拟南芥突变体myc234的鉴定及其对MeJA的敏感性分析
2022-08-16王炎炎张斐斐董萍萍殷伊君张有刚周丽娟
王炎炎,张斐斐,熊 娟,王 雪,李 林,冯 蕊,董萍萍,殷伊君,张有刚,周丽娟*
(1.云南省农业科学院 国际农业研究所,云南 昆明 650205;2.云南省农业科学院生物技术与种质资源研究所,云南 昆明 650205;3.昆明学院 农学与生命科学学院,云南 昆明 650214;4.云南师范大学 能源与环境科学学院,云南 昆明 650504)
茉莉酸(Jasmonates,JA)及其衍生物是植物体内重要的信号分子,其具有调节多个发育过程及抗逆防御反应的功能[1-2].在茉莉酸参与的众多生物学信号传导过程中,所依赖的大多为其下游的bHLH转录因子MYCs.不同的JA的信号转导过程中主要以MYC家族成员的MYC2为主,同时还有其同系物MYC3和MYC4发挥同功能冗余作用[3-6].在拟南芥(Arabidopsisthaliana)中,茉莉酸参与诱导芥子油苷合成所介导的防御反应.其中MYC2,MYC3和MYC4能够与MYBs互作促进下游基因的转录,也能够结合到下游芥子油苷相关合成酶启动子激活转录因子转录[5-7].在茉莉酸调控叶片衰老的发育过程中,MYC2/3/4结合到PAO的启动子上,促进叶绿素降解关键酶的表达[8],同时MYC2,MYC3,MYC4及MYC5还可以协同调控病虫害防御、叶片衰老以及侧根的发育[9].而JA响应反应中的几个MYCs转录因子在功能上存在冗余,单个基因的突变体表型往往不明显,只有获得纯合多突变体才能更好地对其基因功能丧失后的表型进行观察,从而确定其基因功能.因此,为研究JA及其下游转录因子的功能,制备myc234三突变体显得至关重要.基于此,本研究拟通过单突变体的杂交、自交获得初步确定的myc234纯合三突变体的F3代种子,结合PCR技术、SqRT-PCR技术以及JA敏感性试验综合验证纯合三突变体,旨在为JA相关研究提供材料,同时也可为相关突变体的鉴定提供具体的方法和数据.
1 材料与方法
1.1 试验材料
野生型拟南芥(Arabidopsisthaliana)为Columbia(Col-0)生态型,myc2为点突变,myc3(GK44B11),myc4(GK491E10)为T-DNA插入[10],上述材料由云南大学陈小兰教授惠赠.3个单突变体通过杂交、自交后对其F2代进行初筛,并将初筛成功的植株进行单株收种,获得稳定遗传的F3代.在茉莉酸羧基甲基转移酶催化下形成茉莉酸甲酯(Methyl jasmonate,MeJA),MeJA是植物体内主要的JA活性物质[4].本研究对野生型拟南芥及三突变体外源施加MeJA ,分析突变体对MeJA的敏感性变化.
1.2 材料培养
种子用V(30%过氧化氢)∶V(75%乙醇)=4∶ 1混合液消毒30 s,无菌水洗3次,分别点播于浓度为20,40 μmol/L MeJA的1/2MS固体培养基上,以不含MeJA的1/2MS培养基为对照,parafilm膜封口.倒置于4 ℃冰箱春化48 h,移至光照培养箱中进行垂直培养.培养条件为光照16 h/d,22 ℃,相对湿度60%~70%,在培养的第21天观察根的生长情况.
1.3 突变体的鉴定
植物总DNA的提取采用CTAB法,并将其保存于-20 ℃环境中.
引物设计:myc2为点突变[10],在其突变位点上下游设计引物,对PCR扩增片段进行测序鉴定.myc3和myc4的突变信息于SIGnAL数据库查询(http://signal.salk.edu/cgi-bin/tdnaexpress/),结合“双引物法”进行鉴定[11].利用primer 5进行引物设计(表1),并由生工生物工程(上海)有限公司合成.
PCR扩增体系(20 μL):GreenTaqMix(南京诺唯赞),10.0 μL,上下游引物各0.5 μL,ddH2O 8.0 μL,模板DNA 1.0 μL.扩增条件:预变性(94 ℃,5 min),变性(94 ℃,30 s),退火(60 ℃,30 s),延伸(72 ℃,45 s),33个循环,延伸(72 ℃,5 min).
1.4 SqRT-PCR
采用TRIzol法提取总RNA,按照MonScriptTMRTIIIAll-in-OneMixMonad(购于莫纳生物)说明书的方法建立20 μL标准反转录体系,并去除基因组DNA及合成cDNA.利用primer 5进行引物设计(表1),并由生工生物工程(上海)有限公司合成.SqRT-PCR以ACTIN基因为内参,分别在野生型col-0和myc234中扩增ACTIN,MYC2,MYC3和MYC4.PCR扩增体系(20 μL):GreenTaqMix(南京诺唯赞),10.0 μL,上下游引物各0.5 μL,ddH2O 8.0 μL,模板DNA 1.0 μL.扩增条件:预变性(94 ℃,5 min),变性(94 ℃,30 s),退火(60 ℃,30 s),延伸(72 ℃,30 s),28个循环,延伸(72 ℃,5 min).

表1 引物序列
1.5 数据统计与分析
MeJA敏感性试验中,用游标卡尺准确测量培养了21 d拟南芥各处理的根长,并利用SPSS 21.0软件对测量数据进行方差分析.
2 结果与分析
2.1 myc2纯合突变体的鉴定
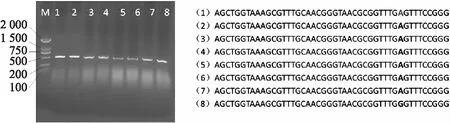
myc2突变位点是位于582 bp的G颠换为A.通过PCR扩增,扩增产物经1.0%琼脂糖凝胶电泳检测,显示条带清晰,且与目标条带一致,结果详见图1(a).通过测序及序列比对可知7株突变体材料在突变位点均为碱基A的单峰,而Col-0野生型对照(8号样)为碱基G的单峰(结果见图1(b)),说明所鉴定的7株突变体样品均为myc2的纯合突变体.
2.2 myc3纯合突变体的鉴定
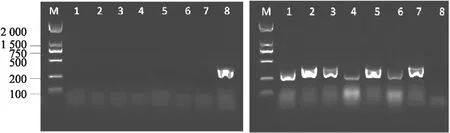
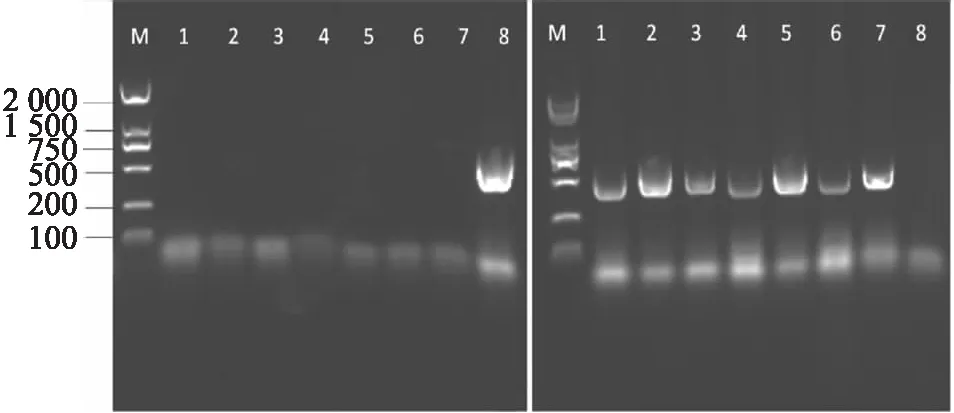
myc3(GK445B11)为拟南芥MYC3(AT5G46760)基因的T-DNA插入突体,其插入位置为 1 587 bp处外显子[10].通过双引物法,分别以LP+RP和RP+LB为引物进行两组PCR扩增.PCR扩增产物经1%琼脂糖凝胶电泳结果分析可知,以LP+RP为引物时仅有Col-0野生型有目标条带,而7株突变体均无目标条带(图2(a));在RP+LB为引物进行PCR扩增时仅Col-0无目标条带,而7株突变体均有目标条带(图2(b)),综合两组试验结果说明1~7号植株均为myc3纯合突变体.
(a)PCR产物电泳图 (b)PCR产物测序结果
(a)MYC3全长 PCR检测 (b)突变体T-DNA插入位点PCR检测
2.3 myc4纯合突变体的鉴定
myc4(GK491E10)为拟南芥MYC4(AT4G17880)基因的正向T-DNA插入突体,其插入位置为外显子 271 bp 处[10].试验分别以LP+RP和LP+LB为引物进行两组PCR扩增并进行琼脂糖凝胶电泳,以LP+RP为引物的扩增结果显示仅有Col-0野生型有目标条带,7株突变体均无目标条带(图3(a));在LP+LB为引物进行PCR扩增的试验结果中显示7株突变体均有目标条带,而Col-0无目标条带(图3(b)).因此,7株突变体均为myc4的纯合突变体.
(a)MYC4全长PCR检测 (b)突变体T-DNA插入位点PCR检测
2.4 突变体基因表达量分析
将qRT-PCR产物通过1.5%琼脂糖凝胶电泳的结果显示(图4),内参ACTIN在突变体myc234和野生型Col-0的植株中均能正常扩增出清晰条带,而MYC2,MYC3,MYC4这3个基因在myc234中表达量较野生型对照明显下降,进一步证实PCR鉴定出的纯合三突变体植株中MYC2,MYC3和MYC4基因在转录水平上几乎无表达.
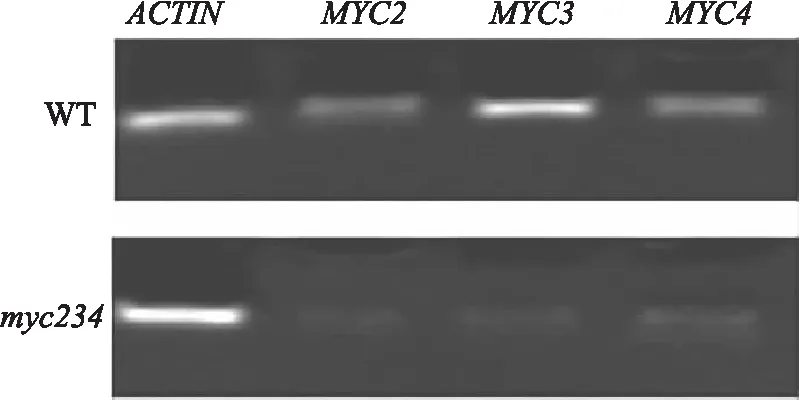
图4 MYCs基因SqRT-PCR扩增产物
2.5 突变体对茉莉酸的敏感性试验
试验将消毒后的野生型Col-0及纯合三突变体种子点种于含有40 μmol/L MeJA的1/2MS培养基上,并设不含MeJA的1/2MS的培养基为对照,通过垂直培养观察幼苗根的发育的情况(图5).试验结果表明,培养21 d后,MeJA对野生型幼苗根的发育造成了严重的抑制作用,而myc234突变体对MeJA的抑制效应的敏感性较对照明显下降.野生型植株在40 μmol/L处理试验中的平均根长比空白组下降了39.57%,差异有统计学意义;而myc234纯合突变体根长反而上升3.54%,差异无统计学意义.上述结果表明,myc234突变体的根发育对JA的调控作用不敏感.
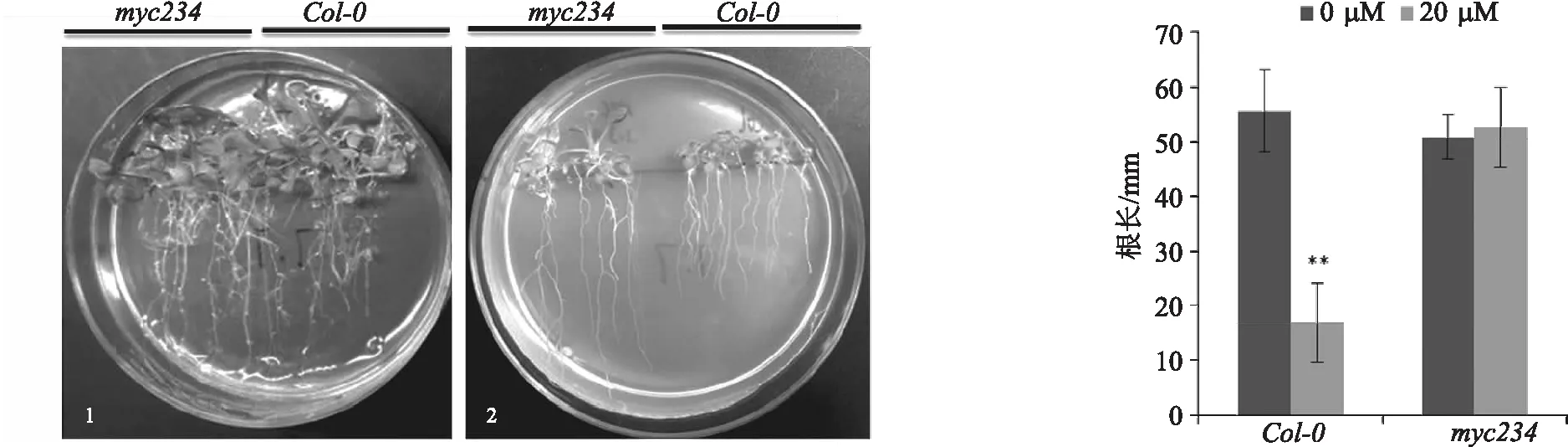
(a)拟南芥野生型和突变体根生长对MeJA敏感性表型 (b)拟南芥野生型和突变体根生长对MeJA敏感性统计分析
3 讨论与结论
3.1 讨论
在茉莉酸调控的根发育、气孔发育、雄蕊发育及芥子油苷合成的生物学过程中[12-15],MYC2,MYC3,MYC4均以功能冗余的方式参与其中,但在某些发育过程中三者也具有各自独立的特异性功能,如MYC2在整个植物中都有表达,特别是在根组织中表达强烈,MYC3主要在营养组织中表达,MYC4在维管组织中表达[16].就目前的研究报道来看,MYCs的三个成员的基因功能还有很多不清楚的地方.同时,对于myc234三突变体的鉴定方法也存在很多差异,因此鉴定获得myc234纯合三突变体材料,建立其鉴定方法体系,对于研究JA相关调控机制,深入了解MYC2,MYC3,MYC4的具体生物学功能具有重要的意义.
本研究利用40 μmol/L MeJA对拟南芥进行外源化学处理以确定突变体对JA的敏感度.在野生型中,MeJA处理后其根长较未处理对照下降了39.57%,两者差异有统计学意义;而在myc234纯合三突变体中,处理样较对照样根长未受抑制,反而增加了3.54%,说明该myc234纯合三突变体对JA的敏感性明显下降.这一结果与Chen等[12]的研究结论的总体趋势一致.但本研究发现,在对JA敏感的度的测试试验中与多篇文章所用的MeJA浓度有所区别,Chen等[12]用的是20 μmol/L,乔菊香等[14]用的是28 μmol/L,Zhuo等[15]用的是 5 μmol/L,而且在处理的时间上也各不相同,由此可以看出,在对拟南芥进行外源化学处理的敏感试验中,虽然各个试验的总体趋势一致,但未形成统一的处理标准,在数据的具体细节方面存在一定的差异.因此,在后续试验中应考虑浓度梯度的问题,加强试验的全面性及系统性.
3.2 结论
通过PCR鉴定,SqRT-PCR半定量试验以及MeJA敏感试验,证明了通过杂交获得的突变体材料为myc234的纯合三突变体.该突变体材料中MYC2,MYC3和MYC4在转录水平基本无表达,其根发育对MeJA的敏感程度明显低于野生型,结果可为深入研究MYC2,MYC3和MYC4参与JA以及其他激素调控生理生化过程中发挥的功能提供研究材料.
