漓江大型底栖动物空间分布及水力驱动因子
2022-08-30李若男
刘 睿,李若男
(1.中国科学院生态环境研究中心 城市与区域生态国家重点实验室,北京 100085; 2.中国科学院大学, 北京 100049)
1 研究背景
人类活动改变了河流的水文情势,影响了河流底质、水动力条件及水体营养物质分布,从而改变了河流底栖动物的种群结构及空间格局。底栖生物是河流健康评价的重要指示指标,分析其群落结构的时空变化及水力驱动因子能够为研究河流生态修复和维持河流生态系统生物多样性提供有效的支持。
对于生态环境较好的山区河流来说,水质因子的空间梯度并不明显,水力梯度及地貌的变化使得水动力及底质因子成为影响底栖生物分布的主要影响因素[1]。近年来,Schffer等[2]研究发现底部剪切力以及底质粒径会影响底栖动物的分布。Burgazzi等[3]研究表明枯水期水动力因子决定了底栖动物的群落结构以及生物多样性。Schulz等[4]通过对热带和亚热带河流进行研究发现近床的紊动强度对底栖动物的空间分布有很大的影响。Blackwell[5]研究发现近床紊动强度会影响底栖动物触感器官的反应器,从而影响底栖动物的群落结构和分布。陈含墨等[6]对世界范围内不同气候带的7条大型河流进行研究发现,摇蚊、寡毛纲及蜉蝣目受水动力因子影响较大。方红卫等[7]的研究则表明,山区卵砾石河床的近床水流结构、水沙界面传质机理及潜流交换会产生较大的变化,从而影响底栖动物群落的空间分布。
本研究以广西壮族自治区漓江流域中下游典型河段为研究区域。已有针对漓江流域底栖动物分类、摄食类群以及其对水质因子响应关系的相关研究:曹艳霞等[8]在2008年2—3月份对漓江干流及支流底栖动物进行调查,结果表明昆虫纲是主要的物种类型,且干流与支流存在差异;杨青瑞等[9]研究了漓江全流域底栖动物与水环境之间的关系,认为温度、电导率、盐度、浊度、水深、总磷、化学需氧量为影响大型底栖无脊椎动物群落结构和分布的主要环境因子;王硕等[10]研究了漓江底栖动物的时空分布,研究表明漓江枯水期主要河段的物种多样性高于丰水期;朱晨曦等[11]对漓江底栖动物摄食类型进行研究,发现刮食者占绝对优势。目前对漓江底栖动物与水动力因子关系的研究仍较少。因此,本研究在分析漓江中下游大型底栖生物典型在不同水文时期的群落结构及空间格局的基础上,厘清其对水力要素的响应关系,旨在揭示河段尺度下水力要素对大型底栖动物群落结构及分布格局的影响机制,为变化水力条件下漓江水生态修复提供支持。
2 研究区域与方法
2.1 研究区域概况
漓江流域属珠江干流西江水系,位于广西壮族自治区东北部,是桂北地区的主要河流。漓江发源于兴安县华江乡猫儿山,全长214 km,流域面积12 285 km2,沿途流经兴安县、灵川县和桂林市区,最终于平乐县汇入桂江。本研究于2012年1月(枯水期)和 2012年 8月(丰水期),在漓江中游(长约10 km)及下游河段(长约10 km)各设置了27个采样点,采样点相对位置、采样次数及生境特征见图1、表1。

图1 研究区域与采样点Fig.1 Lijiang River basin and location of sampling sites
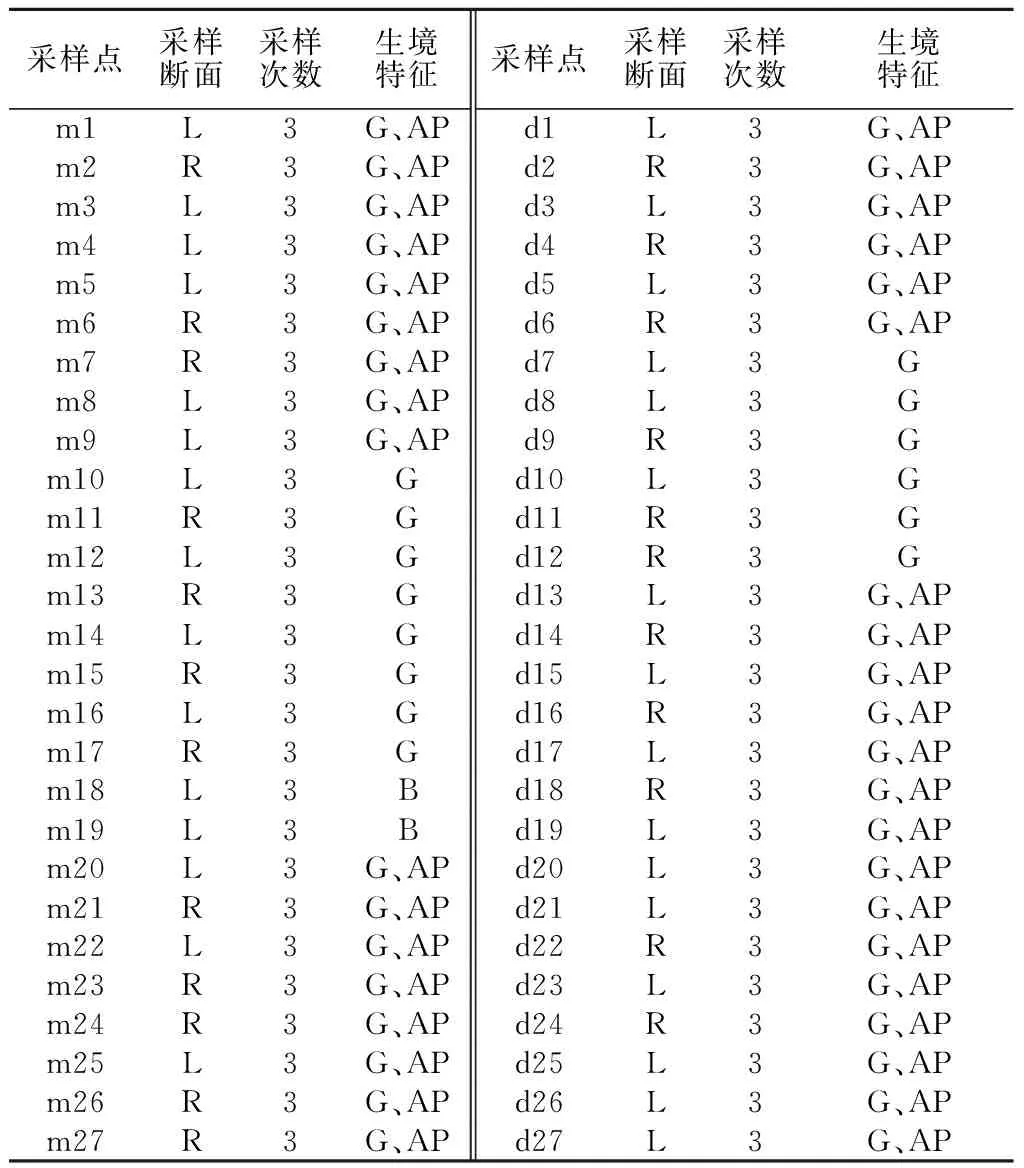
表1 采样点概况
2.2 研究方法
2.2.1 底栖动物数据采集及分析
当水深≤0.5 m 时,采用Surber 网(面积为0.09 m2,40 目(孔径0.425 mm))取样;当水深>0.5 m 时,采用 Peterson 改良式采泥器(面积为0.05 m2)采样。用孔径为 500 μm[12]的筛子现场筛洗样品,筛选出大型底栖动物,置于 100 mL标本瓶中,用5%的甲醛溶液固定生物样品,随后在实验室进行底栖动物种类的鉴定。物种的观察与鉴定采用体式镜(LEICA MZ95),对物种的形态特征进行观察识别,并进行记数。软体动物鉴定到种,摇蚊鉴定到科,其余物种通常鉴定到属级。功能摄食类群的划分根据室内底栖动物鉴定到种和属的结果,参照美国环境保护署(Environmental Protection Agency,EPA)建立的底栖动物摄食类群鉴别方法对已经鉴定出的漓江底栖动物进行分类,主要包括滤食者(Filter-collectors,FC)、刮食者(Scrapers,SC)、捕食者(Predators,PR)和收集者(Gather-collectors,GC)[13]。
采用Shannon-Wiener多样性指数(H)、Simpson指数(λ)、Margalef物种丰富度指数(D)、Pielou均匀度指数(J)来评估底栖生物物种多样性[10]。计算公式分别为:
H=-∑nilog2(ni/N) ;
(1)
(2)
D=(s-1)/lnN;
(3)
J=H/log2s。
(4)
式中:ni为第i种个体数;N为所有物种总个体数;s为总种类数;Pi为第i种个体数占群落总个体数的比例。
2.2.2 底质及水动力因子测量
漓江中游和下游以卵石和砾石为主,每个采样点分3次进行底质的采集,并绘制颗粒级配曲线。底质粒径采用底质质量分数倒推的粒径大小的平均数来计算平均粒径,D84、D50、D16分别表示底质质量分数为84%、50%、16%时对应的粒径。流速采用转子式流速仪(LS300-A)测定,水深采用塔尺测量。
2.2.3 底质及水动力因子选择
选择底质因子中的平均底质粒径、分选系数、河床糙率,水动力因子中的流速、水深、弗劳德数、雷诺数、边界雷诺数、剪切流速、底部剪切力、层流层厚度作为初始的底质和水动力因子[14-17]。为了减少冗余以及多重共线性(相关系数r>0.8)[18-19],对底质因子和水动力因子进行Pearson相关性分析,最后确定底质粒径、分选系数、流速、水深、底部剪切力和层流层厚度作为动因子。其中,底质粒径及分选系数为底质因子;其余动因子为水动力因子。测量及计算方法见表2。

表2 底质因子、水动力因子及计算方法
3 研究结果
3.1 底栖生物群落构成
漓江底栖动物群落结构组成如图2所示,漓江中、下游底栖动物种类名录如表3所示。枯水期共采集到35个分类单元,2 139个底栖动物个体,隶属于3门6纲27科。从分类单元看,枯水期腹足纲与昆虫纲占比最高,分别为40.00%和37.16%,其次为瓣鳃纲、寡毛纲、软甲纲和蛭纲(图2(a));对于丰水期,分类单元中腹足纲(37.50%)与昆虫纲占比最高(37.50%),其次为寡毛纲、瓣鳃纲、蛭纲和软甲纲(图2(b))。

图2 漓江底栖动物群落结构组成Fig.2 Structural composition of macrobenthos community in Lijiang River

表3 漓江中、下游底栖动物种类名录
由图2(c)、图2(d)及分析相关数据可知,枯水期腹足纲个体数占绝对优势(79.99%),优势种主要由中华圆田螺(Cipangopaludinacahayensis)和短沟蜷(Semisulcospirasp.)组成,底栖动物密度为264 ind/m2(每平方米动物个体数量为264)。丰水期共采集到40个分类单元,6 633个底栖动物个体,隶属于3门6纲30科;丰水期腹足纲个体数仍占据绝对优势(95.61%),优势物种为短沟蜷(Semisulcospirasp.)、中华圆田螺(Cipangopaludinacahayensis)及河蚬(Corbiculafluminea),底栖动物密度为819 ind/m2。
3.2 底栖动物摄食类群空间变化特征
从摄食类群相对丰度(图3)来看,刮食者(SC)在丰水期和枯水期的不同河段均占比最大,其中中游占比在85%以上,下游占比在70%以上;其次为滤食者(FC)、收集者(GC)和捕食者(PR)。从空间上看,中游刮食者(SC)的相对丰度大于下游,滤食者(FC)、收集者(GC)和捕食者(PR)的相对丰度均低于下游。从不同水文时期来看,丰水期的刮食者(SC)相对丰度高于枯水期,但其余摄食类群相对丰度均低于枯水期。

图3 漓江底栖动物功能摄食类群相对丰度占比Fig.3 Relative abundance proportions of functional feeding groups in Lijiang River during dry and wet seasons
3.3 底栖动物多样性空间变化特征
从研究河段看,漓江底栖生物多样性指数存在明显的空间异质性,中游普遍低于下游(表4),下游弯曲河段多样性高于上游顺直河段(图4),其中增幅最大的是丰水期Pielou均匀度指数(J),其次为Simpson指数(λ)和Shannon-Wiener指数(H);从不同水文时期来看,丰水期各类指数均呈现下游大于中游的趋势,变化幅度依次为Pielou均匀度指数(J)>Simpson指数(λ)>Shannon-Wiener指数(H)>Margalef物种丰富度指数(D);枯水期指数增幅趋势与丰水期相同,但枯水期中游底栖生物个体数相对物种数较少,使得中游Margalef物种丰富度指数(D)高于下游。

图4 漓江典型河段多样性指数空间格局Fig.4 Spatial pattern of diversity index at typical stream segments in Lijiang River
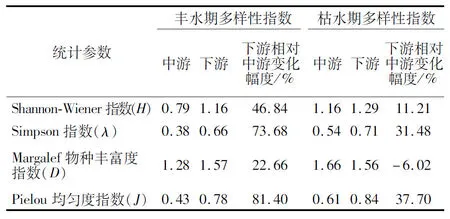
表4 典型水文时期多样性指数平均值
3.4 底栖动物与水动力因子响应关系
枯水期及丰水期底栖动物与水动力因子的典型关联分析(Canonical Correlation Analysis,CCA)如图5 所示。结果表明,漓江环节动物的分布与水深相关性最为密切,丰水期中环节动物与分选系数也有较高相关性,但枯水期中颤蚓目和吻蛭目的分布与层流层厚度相关性较高,颚蛭目则与底部剪切力大小有关。漓江软体动物丰、枯水期的主要影响因素相似,帘蛤目与基眼目与分选系数最为相关,中腹足目与贻贝目则与底质粒径、流速和剪切力相关。漓江节肢动物中的十足目、蜻蜓目和双翅目在枯水期和丰水期均受分选系数和层流层厚度的显著影响;而毛翅目及蜉蝣目在枯水期则主要受流速、底质粒径和剪切力的影响,丰水期则受分选系数、底部剪切力和流速的影响。
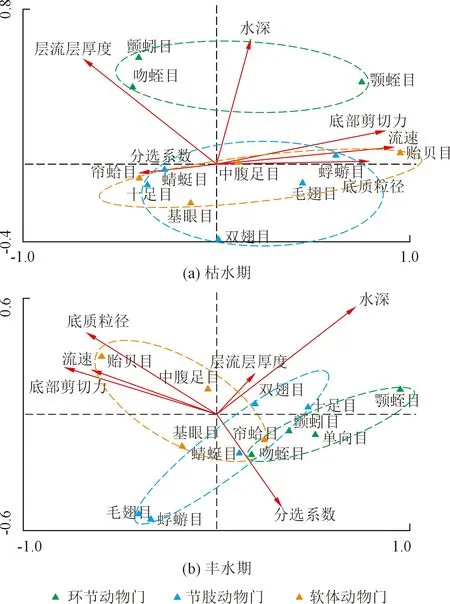
图5 底栖动物与水动力因子的CCAFig.5 CCA biplot of macroinvertebrates and hydrodynamic variables
图6为底栖生物多样性与水动力因子CCA。分选系数、层流层厚度和水深是漓江底栖动物多样性最主要的影响因素。其中底质分选系数是Shannon-Wiener指数(H)、Simpson(λ)指数和Pielou均匀度指数(J)最主要的影响因素,其次为层流层厚度;Margalef物种丰富度指数(D)在枯水期最主要的影响因素是水深,丰水期则是底质分选系数。

图6 底栖生物多样性与水动力因子CCAFig.6 CCA of macrobenthos biodiversity and hydrodynamic factors
4 成果讨论
4.1 底栖生物群落结构及摄食类群
研究河段内枯水期采集到的物种分类单元数与个体数均小于丰水期。这与在我国其他流域内的研究结果相似[20-21]。漓江丰水期水位上涨,使得原来裸露的卵石滩(卵石直径约20~115 mm)变成适宜腹足纲生活的浅滩生境,良好的生境使得腹足纲生物大量繁殖,从而在调查中获得了较多的腹足纲软体动物样本。
从摄食类群角度看,漓江中游和下游均以刮食者为主,其主要构成为腹足纲的软体动物。以腹足纲为主的刮食者体表有厣,足腺多分泌黏液有利于吸附在长有硅藻的卵石表面。陈凯等[22]的研究表明,漓江的捕食者一般以毛翅目为主,该类物种生物性状表现为身上有一层硬壳,不喜筑巢,在水中自由活动,喜欢生活在粒径较小的鹅卵石中,而漓江中下游底质粒径较大,不适宜该物种生活;漓江收集者一般以双翅目为主,该类物种不能直接摄取食物,多以微粒有机颗粒为食,喜欢在水生植物表面构筑巢穴,本研究中沉水植物覆盖稀疏,因此采集到的收集者较少。
从河段空间位置看,下游摄食类群与中游摄食类群相比刮食者减少,滤食者增加。这一方面是由于下游生境类型丰富,腹足纲等刮食者的生存空间被其他物种挤占,下游淤泥与细沙的比例高于中游,这种生境适宜颤蚓目滤食者生活,该类物种吸食淤泥或过滤有机碎屑;另一方面,王硕等[10]研究发现漓江下游水质略低于漓江中游,而颤蚓目为耐污种,更易出现在水质较差的区域。而下游流速小,更易于污染物的沉积。
4.2 底栖动物多样性时空格局
Beauger等[23]提出大型底栖动物可分为激流生物和缓流生物两类;Growns等[24]根据底栖动物的习性将底栖动物分为亲水种、厌水种及漂流种。通过对漓江底栖生物多样性、均匀度指数、丰富度指数空间分布的研究可以看出,底栖动物多样性与流速呈负相关,与底质粒径呈正相关。从研究河段典型水文季节来看,枯水期流速较缓,亲水种例如短沟蜷和厌水种例如卵萝卜螺均可以在漓江干流生活;丰水期流速较高,一些漂流种例如中华齿米虾随着水流漂流到缓流区,而厌水种例如卵萝卜螺则会消失,故枯水期的底栖动物多样性要高于丰水期。
此外,漓江浅滩底质以卵石为主,且通常以鱼鳞状排列,底质间孔隙度大,稳定性好,生境异质性高。这种河床结构可以降低河床扰动对底栖动物的伤害,可以为底栖动物提供庇护所。漓江深潭多分布在河道弯道凹岸一侧,水面比降减小,底质粒径较小,生境异质性不高,有适宜底栖动物附着的空间。Cobb等[25]发现洪水扰动会导致河床不稳定,水生昆虫的数量下降能达到94%。Brown[26]研究表明浅滩的河床结构可以附生更多的藻类,从而保持较高的底栖动物多样性。段学花等[27]研究表明粗颗粒底质中底栖动物多样性高于细颗粒底质。从研究河段空间特征来看,漓江丰水期河宽显著增加,河床的凹凸起伏对水面的比降影响降低,下游河道明显变宽,流速减缓,水生植物增多,生境类型更加多样化,使得底栖生物多样性增加。
4.3 环境因子对底栖动物分布的影响
通过对漓江中游和下游的枯水期和丰水期整体CCA可知,绝大多数物种的分布与分选系数以及层流层厚度密切相关,偏好分选系数适中、底质粒径均匀、层流层厚度较大的微生境。Growns等[24]通过研究发现,均匀的底质环境有利于有机质的沉积以及藻类的生长,适合刮食者的生存。本研究结果中,与分选系数以及层流层厚度相关的物种,如短沟蜷、赤豆螺、中华圆田螺等均为刮食者。
CCA结果还表明,枯水期时受水深影响的物种仅为部分环节动物,而丰水期所有环节动物、大部分节肢动物和软体动物的分布均与水深密切相关,这与Prezant等[28]的结果相似。Dudgeon[29]研究发现,部分十足目(米虾)通过漂浮来获取食物以及躲避天敌的摄食,枯水期水浅不利于漂浮,在这种情况下米虾需要一个稳定的底质环境来获取食物以及庇护,因此在枯水期米虾更多地受分选系数的影响,在丰水期则受水深与分选系数共同作用。此外,中国圆田螺、宽体金线蛭等软体动物门物种受流速和底部剪切力影响较大。
5 结 论
(1)漓江流域底栖生物分类单元中腹足纲与昆虫纲占主导地位,个体数上腹足纲占绝对优势;漓江典型水文时期底栖动物种类差别不大,但丰水期底栖生物密度为枯水期的3.1倍。
(2)从摄食类群看,刮食者在漓江典型河段均占主导地位,中游相对丰度高于下游;其次为滤食者,中游相对丰度小于下游。
(3)从多样性指数看,同一河段的河湾处各类指数低于平直河段,下游河段各类指数总体高于中游,丰水期不同河段的差异高于枯水期。
(4)从物种分布的主要影响因素看,底栖动物在枯水期间主要受流速、底部剪切力及底质粒径变化的影响,丰水期则主要受水深和层流层厚度的影响。
(5)从多样性指数的主要影响因素看,分选系数是多样性指数、均匀度指数和丰水期丰富度指数的主要影响因子,水深则影响枯水期物种丰富度的变化。
