北冰洋中部阿尔法脊晚第四纪介形虫化石群与古海洋环境变迁
2022-08-15王雨楠周保春王汝建肖文申
王雨楠,周保春,王汝建,肖文申
1. 上海自然博物馆(上海科技馆分馆),上海 200041
2. 同济大学海洋地质国家重点实验室,上海 200092
北冰洋被高耸的洋中脊—罗蒙诺索夫脊分隔为东、西两部分。现代北冰洋的显著特点之一是海水的分层结构[1],它拥有以下四大主要水团:极地表层水(PSW,水深0~50 m,水温-2~0 °C,盐度32~34 psu),大西洋水(AW,水深200~1 000 m,水温≥0 °C,盐度34.3~34.8 psu),北冰洋中 层 水(AIW,水温-0.5~0 °C,盐度34.6~34.8 psu),以及北冰洋深层水(AODW);AODW 分为加拿大海盆深层水(CBDW,水深>1 500 m,水温-0.5~-0.3 °C,盐度34.95 psu)和欧亚海盆深层水(EBDW,水深>2 000 m,水温-1.0~-0.6 °C,盐度34.94 psu)[2-5]。AW和PSW被一层发育良好的盐跃层(水深<200 m)所分隔[6]。温度和盐度较高的北大西洋水体经由弗拉姆海峡和巴伦支海流入北冰洋,潜入相对低盐的盐跃层之下并逆时针流动[4,7],形成了位于中层的AW和AIW[8],因此北大西洋是北冰洋重要的热量和盐分供给源。在北冰洋,大尺度的表层洋流有从西伯利亚陆架流向弗拉姆海峡的穿极洋流,以及加拿大海盆的波弗特环流(图1a)。北冰洋从欧亚大陆和北美大陆接受了约占全球总量10%的淡水流量[9],而波弗特环流夹裹了大量的淡水[10-12]。阿尔法脊的表层水被常年海冰所覆盖[13],同时也处于波弗特环流的影响之下。
Haley等[14]对北冰洋岩芯中的钕同位素分析结果显示,北冰洋海水的分层格局形成于早更新世,但中层水体(包括AW和AIW)的性质随气候而变化:间冰期,中层水体主要来自北大西洋中层水的贡献,而冰期则主要来自于在欧亚大陆架上形成的卤水。另一方面,Cronin等[15]利用介形虫壳体Mg/Ca古温度计重建了50 ka以来北冰洋的水团变化历史,得出了末次冰期中盐跃层增厚,AW在整个北冰洋均向下被挤压到1 000~2 500 m深度的著名结论。然而,我们发现Cronin等的结论虽然可以很好地解释末次冰期中介形虫化石群在北冰洋罗蒙诺索夫脊以东的分布方式,却无法解释它们在罗蒙诺索夫脊以西的分布。
介形虫是北冰洋微体化石的重要门类之一。Gemery等[16]基于整个北冰洋1 000多个表层沉积样数据而建立的北冰洋介形虫数据库(Arctic Ostracoda Database, AOD)表明,介形虫属种与海洋环境之间存在着明确的对应关系。自20世纪80年代以来,介形虫在北冰洋新近纪—第四纪古海洋学中的应用越来越多,揭示了介形虫化石群与古气候事件的关联性[17]。在过去几年里,本文作者对中国北极科学考察期间“雪龙号”科考船在西北冰洋楚科奇海台一带钻取的3根岩芯(水深<1 000 m)中的介形虫及部分底栖有孔虫属种进行详细研究,并对前人的北冰洋介形虫化石数据[18-19]进行重新分析后,得出了如下结论:在包括末次冰期在内的 MIS 5–2期,AW在西北冰洋并非迁移到更深水域,而是因受罗蒙诺索夫脊的阻隔而变弱甚或消失了[17,20]。
我们的上述结论是对Cronin等[15]结论的重要修正,揭示了气候寒冷期里东、西北冰洋迥异的水体结构,它有待更多的研究来进一步证实。这同时也衍生出了新的科学问题:在更深水域中,北冰洋的水团是否也随气候冷暖交替而发生变化?作为解答这一问题的第一步,本研究选取了中国第三次北极科学考察期间在北冰洋中部阿尔法脊采取的岩心ARC3-B84A(站位水深2 280 m),对其中的介形虫化石群进行了详细分析。研究内容包含两个方面:首先,基于北冰洋介形虫数据库(AOD)[16],对ARC3-B84A岩芯中介形虫主要属种在现代北冰洋的生态分布,尤其与水团的对应关系进行了分析;然后,根据主要属种在该岩芯中的时代分布,重建了MIS 13期以来深部水团在阿尔法脊一带海域的变迁历史。
1 材料及方法
1.1 研究材料
本研究材料来自中国第三次北极科学考察[21]于2008年夏季在阿尔法脊采集的ARC3-B84A岩芯(以下简称B84A岩心)。该岩芯长188 cm,站位水深2 280 m,位于AODW上部;与之相比较,用于西北冰洋中层水团研究的3根岩芯(ARC3-P31、ARC6-R14及ARC7-P12)[17]均采自楚科奇海台,站位处于现代北冰洋AW之内(图1,表1)。
1.2 研究方法
本研究采用标准微体古生物学分析方法对B84A岩芯沉积物样品进行处理。岩芯按2 cm间隔取样,共获得94个样品。在同济大学海洋地质国家重点实验室,将分割的样品用烘箱烘干、称重,然后经水泡分散后,用孔径63 μm的铜筛水洗。将>63 μm的屑样烘干后,再用孔径150 μm的筛子干筛。在体视显微镜下,将>150 μm屑样中的介形虫个体全部挑出用于属种鉴定统计分析。本文主要采用Yasuhara等[25]、Gemery等[16]作为北冰洋介形虫分类鉴定标准。此外,利用同济大学海洋地质国家重点实验室的扫描电子显微镜(TESCAN Mira 3)对介形虫标本进行观察并拍摄照片。
本研究计算了每个样品中的介形虫丰度(每克干样中介形虫的壳瓣数),因为它反映了底栖生物古生产力变化;计算了常见属种在每个样品中的个体数量百分比,以便复原古水团变迁。介形虫个体数用单瓣数来表示,1个合瓣计为2个单瓣。在计算各属种的百分比时,为了减少误差,本文将个体数量较少的相邻样品合并起来,以使合并后的瓣数大于50。
为了准确把握介形虫属种的水深分布及其与水团的对应关系,本文使用软件Ocean Data View(ODV)[26]分析了B84A岩芯中的常见种在现代北冰洋的分布。共计选取了北冰洋638个表层沉积物样品的介形虫数据,并将数据输入ODV软件绘制各个属种在现代北冰洋的丰度等值线断面图。这些样品由AOD[16]的1 340个样品中的626个,以及我们自己拥有的231个西北冰洋样品中的12个构成,每个样品都包含了50个以上介形虫个体。
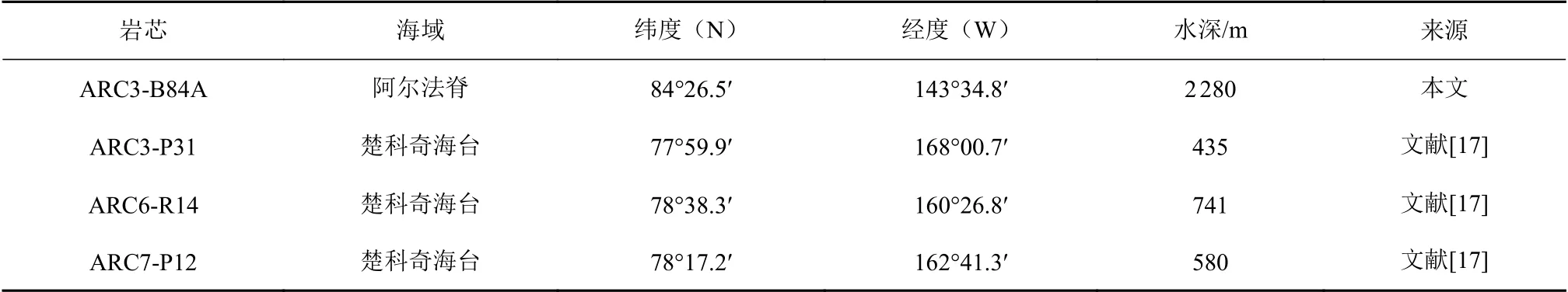
表 1 本研究使用的岩芯及其信息汇总Table 1 Information for all the cores used in this study
2 地层年代框架
B84A岩芯沉积物主要由粉砂质黏土构成。Wang等[27]综合运用AMS14C测年、层序对比、旋回地层学校准等方法,建立了该岩芯可靠的年代框架。具体步骤如下:首先基于AMS14C测年、Ca和Mn元素峰值、地层标志种的分布等,确定氧同位素期次;然后把B84A岩芯的Ca和Mn的stack与LR04深海氧同位素曲线[28]对比,确定年龄控制点后转换为深度–年龄模式;最后分别通过“插值法”获得每个2 cm间隔岩芯样品的年龄。根据年龄模式,该岩芯包含了MIS 1期及MIS 3–13期的沉积记录,MIS 2期缺失。
3 结果及讨论
3.1 B84A岩芯中的介形虫及其生态特性
阿尔法脊B84A岩芯中介形虫化石数量丰富,从94个样品中共挑出介形虫7 011瓣,鉴定出8属11种。化石群的优势种为Polycopespp.(占总个体数量的52.2%)和Cytheropteron sedoviSchneider 1969(占20.7%),常见种为Cytheropteron scoresbyiWhatley and Coles 1987(占4.9%)、Acetabulastoma arcticumSchornikov 1970(占7.0%)、Henryhowella asperrima(Reuss 1850)(占5.5%)、Microcythere medistriatumJoy and Clark 1977(占1.8%)、Pseudocythere caudataSars 1866(占1.9%)、Cytheropteron higashikawaiIshizaki 1981(占1.8%)以及Pedicytherespp.(占1.0%)。有趣的是,北冰洋深海介形虫的代表属之一Krithe[17-19],在该岩芯中仅发现6瓣;而北冰洋大西洋水(AW)的指示者Cytheropteron perlariaHao 1988[17],在该岩芯中仅出现1瓣(42~44 cm孔深处)。图2为部分属种的扫描电子显微镜照片。
基于来自北冰洋638个表层沉积物样品的介形虫数据,本文分析了B84A中的2个优势种和7个常见种在现代北冰洋的分布,以便确定它们与海洋环境、尤其与水团的对应关系。9个属种的百分含量等值线图用ODV绘制,投影在连接加拿大海盆和巴伦支海的横断面上;同时还绘制了介形虫百分含量-水深散点图(图3),为介形虫属种与特定海洋环境的对应关系提供了新认知。9个属种与现代北冰洋水团的对应关系归纳如下(表2)。
(1)北冰洋中层水(AIW)属种:Polycopespp.、Pseudocythere caudata和Pedicytherespp.的百分含量在1 000~2 000 m水深范围内(即AIW内)最高,但在该深度以上和以下明显降低(图3a, c, d;表2)。

图 2 阿尔法脊B84A岩芯中介形虫标本扫描电子显微镜照片a. Polycope biretculata Joy and Clark 1977,右壳瓣;b. Polycope inornata Joy and Clark 1977,右壳瓣;c. Polycope horida Joy and Clark 1977,右壳瓣;d. Polycope moenia Joy and Clark 1977,右壳瓣;e. Polycope arcys Joy and Clark 1977,左壳瓣;f. Acetabulastoma arctium Schornikov 1970,左壳瓣;g. Cytheropteron scoresbyi Whatley and Coles 1987,左壳瓣;h. Cytheropteron sedovi Schneider 1969,右壳瓣;i. Microythere medistriatum Joy and Clark 1977,左壳瓣;j. Pedicythere neofluitans Joy and Clark 1977,右壳瓣;k. Henryhowella asperrima (Reuss 1850),右壳瓣;l. Pseudocyhere caudata Sars 1866,右壳瓣。Fig.2 SEM photographs of ostracodes from core B84Aa. Polycope biretculata Joy and Clark 1977, RV; b. Polycope inornata Joy and Clark 1977, RV; c. Polycope horida Joy and Clark 1977, RV; d. Polycope moenia Joy and Clark 1977, RV; e. Polycope arcys Joy and Clark 1977, LV; f. Acetabulastoma arctium Schornikov 1970, LV; g. Cytheropteron scoresbyi Whatley and Coles 1987, LV; h. Cytheropteron sedovi Schneider 1969, RV; i. Microythere medistriatum Joy and Clark 1977, LV; j. Pedicythere neofluitans Joy and Clark 1977, RV; k. Henryhowella asperrima (Reuss 1850), RV; l. Pseudocyhere caudata Sars 1866, RV.
Polycopespp.营底栖生活,它们在北冰洋代表着一类机会种,在合适的环境中能够快速繁衍[29-30]。以往的研究已经发现Polycope与AIW之间存在着对应关系,并将其应用于古海洋学研究[17-19,29,31-32]。本研究对现代北冰洋数据的分析结果,再次证实了该属对AIW的适应性。
Pseudocythere caudata最早发现于北大西洋的潮下带[33],但是它在北冰洋栖息的水深范围达到3 000 m,且主要分布在AIW中。以前的研究已经发现P. caudata与Polycopespp.在化石群中存在着共现关系[29,34],并发现该种偏好AIW和AODW[17,19]。
Pedicythere属在北冰洋有两个种,即P. neofluitansJoy and Clark 1977和P. arcticaYasuhara et al. 2014。该属零星分布于100~3 400 m水深范围,百分含量通常小于2%,最大丰度出现于1 000~2000 m水深范围,即AIW内。
(2)北冰洋深层水(AODW)属种:Cytheropteron sedovi、Henryhowella asperrima以及C. higashikawai的百分含量在>2 000 m水深范围(即AODW内)最高,而在其他水深范围内明显降低(图3h, e, i;表2)。在AODW内,C. sedovi和C. higashikawai主要分布于>3 000 m水深范围,显示它们适应AODW下部;而H. asperrima则主要分布于2 000~3 000 m范围,说明该种适应AODW上部环境。
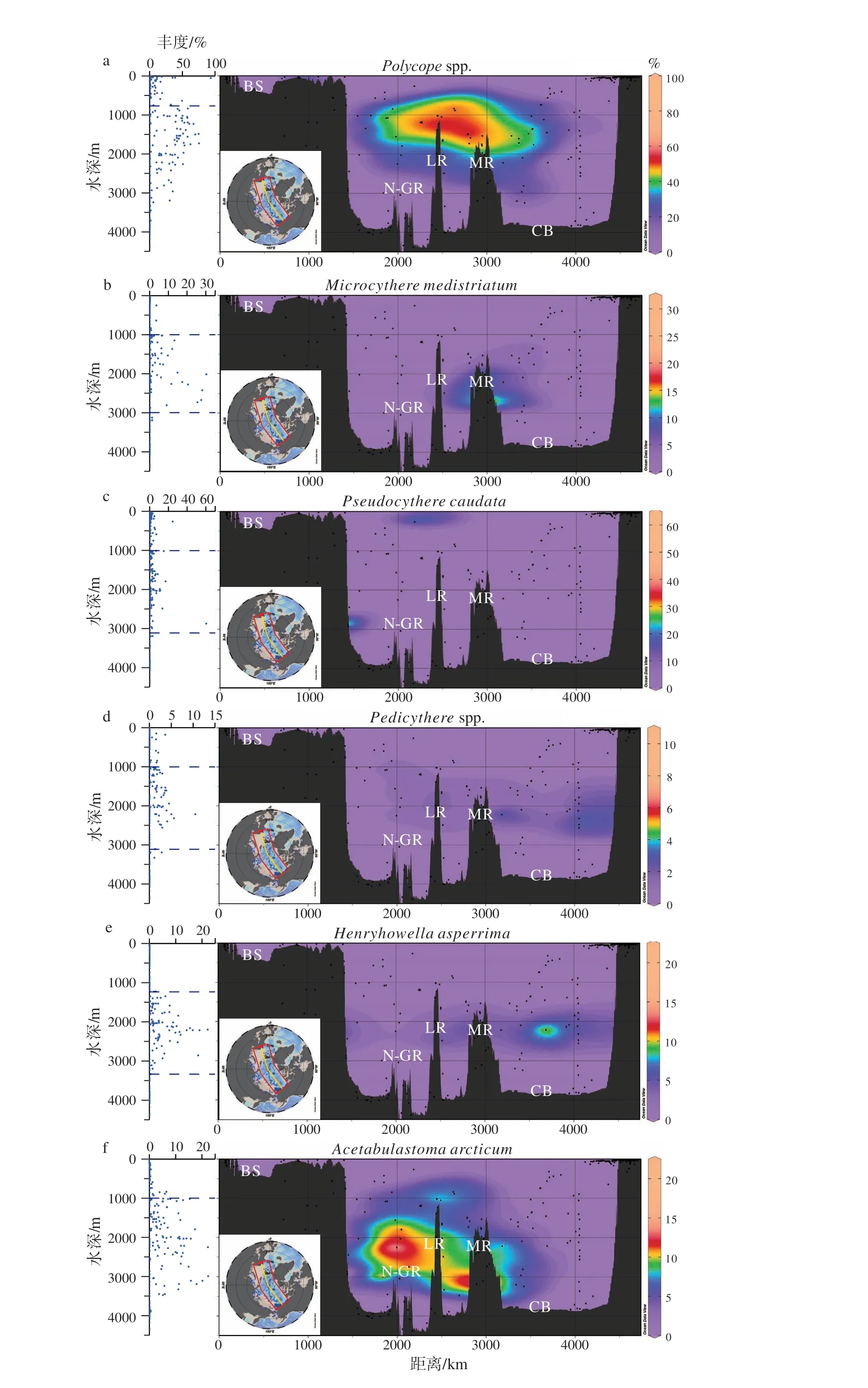
(3)北冰洋中层水和深层水(AIW & AODW)属种:在>1 000 m水深范围(即AIW和AODW内),Microcythere medistriatum和Cytheropteron scoresbyi的百分含量一直保持较高值,而在<1 000 m水深则明显降低(图3b, g;表2)。本研究的分析结果与之前的研究结论[17]一致,即M. medistriatum和C.scoresbyi偏好AIW和AODW。
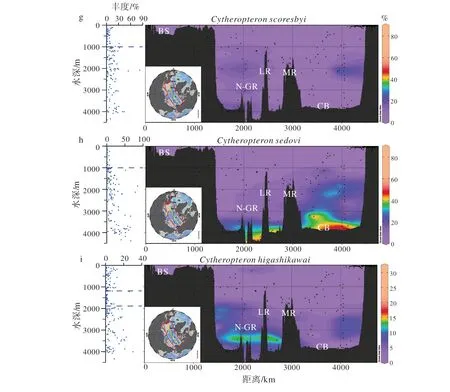
图 3 B84A岩芯中介形虫主要属种在现代北冰洋的分布BS:巴伦支海,N-GR:南森-迦凯脊,LR:罗蒙诺索夫脊,MR:门捷列夫脊,CB:加拿大海盆。Fig.3 Distribution of key ostracode taxa in modern Arctic OceanAbbreviations: BS (Barents Sea), N-GR (Nansen-Gakkel Ridge), LR (Lomonosov Ridge), MR (Mendeleev Ridge), CB (Canada Basin).

表 2 B84A岩芯中的介形虫优势种和常见种在现代北冰洋各水团中的百分含量Table 2 Relative frequencies (%) of ostracode taxa, which are abundant or common in core B84A, in the Arctic water masses
Acetabulastoma arcticum的现生分布仅限于北冰洋中央水域,其百分含量不超过10%(图3f)。该种是一个特例,因为它并非底栖种,而是附着在端足类Gammarus wilkitzkii体表的寄生种,这种端足类在整个生活史都处于北冰洋海冰卤水通道(brine channels)及融化穴(melting holes)中。因此,A.arcticum被视作北冰洋及周边海域常年海冰的指示种[16,30,35],它的分布与水团无关。
以上结果表明,B84A岩芯中的9个介形虫优势属种和常见属种中,除A. arcticum之外,其余8个属种的现生分布均与北冰洋特定的水团相关联,因此可以用于古水团重建的研究。
3.2 MIS 13期以来介形虫化石群变化特征及其影响因素
3.2.1 古生产力与海冰变化
B84A岩芯的94个样品中,介形虫平均丰度为2.9瓣/g,但丰度在不同时期有明显差异。介形虫丰度变化基本上与浮游、底栖有孔虫的丰度变化趋势同步(图4):总体上,丰度在MIS 13–10期很低,平均值仅为0.4瓣/g,而在MIS 9期之后显著升高,平均达到4.3瓣/g。MIS 10期之后,介形虫丰度的高值出现在间冰期的MIS 9、7、5、3、1期,低值则出现在冰期的MIS 8、6、4、2期。微体化石在间冰期多、冰期少的变化趋势在北冰洋似乎是普遍规律[17,30,36-38]。影响北冰洋沉积物中方解石质微体化石丰度的因素有若干个,包括沉积速率、溶蚀作用、生物生产力等;这些因素中,生物生产力似乎是最重要因素,因为本研究(图4)和以往的研究[30]都说明介形虫丰度与氧同位素曲线之间存在着非常好的相关性。本研究结果也证明了北冰洋> 2 000 m水域底栖动物的生产力与较浅水域一样,受冰期–间冰期冷暖变化的影响。
在MIS 13–10期,介形虫和底栖、浮游有孔虫丰度均呈现显著的低值现象。在年代较老的地层中,溶蚀作用可能会损耗更多的介形虫壳体,造成丰度降低的假象,但是我们认为在B84A中并不存在这种可能性,理由有两个:首先,介形虫壳体保存状态在MIS 9期之前和之后没有可辨识的差异,而且壳体相对较薄且易碎的Polycopespp.在MIS 12–11期也有较高的百分含量(图4),说明介形虫化石群在这一时期的保存条件一直是良好的;第二,沉积速率在MIS 13–10期(平均0.42 cm/ka)和MIS 9–1期(平均0.37 cm/ka)之间没有明显差别。因此,有孔虫和介形虫在MIS13–10期的低丰度现象,可能是该时期海洋古生产力显著降低的真实反映。当然,这一推测尚有待北冰洋中部晚第四纪总有机碳分析数据的佐证。
Acetabulastoma arcticum是北冰洋常年海冰的指示种。图4显示A. arcticum在MIS 13–10期几近缺失,在孔深116 cm处(对应MIS 9期)开始较多地出现,因此MIS 9期很可能是常年海冰在罗蒙诺索夫脊以西最早出现的时期,正如Cronin等[30]所指出的那样。然而,尚难以解释为何在常年海冰不存在的MIS 13–10期,由有孔虫和介形虫所代表的海洋古生产力反而降低了。MIS 9之后,A. arcticum在冰期里(MIS 8、6、4期)非常稀少甚至缺失,但在间冰期(MIS 9、7、5、3期)通常占了介形虫总个体数的5%以上,最高达到>30%。在北冰洋中部[18]和西部[30]岩芯记录中,我们发现A. arcticum具有与B84A岩芯中相同的分布方式,即间冰期出现、冰期消失,说明该种及其寄主只能生活在常年海冰和季节性海冰共存的间冰期,而不能生活在洋面完全冻结的冰期环境。
3.2.2 各属种百分含量与古水团变迁
B84A岩芯中,占优势的介形虫属种是Polycopespp.(AIW指示种)和Cytheropteron sedovi(AODW指示种)。MIS 13期以来,这两个属种的百分含量曲线几乎呈镜像对称,二者呈负相关关系且发生了4次较大的转折,据此可以将岩芯记录分为5段,即MIS 13–12期、MIS 11–10期、MIS 9–MIS 5早期、MIS 5中–晚期以及MIS 4–1期。伴随着以上两个优势属种的此消彼长,其他6个与水团相关联的常见种(C. scoresbyi,C. higashikawai,M. medistriatum,Pseudocythere caudata,Pedicytherespp. 以及Henryhowella asperrima)的百分含量也相应发生了显著变化(图4,表3)。5个时期中,各属种的分布及其与水团变迁的关系简要描述如下。

表 3 B84A岩芯中介形虫主要属种指示的水团及其在各时代的百分含量Table 3 Correspondence of ostracode taxa with water masses, and their relative frequencies (%) in different times as recorded in core B84A

图 4 B84A岩芯中有孔虫、介形虫丰度及介形虫主要属种百分含量变化年代框架和有孔虫丰度值引自Wang等[27]。Fig.4 Foraminiferal and ostracode abundances, and relative frequencies (%) of key ostracode taxa in core B84AChronological framework and foranimiferal abundance data are from Wang et al.[27]
MIS 13–12期(约533~424 ka):Polycopespp.和Cytheropteron sedovi百分含量皆约为40%,相当于这两个属种在现代北冰洋2 000~3 000 m水深(即AODW上部)的含量。这一时期,AODW & AIW指示种Cytheropteron scoresbyi也出现,但是含量较低(平均4.3%)。
MIS 11–10期(约424~337 ka):Cytheropteron sedovi丰度升至60%~90%,而Polycopespp.降至约20%,相当于这两个属种在现代北冰洋>3 000 m水深(即AODW下部)的百分含量。这一时期Cytheropteron scoresbyi的含量也较高(平均12.8%)。
MIS 9–MIS 5早期(约337~100 ka):Cytheropteron sedovi降至10%~20%,Polycopespp. 升至60%~80%,相当于处在现代北冰洋1 000~2 000 m的AIW中。另一个AIW指标种Pseudocythere caudata在这一时期的含量也达到最高(平均2.6%)。
MIS 5中–晚期(约100~71 ka):Polycope从>60%降到不足10%,而Cytheropteron sedovi从约10%增加到90%,显示AODW下部(>3 000 m)的水团快速上涌。这一时期里,AIW指示种Pseudocythere caudata(平均0.9%)和Pedicytherespp.(平均1.7%),AIW & AODW指示种Microcythere medistriatum(平均1.1%)和Cytheropteron scoresbyi(平均5.4%),以及AODW下部指示种Cytheropteron higashikawai(平均1.8%)也出现,但是数量都很少。
楚科奇海台介形虫化石群的记录显示,MIS 5中–晚期时,在西北冰洋中层水域(<1 000 m)曾发生过水团上涌事件:下方的AIW向上迁移,占据了原本属于AW的空间[17]。本研究则揭示了同一时期北冰洋中部更深水域(>2 000 m)的水团变迁:AODW向上迁移,将盘踞于此的AIW排挤了出去。MIS 5中–晚期的水团上涌事件,在罗蒙诺索夫脊以西的中层和深层水域可能是联动发生、同时进行的。本研究结果丰富了我们对于北冰洋深层水团变迁的认知,同时也印证了此前的研究结论[17]的可信度。
MIS 4–MIS 1期(约71 ka至今):Polycopespp.含量逐渐恢复到20%~60%,Cytheropteron sedovi则降到约20%,相当于处在现代北冰洋2 000~3 000 m的 AODW上部。这一时期AODW上部指示种Henryhowella asperrima首次出现并且其含量达到了很高值(平均24.0%)。
综上,本研究通过分析来自阿尔法脊的一根岩芯中的介形虫化石群,重建了北冰洋中部深层水团自MIS 13期以来所经历的下沉—上涌—下沉的变迁过程,同时也揭示了古海冰消长和古生产力变迁历史。本研究再次证明介形虫是北冰洋古海洋环境的有用指标,研究结论有待更多的北冰洋微体化石记录以及地球化学指标来进一步证实。
4 结论
(1)由介形虫所代表的底栖生物古生产力在MIS 13–10期很低,进入MIS 9期之后显著升高;在MIS 9期之后,古生产力随气候冷暖交替而增减,间冰期的生产力通常显著高于冰期。
(2)海冰指示种Acetabulastoma arcticum在B84A岩芯中的分布,表明常年海冰很可能是在MIS 9期之后才出现的。
(3)基于AIW指示种Polycopespp.和AODW指示种Cytheropteron sedovi百分含量而重建的阿尔法脊约2 280 m水深处水团变迁历史如下:MIS 13–12期,处于AODW上部;MIS 11–10期,AODW下部上涌,取代了AODW上部;MIS 9–MIS 5早期,上方的AIW下潜,取代了AODW;MIS 5中–晚期,AODW下部快速上涌,将AIW排挤出去;最终在MIS 4–1期,水团定格在AODW上部。其他几个常见种的百分含量变化与上述结论互不矛盾或互为印证。
致谢:本研究使用的岩芯由“雪龙号”科考船所采取。中国极地研究中心样品库为本研究提供了岩芯样品。感谢同济大学张灵敏博士在SEM照片拍摄过程中给予的指导;感谢上海交通大学章陶亮博士在绘制本文插图时给予的帮助。
