草地退化对三江源国家公园高寒草甸表层土壤有机碳、全氮、全磷的空间驱动
2022-08-15张法伟李红琴仪律北罗方林张光茹王春雨杨永胜李英年
张法伟,李红琴,仪律北,罗方林,张光茹,王春雨,杨永胜,李英年
1 中国科学院三江源国家公园研究院, 西宁 810001 2 洛阳师范学院生命科学学院, 洛阳 471934 3 中国科学院西北高原生物研究所高原生物适应与进化重点实验室, 西宁 810001 4 青海省林业和草原局林业碳汇服务中心, 西宁 810008
青藏高原是中国重要的生态安全屏障区,高寒草甸面积约为58.2万km2,是高原的主要植被类型和生态屏障功能发挥的基质[1—2]。在气候变化和人类活动的双重压力下,草地退化态势依然严峻,严重危及高寒草甸碳素固持、水源涵养及气候调节等生态服务功能[3—5]。土壤退化是草地退化的核心问题,不仅影响了土壤的碳氮磷等养分的供需平衡及植被生长,而且会加速草地退化的演替进程[6—7],甚至改变区域气候变化的趋势[8—9]。深入研究草地退化对土壤营养元素的驱动机制是高寒草甸可持续发展的关键,也是退化高寒草甸近自然恢复[10]和青藏高原国家生态安全屏障建设的关键之一[2],具有重要的理论价值和现实意义。
土壤有机碳是草地碳循环及草地质量评价的核心指标[11—12],土壤氮、磷被认为是草地生态系统生产力的主要限制因素[9, 13—14]。草地退化通过改变高寒草甸植被地上、地下输入物的数量与性状及土壤水热微环境[7, 15],调控微生物的群落组成、生理性状及残体数量[12, 16],导致土壤有机碳、全氮含量下降[17—18],但降低程度与研究区的气候因素及土壤深度密切相关[1, 19—21]。但草地退化[22—24]和围栏封育[25]对土壤全磷含量均没有显著影响。草地退化显著改变了土壤而非植被的生态化学计量[21, 26],从而影响了土壤-牧草的营养供需平衡[6, 25]乃至系统稳定性及演替进程[24, 27]。目前相关的研究大多集中在单点尺度,但高寒草甸分布范围广,气候异质性强,限制了研究结果的空间代表性[28]。
三江源国家公园位于青藏高原的核心区,总面积为12.31万km2,是中国首批正式成立的国家公园之一,是高原生态安全屏障的重要地理单元[29]。高寒草甸约占园区面积的44%,是国家公园的主体植被类型,虽然经过近二十年的生态工程治理,但草地退化态势仍然严峻, 削弱了区域的土壤承载力及生态服务功能[1, 30]。本文基于2020年7—8月的三江源国家公园高寒草甸原生植被和退化植被的60个配对采样,研究草地退化对表层(0—30 cm)土壤有机碳(SOC)、全氮(TN)和全磷(TP)含量及其计量特征的空间驱动强度和潜在生态过程,为三江源国家公园高寒草甸土壤承载力的评估和退化草地的恢复治理提供数据支撑和理论基础。
1 材料与方法
1.1 配对采样与室内分析
本次采样集中在三江源国家公园高寒草甸的典型分布区(图1)。原生植被中,以小嵩草(Kobresiapygmaea)、矮嵩草(K.humilis)、早熟禾(Poapratensis)和麻花艽(Gentianastraminea)等为群落优势种,群落相对盖度为85%以上;退化样地内,优势种包括细叶亚菊(Ajaniatenuifolia)、鹅绒委陵菜(Potentillaanserina)、甘肃马先蒿(Pediculariskansuensis)、黄花棘豆(Oxytropisochrocephala)、兰石草(Lanceatibetica)等,群落盖度一般小于40%,为重度退化的稳态[7]。在研究区域内,高寒草甸的原生植被和退化植被通常会相邻出现,且少有退化过程的中间稳态(图1)。因此,本研究采用配对采样的方法,以降低气候、土壤的空间异质性对研究结果的影响。在2020年7—8月,兼顾采样难度和代表性,沿公路两侧1 km外共采集了20组60个配对样地(长江源园区、黄河源园区和澜沧江源园区分别采集了10、7、3组),基本覆盖了三江源国家公园高寒草甸的主要分布区。

图1 三江源国家公园高寒草甸采样点及原生植被和退化植被的配对样地的典型景观
在原生植被和退化植被的每组配对样地内,半随机选取三个样方(以期符合原生植被和退化植被的群落表观特征),按照0—10、10—20、20—30 cm的土壤深度,用内径7.0 cm的土钻采集土壤样品,过2.0 mm筛子后装入土袋,带回室内阴干,制成0.25 mm的土壤粉末,进行SOC、TN和TP的含量(%,g/100g)测定。其中SOC用重铬酸钾硫酸外加热法, TN用凯氏定氮法,TP采用硫酸-高氯酸-钼锑抗比色法测定。以SOC/TN,SOC/TP和TN/TP表示C/N、C/P和N/P等土壤化学计量特征。
1.2 统计方法
利用配对样本t检验(Paired-samplest-test)分析草地退化对SOC、TN和TP的影响强度。以原生植被与退化植被的SOC、TN、TP、C/N、C/P、N/P及二者差值(ΔSOC、ΔTN、ΔTP、ΔC/N、ΔC/P和ΔN/P)为因变量,以采样点的经度、纬度、海拔、草地退化(分类变量)和土壤深度(分类变量)为自变量,利用一般线性模型探讨草地退化对高寒草甸土壤营养性状对空间驱动强度的环境调控机制。统计分析和绘图分别在R 4.0.2[31]和OriginLab 2016中完成。
2 结果与分析
2.1 土壤有机碳、全氮和全磷的响应及环境调控
配对样本t-检验表明,草地退化对SOC和TN的降低效应极显著(P<0.001),但对TP无显著影响(图2)。原生植被的SOC和TN分别为(2.45±2.05)%(平均值±标准差,下同)和(0.25±0.20)%,退化植被的SOC和TN较原生植被分别低44.0%和35.6%。原生植被和退化植被的TP分别为(0.039±0.015)%和(0.038±0.014)%,二者差异不显著(P=0.22)。原生植被与退化植被的0—10 cm和20—30 cm的SOC差异相对较大,分别为47.0%和45.0%,而中层(10—20 cm)的差异为40.3%。TN也表现出相似的趋势,即浅层和下层差异相对较大,分别为38.4%和38.3%,中层差异为31.9%。原生植被和退化植被的TP在表层、中层和深层的差异均小于6%,也表现出表层和下层略大(5%),而中层略小(1%)的特征。原生植被和退化植被的土壤容重平均分别为(0.93±0.34)g/cm3和(1.12±0.21)g/cm3(二者差异不显著,P=0.06),因此,表层SOC和TN的绝对损失量分别为2.23 kg/m2和0.16 kg/m2。

图2 三江源国家公园高寒草甸原生植被和退化植被的0—30 cm土壤有机碳(SOC)、全氮(TN)和全磷(TP)含量的配对检验
高寒草甸土壤营养元素与草地退化(分类变量)、土壤深度(分类变量)、经度、纬度和海拔的一般线性模型表明,SOC和TN主要受草地退化、土壤深度和纬度显著影响,而TP则受经度、纬度和土壤深度调控(表1)。但草地退化对ΔSOC的空间降低效应主要受纬度和海拔影响,和经度、土壤深度的关系较小(表1)。草地退化导致ΔSOC的降低幅度平均为(1.09±1.83)%,与纬度呈现出线性负相关特征(斜率=-0.98,R2=0.16,P<0.001),和海拔呈现出线性正相关特征(斜率=0.0034,R2=0.08,P<0.001)。草地退化导致ΔTN的降低幅度平均为(0.092±0.16)%,也表现出类似的环境调控特征,随纬度降低和海拔升高而升高。ΔTP对草地退化存在降低响应趋势,也无显著环境影响因素。因此,三江源国家公园中,低纬度高海拔区域的高寒草甸SOC和TN对草地退化响应更为强烈。
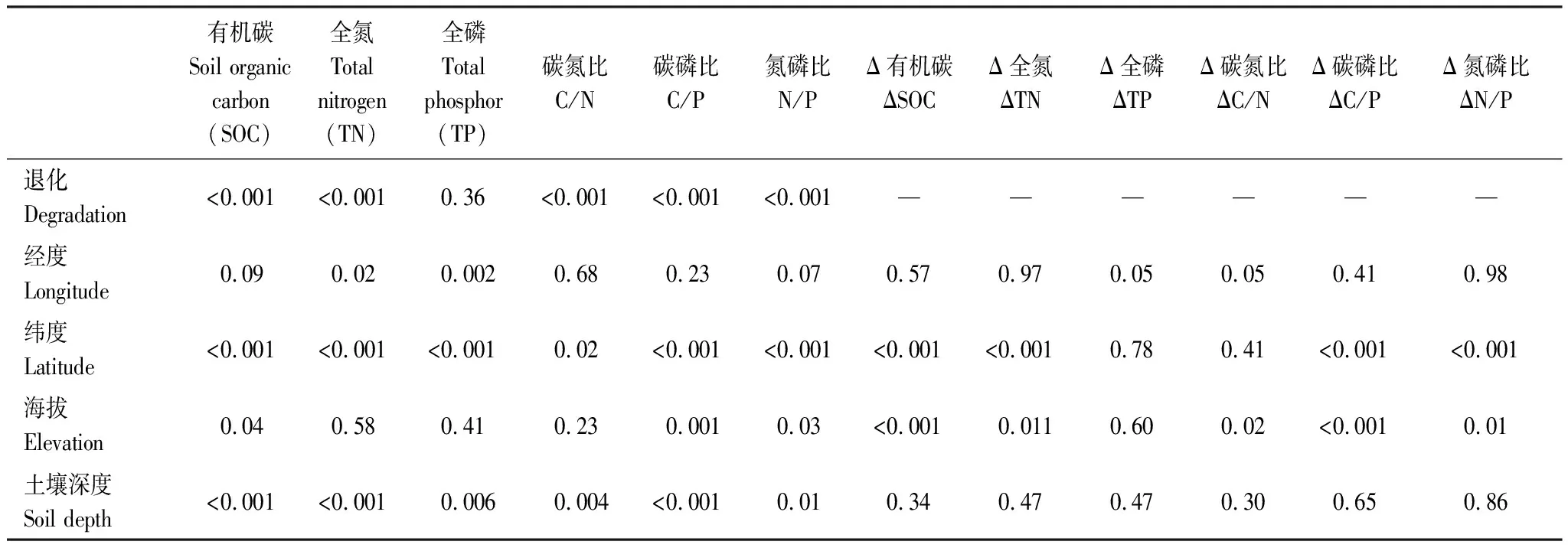
表1 三江源国家公园高寒草甸原生植被与退化植被的土壤营养元素及其差异与环境因子的一般线性模型的显著性检验
2.2 土壤生态化学计量特征的响应及环境调控
原生植被和退化植被的C∶N∶P平均分别为59.6∶6.2∶1.0和35.6∶4.2∶1.0,草地退化导致计量值的降低程度平均为28.3%。原生植被和退化植被的C/N分别为9.58±2.35和8.33±1.23。相关分析表明原生植被和退化植被中的C/N、C/P均主要受SOC影响(P<0.001),而与TN或TP的关系较小(P>0.07)。N/P也主要受TN(P<0.001)而非TP(P>0.06)的影响。C/N与草地退化(分类变量)、土壤深度(分类变量)、经度、纬度和海拔的一般线性模型表明, C/N主要受草地退化、土壤深度和纬度显著影响(表1)。草地退化和土壤深度没有显著交互作用(P=0.83)。C/N随草地退化和土壤深度显著下降。除了经度无显著影响外,草地退化、土壤深度、纬度和海拔均对C/P和N/P有显著作用(表1)。
原生植被的C/P和N/P分别为59.64±39.10和6.16±3.65,配对样本检验表明草地退化导致C/P和N/P分别显著降低了40.3%和31.4%。草地退化对ΔC/N的空间驱动强度主要取决于海拔(表1),即随着海拔高度增加,草地退化的效应越强。草地退化对ΔC/P和ΔN/P的降低效应主要受纬度和海拔共同影响,其与纬度负相关,和海拔正相关。因此,低纬度高海拔地区的高寒草甸土壤化学计量特征对草地退化的响应更为强烈。
3 讨论
3.1 土壤养分含量对草地退化的响应特征
草地退化导致高寒草甸损失约一半SOC和三分之一 TN,但对TP无显著影响,这与三江源高寒草甸相关研究的结果基本一致[16, 18]。因此,高寒草甸SOC和TN对草地退化的响应最为敏感,而全磷的敏感性较低[21, 24], 即SOC和TN是表征高寒草甸土壤退化的适宜养分指标[7, 18, 26]。SOC主要取决于植物及微生物残体输入与土壤微生物呼吸分解的平衡,而表层SOC的稳定性更多地受植物输入性状和水分状况的影响[8, 12]。草地退化减少了地上植被生物量,同时植被群落也由易矿化的禾草、莎草转变为难分解的杂类草[7],降低了凋落物和根系的数量及质量[1];而且土壤微生物残体做为高寒草甸表层SOC的重要来源[11—12],也随着草地退化而显著下降[16]。另外,退化草地地表近乎裸露,土壤侵蚀风险增大[32],而表层水热变化变幅增加也导致植被生长季的土壤呼吸作用增强[15],共同降低了表层SOC。在土壤可利用碳水平较低时,微生物处于休眠或不活跃状态,降低了微生物的矿化能力和土壤营养的供给水平,有利于草地退化稳态的维持[25]。
相对SOC,TN降低程度略低,可能是在退化草地中,较低的植被覆盖及生产力缓解了植被生长对土壤氮素的需求量[33],且可食牧草的降低也减少了因家畜采食而导致的氮素迁移损失[6],同时豆科(如黄花棘豆)等不可食牧草的增多也可一定程度提高土壤氮含量。因此,退化草地的土壤氮素含量虽然下降,但其供给却可能满足当前植被生长对氮素的需求[6],是高寒草甸退化稳态的重要维持因素之一[5]。草地退化对土壤TP的影响不显著,这也与前人的研究结果一致[22—24]。由于磷素受土壤母质的影响[33],主要来源于矿石的风化而并非有机质的分解[24]。同时,磷没有气体形式,而磷酸盐在高寒草甸碱性土壤环境中的可溶性较低,难以淋溶[25]。因此,三江源高寒草甸TP对草地退化没有显著响应,可能并非植物生长的主要限制因子[26]。
草地退化对ΔSOC和ΔTN的影响强度与纬度、海拔显著相关,但与经度和土壤深度的关系较小(表1)。这主要由于研究区域纬度、海拔与气温、降水有显著的空间相关性,而热量状况和水分含量是土壤呼吸[1]及有机碳、氮矿化[8, 34]的主要环境调控因子。而且纬度的负作用相对更强(表1),暗示高寒草甸所处地域的气温越高,ΔSOC和ΔTN对草地退化的响应越强。这可能由于高寒草甸多为温度限制系统[9],在温暖环境中,土壤微生物的维持呼吸也相对较高,导致作为SOC重要来源的微生物残体减少[11—12],加之植物根系生命周期相对缩短,不利于死亡根系对SOC的补充和保留,加速了SOC的周转[8—9]。草地退化导致ΔSOC的空间降低效应与土壤深度的关系微弱(表1),可能是随着土壤深度增加,透气性降低,可供降解的SOC及微生物数量迅速下降,且下层SOC的稳定性更多受到土壤矿质的影响,而并非植被有机碳的输入调控[8, 12]。
3.2 土壤生态化学计量对草地退化的响应
土壤生态化学计量是不同化学元素之间的稳态平衡,影响着生态系统的演替进程及对环境变化的响应方式,是植被群落健康和系统稳定性的重要指标[14, 26]。三江源国家公园高寒草甸表层土壤生态化学计量特征,处在前人对三江源高寒草甸的相关研究范围之内[26, 35],但显著低于全球土壤的平均水平(186∶13∶1)[36]。草地退化显著降低了土壤C/N、C/P和N/P,表明草地退化深刻改变了土壤营养元素的供给比例[24, 26]。但C/N的降低幅度较小(13.0%),这由于土壤氮素主要存在于土壤有机质中,二者变化相对一致[13]。草地退化降低了三江源高寒草甸土壤C/N,加之植被覆盖降低,暗示草地退化可能一定程度上缓解了植被生长的氮限制[6],有利于退化稳态的维持[5]。三江源国家公园高寒草甸土壤C/P远小于我国土壤C/P(105∶1),这主要由于青藏高原土壤发育年轻导致磷素的矿化速率相对较高但利用水平相对较低[24]。土壤N/P小于10,暗示高寒植物生长表现为N限制,这与青藏高原植被生产力和氮素密切耦合的结论一致[26, 35]。草地退化对土壤生态化学计量特征的空间降低效应主要取决于海拔和纬度,与草地退化对SOC和TN的空间影响因素基本一致,这主要因为C/N、C/P主要受控于SOC,而N/P则受控于TN。
4 结论
基于三江源国家公园高寒草甸原生植被和退化植被的配对研究,草地退化显著降低了表层土壤有机碳和全氮含量及土壤生态化学计量特征,但对土壤全磷含量的影响微弱。草地退化对土壤有机碳和全氮的空间降低效应主要受控于纬度和海拔,对低纬度高海拔高寒草甸的驱动更为强烈。在三江源国家公园中,草地退化导致高寒草甸土壤碳氮损失严重,改变了土壤营养元素的供需平衡,是退化稳态维持的重要因素之一。
