1 段内含子,且均具有5'UTR 和3'UTR。ZmS1Fas 启动子区域含有丰富的与激素响应和非生物胁迫相关的调控元件。笔者推测S1Fa 转录因子作用元件可能参与植物逆境胁迫的响应,有利于植株的生长发育。
在正常生长条件下,除B73 成熟花粉(B73 Mature Pollen),各组织均有ZmS1Fas 基因表达,且多数组织中ZmS1Fa2 的表达量均比ZmS1Fa1 的表达量高。受到干旱胁迫时,ZmS1Fas 在转录水平与正常生长材料中表达量差异不显著。自交系Oh43 和杂交种Oh43×B73 受高温胁迫,ZmS1Fas 均显著上调表达;自交系PH207 受低温胁迫,ZmS1Fas 均显著下调表达。qRT-PCR 检测了热胁迫时ZmS1Fas 基因的表达量,结果表明,ZmS1Fa1 和ZmS1Fa2 在B73中均受热胁迫轻微下调表达;而在Oh43 中受热胁迫显著上调表达,与生物信息学预测的“ 不同玉米材料中ZmS1Fas 响应温度胁迫模式不同” 结果一致。这表明S1Fa 基因在复制后有可能发生功能分化,从而导致S1Fa 基因的功能多样性,促进了植物的生长调控和对自然环境的适应,为进一步解析ZmS1Fas在响应非生物胁迫应答中的生物学功能奠定基础。
