不同覆盖作物模式对茶园土壤理化性质及有机碳矿化的影响
2022-08-10龚玲玄王丽丽赵建宁刘红梅杨殿林张贵龙
龚玲玄,王丽丽,赵建宁,刘红梅,杨殿林,张贵龙
农业农村部环境保护科研监测所,天津 300191
土壤是陆地生态系统中最大的碳储存库(Lugato et al.,2021)。全球土壤碳总量为2500 Gt,包括约1550 Gt土壤有机碳(SOC)和950 Gt土壤无机碳(1 m)(Lal,2004a)。世界耕地面积占地球陆地表面的 40%(García-Palacios et al.,2017),而世界土壤腐殖质以每年25.3 Tg的速度在减少(Lal,2003)。覆盖作物(Cover crops,CCs)被美国土壤协会(Soil Science Society of America,2022)定义为“在正常作物生产期间,或在果园和葡萄园的树木和藤蔓之间,提供土壤保护、种子保护和土壤改良的密植作物”,也有一些学者从土壤生态功能的角度出发,认为覆盖作物是能够保持土壤功能(刘晓冰等,2002),为后续作物提供有利条件(Yunusa et al.,2003)的作物,通常在收获与播种季间的空隙种植(Fan et al.,2021),是美国8种“准备实施”的农业管理行动之一(McClelland et al.,2021a)。
目前,虽然覆盖作物能够增加土壤有机碳固持的观点已得到广泛认同,但种植时间和生物量(McClelland et al.,2021b)对土壤有机碳变化动态存在较大影响,且种植时间与土壤有机碳固持间的最佳模式尚未明晰。有观点认为覆盖作物对于土壤碳的影响是一个长期渐进的过程(Ismail et al.,1994;Schipanski et al.,2014),土壤碳库并不能在短期内增加(Lal,2004b),还有些研究认为即使是短时间种植也能使土壤有机碳含量发生显著变化。(Haruna,2019;Faé et al.,2009;Liu et al.,2005)。覆盖作物已被广泛应用在猕猴桃园(李青梅等,2020)、坡地玉米(Leuthold et al.,2021)、葡萄园(Novara et al.,2019)等地区,但多集中于多种单一不同覆盖作物模式-常规作物耕作模式之间(Qi et al.,2011;Hubbard et al.,2013)、单一不同覆盖作物之间(Kuo et al.,1997;Sainju et al.,2002;Kaspar et al.,2006)、多种覆盖作物-常规作物轮作之间(Islam et al.,2006)、单覆盖作物和多种覆盖作物之间(Ranells et al.,1997;Sainju et al.,2007;Blanco-Canqui et al.,2012)的比较,而很少有同时种植多种覆盖作物模式之间的对比,覆盖作物种类多局限于豆科、禾本科,管理方式以休耕、免耕、轮作为主,主要作物多为玉米(ZeamaysL.)、小麦(TriticumaestivumL.)、水稻(OryzasativaL.),很少有覆盖作物与多年生木本植物共生的研究。以往研究中试验区多集中于平原地区,缺乏坡地实践,面对坡地水土流失的环境,短期内覆盖作物的固碳效果与平原地区相比如何,目前尚不明确。
茶作为中国传统的种植作物,拥有悠久的种植历史。中国是世界上第一茶园面积大国,茶园种植也是中国南方丘陵山区的重要土地利用方式(叶菁等,2016)。茶园土壤是一个重要的碳库之一,但新种植的茶园因土壤侵蚀,土壤有机碳(尤其是表层土)处在流失状态(李世玉,2010),短期种植覆盖作物对于茶园土壤而言可能是一种有效途径。基于此,本试验提出两点假设,(1)随着覆盖作物多样性的增加,该坡地茶园表层土壤理化性质改善效果越好。(2)在短时间内,随着覆盖作物多样性的增加,该坡地茶园土壤有机碳矿化作用增加。基于以上研究,从固碳减排角度为该坡地茶园选择覆盖作物搭配的最佳方案。
1 材料与方法
1.1 试验地点概况
试验于湖北十堰郧阳区谭家湾镇圩坪寺村的幼龄茶园开展(图1),该地区属秦巴山区旱地二熟一熟兼水田二熟区,亚热带季风气候区,境内多丘陵,海拔200—300 m,多年平均气温15.4 ℃,平均降水量为769.6 mm,集中于夏季,该茶园坡向为南(157.5°—202.5°)。

图1 实验地点Figure 1 Experiment location
1.2 试验设计
2016年开始每年 3月在幼龄茶园茶行间全面播种覆盖作物,茶树种类为“鄂茶10号”,本试验设计了4种覆盖作物模式(表1),每个模式设置3个重复,每个正方形小区400 m2,共12个试验小区(图2),其中,各小区的茶树间距均为1.5 m,各试验小区中,不同作物均为等量混播,播种密度为25.0 g·m-2,试验期间,及时清除各小区杂草。
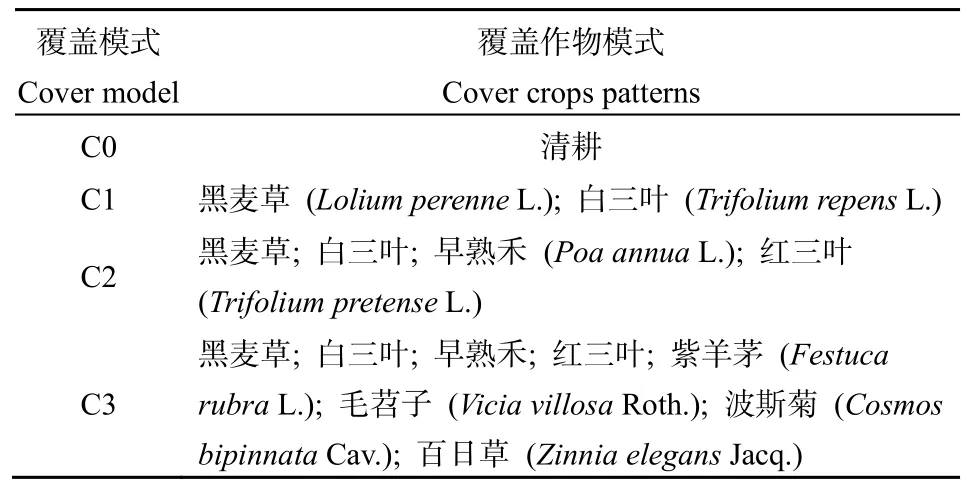
表1 试验期间不同覆盖作物模式Table 1 Different crop coverage patterns during the experiment

图2 试验布置Figure 2 Test layout
考虑到坡地茶园弱酸性的特点,试验选取了8种能够改善土壤结构、增加土壤有机碳、减少水土流失的作物,涵盖禾本科及豆科、菊科,分别为:黑麦草(LoliumperenneL.),紫羊茅(FestucarubraL.),白三叶(TrifoliumrepensL.),红三叶(TrifoliumpretenseL.),早熟禾(PoaannuaL.),毛苕子(ViciavillosaRoth.),百日草(ZinniaelegansJacq.),波斯菊(CosmosbipinnataCav.)(汪洋等,2020)。试验材料中,黑麦草、紫羊茅为多年生禾本科植物,白三叶、红三叶为多年生豆科植物,早熟禾、毛苕子分别为一年生禾本科、豆科植物,百日草为一年生菊科植物,本研究选用的波斯菊为一年生菊科植物。
黑麦草根系浅且密度大,白三叶通过匍匐茎繁殖利于填补覆盖层空隙,红三叶生长迅速,生物量大,固氮能力强,毛苕子耐低肥力,紫羊茅是良好的护坡植物,利于土壤保持,波斯菊、百日草生长速度快,耐贫瘠。考虑成本因素及树冠下光照条件及作物根系特征及生长特性(Snapp et al.,2005),由于茶树种植间距大易有杂草生长,试验选择覆盖模式时将优先按多年生豆科和生长较快的禾本科植物进行搭配。
1.3 样品采集
1 m深度的土壤有机碳一半在可耕层(0—30 cm)(Bolinder et al.,2020),故本试验选择采集 0—30 cm耕层土壤样品。于种植覆盖作物生长良好稳定后(2019年8月底),在覆盖小区内采用“S”取样法在每个小区选取 15个点进行取样,按覆盖模式进行混合,过2 mm筛,将土样置于冷藏箱中带回实验室,一部分用于测定土壤理化性质,土壤含水率采用烘干法测定,硝态氮、铵态氮经CaCl2浸提后使用流动分析仪(AA3,Bran+Luebbe Crop,德国)测定(Houba et al.,1986),微生物量碳(MBC)与微生物量氮(MBN)于25 ℃下经氯仿熏蒸48 h后用TOC仪(Multi N/C3100,德国)测定(Wu et al.,1990);另一部分用于土壤室内培养实验,研究分析土壤有机碳的矿化特征。
1.4 土壤有机碳矿化特征指标和参数的测算
在培养前期,为使土壤恢复自然状态,随机取相当于60 g干土的新鲜土壤均匀置于250 mL玻璃培养瓶底部,于 25 ℃恒温培养箱避光保存一周之后进行土壤有机碳培养,之后分别在第 1、3、5、7、14、21、28、35、42、49、56、63、70、84、98、112、126、140、214、242、269、298、337、471 天时用注射器每次抽取20 mL气体,置于气相色谱仪(Agilent Technologies 7890B)中分析CO2浓度,在每次抽气后将培养瓶放置在通风处20 min,试验重复5次,并设置5个空瓶作为对照,相关采气流程与刘德燕等(2008)一致。
土壤有机碳累计矿化量(CO2-C)采用一级动力学方程来进行拟合,公式为:

式中:
CP——潜在矿化势,表示土壤潜在可矿化有机碳量(CO2-C),单位为 mg·kg-1;
C——在时间t累积土壤有机碳的矿化量(CO2-C);
t——矿化培养时间,单位为d-1;
k——矿化常数,代表有机碳库的周转速率常数(郝存抗等,2020),单位为d-1。
为进一步探究有机碳矿化过程中不同组分碳(CO2-C)的变化情况,使用C.Allan Jones的指数加常数模型公式进行参数拟合,该公式包含两个主要的库,一个是在易分解的活性碳库,在初期矿化,另一个是惰性碳库,分解得更慢。公式为:

其中:
CL——活性碳库的含量(CO2-C),单位为mg·kg-1;
CR——惰性碳库的含量(CO2-C),单位为mg·kg-1;
kR——惰性碳库矿化常数,单位为d-1(Jones,1984;Alvarez et al.,2020)。
1.5 数据处理
采用单因子方差分析(ANOVA)判断不同覆盖作物模式对土壤理化性质的影响,并用 Excel 2019记录测定培养过程中CO2释放量数据和计算土壤矿化速率与累计土壤有机碳矿化量,并在SPSS 25.0中采用Pearson法进行相关性分析。土壤有机碳矿化动态采用Origin pro 2021b进行拟合制图,Loess变化曲线拟合和变异系数均采用R.1.2进行分析制图。
2 结果与分析
2.1 不同覆盖作物模式对土壤理化性质的影响
不同覆盖作物模式土壤理化性质见表2。
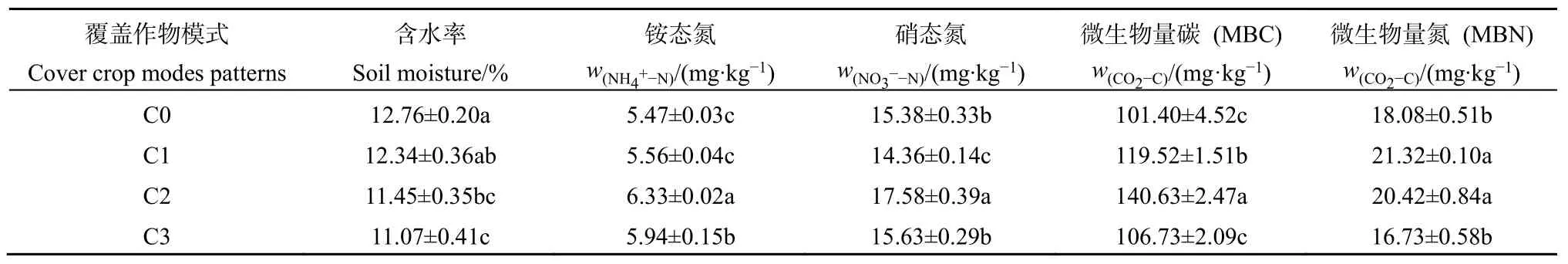
表2 不同覆盖作物模式的土壤理化性质(±SE)Table 2 Soil physical and chemical properties under different cover crop treatments (±SE)
清耕小区(C0)土壤含水率最高,随着覆盖作物多样性增加,土壤含水量逐渐降低。其中,C0模式含水率与C2、C3模式差异显著(P<0.05),但与C1模式差异不显著。
各模式土壤铵态氮含量的相对大小为C2>C3>C1>C0,硝态氮含量为C2>C3>C0>C1,铵态氮、硝态氮、微生物量碳、氮均呈现出“驼峰”模式,即C2的微生物量碳、铵态氮、硝态氮含量最高;C1的微生物量氮含量最高。3种种植覆盖作物的模式土壤含水量比对照低 3.29%—13.24%(平均为8.93%),铵态氮增加了1.65%—15.72%(平均为8.65%),硝态氮增加了-6.63%—14.30%(依绝对值,平均为 7.51%),微生物量碳增加了5.26%—38.69%(平均为20.61%)、微生物量氮增加-7.47%—17.92%(依绝对值,平均为12.78%),从均值来看,覆盖作物对表层土壤微生物量碳的影响最大,其后依次为微生物量氮、土壤含水率、铵态氮和硝态氮。
2.2 不同覆盖作物模式下土壤有机碳矿化特征
为探究短时间不同种类覆盖作物对于茶园土壤有机碳的影响,使用一级动力学方程对土壤培养结果进行拟合。4种覆盖作物模式的土壤矿化速率随培养时间增加均呈“先增后降”的变化模式(图3);在0—3 d,C2的矿化速度高于对照和其他2种覆盖作物模式,C0在63 d、其余覆盖作物模式均于56 d达到最高矿化速率,前者达到峰值的时间与后3者相比延迟了7 d;4种覆盖作物模式最大矿化速率的相对大小为C1>C0>C2>C3。四者在214 d均达到了最小矿化速率。

图3 不同覆盖作物模式下土壤有机碳矿化速率Figure 3 Soil organic carbon mineralization rate under different cover crop models
使用单因素方差对于矿化速率和累计矿化量差异(见图4)进行分析。4种覆盖作物模式矿化速率差异显著(P<0.05),但累计矿化量差异不显著。同时分别对 4种模式进行单因素方差分析,发现C0、C1和C2、C3模式之间差异显著(P<0.05),因此,将4种模式的矿化速率随培养时间的变化曲线分为2类,一类包括C0和C1(整体位于C2、C3上部),为在第 7—337天中矿化速率相对较大型,另一类包括 C2与C3(整体位于下部),为矿化速率相对较小型。为探究4种模式的整体变化过程,采用Loess变化曲线对整个过程进行平滑拟合(平滑度为0.75,见图5)。

图4 不同覆盖作物模式下土壤有机碳累计矿化量Figure 4 Soil organic carbon cumulative mineralization under different cover crop models

图5 4种模式Loess变化曲线,均采用最小二乘法计算Figure 5 Loess variation curves of the four modes, all calculated by the least square method
4种模式的矿化速率平均值分别为 C0:0.24 mg·kg-1·d-1、C1 : 0.25 mg·kg-1·d-1、C2 : 0.22 mg·kg-1·d-1、C3:0.20 mg·kg-1·d-1。3 种含覆盖作物的模式矿化速率平均值比对照组低-4.17%—16.67%(依绝对值,平均值9.72%),随着覆盖作物种类增多,土壤有机碳矿化速率先增后减。为探究不同模式之间矿化速率变化情况,对于Loess变化曲线进行线性拟合,斜率见表3。
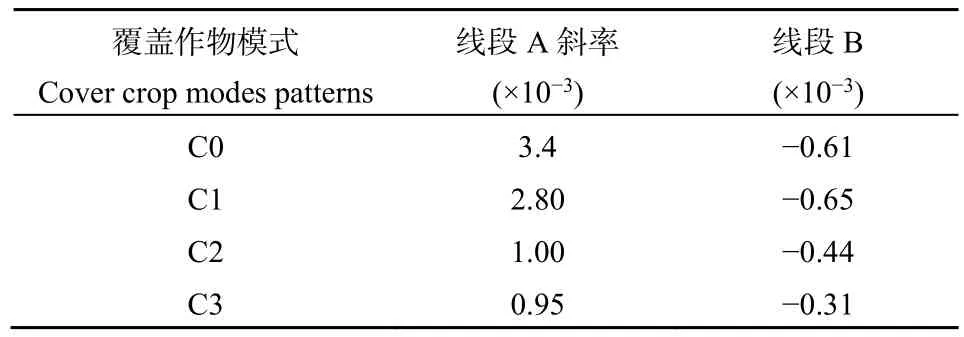
表3 Loess变化曲线线性拟合斜率Table 3 Linear fitting slope of Loess change curve
在初期(1—63 d),3种含覆盖作物的模式矿化速率比对照低 17.65%—72.06%(平均值为53.43%),随覆盖作物种类增多,矿化速率变化逐渐减慢,反应趋于温和。在后期(63—471 d),3种含覆盖作物的模式矿化速率比对照组低-6.56%—49.18%(依绝对值,平均值为27.87%),C1、C2、C3平均值比对照组低12.11%、49.23%、60.62%(依绝对值),表明随覆盖作物多样性的增加,矿化速率趋于“温和”。
变异系数随时间变化见图6。变异系数在21 d达到最大值,为25.56%,之后先减后增,表明4种模式之间的矿化差异逐渐减小,至培养的471 d时,变异系数为4.72%。整个培养过程平均变异系数为12.63%。在1—63 d,平均变异系数为16.85%,比整个过程平均变异系数高33.41%;70—471 d,平均变异系数为8.41%,比整个过程平均变异系数低33.41%;表明在培养的前期,4种覆盖作物矿化速率差异较大,后期差异较小。

图6 4种模式变异系数变化Figure 6 Changes in the coefficient of variation of the four models
4种模式使用一级动力学方程的拟合参数见表4,拟合优度r2均大于0.99,随着覆盖作物种植多样性的增加,土壤潜在可矿化量增多,周转周期减小,CP/k表示碳矿化的强弱,即CP/k越大,矿化作用越强,其变化先减后增。
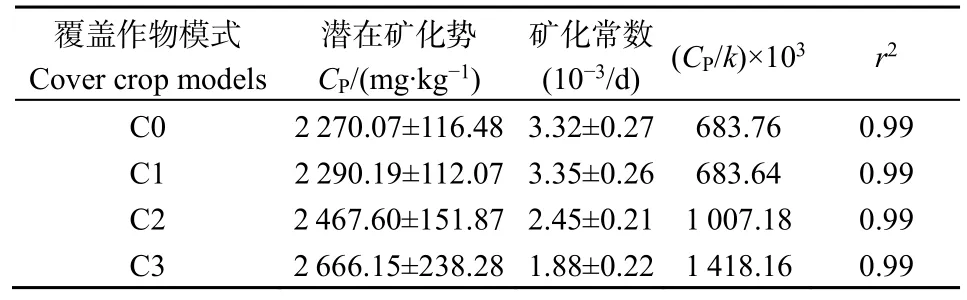
表4 土壤有机碳矿化的一级动力学参数(±SE)Table 4 Parameters of the first-order kinetics for the soil organic carbon mineralization (±SE)
4种不同覆盖模式使用C.Allan Jones公式的拟合参数见表5,拟合优度r2均大于0.99。4种模式均为负值,表明初始矿化出现滞后(Ellert et al.,1988),但随覆盖作物多样性的增加,滞后现象先增后减,至C2滞后现象最小。而惰性碳库不断增加,其周转周期先增后减。

表5 土壤有机碳矿化的指数加常数模型的拟合参数(±SE)Table 5 Fitting parameters of exponential plus a constant model of soil organic carbon mineralization (±SE)
2.3 土壤有机碳矿化特征与土壤理化性质的关系
由表 6,CP与土壤含水率呈显著负相关(P<0.05)。k与土壤含水率呈显著正相关(P<0.05),与kR呈现极显著正相关关系(P<0.01),相关系数为0.99。土壤活性碳库CL与CP、CR呈现出正相关关系,与kR呈现负相关关系。CR与CP呈现出极显著正相关关系(P<0.01),与kR、k呈现出极显著负相关(P<0.01)。土壤的含水率与kR呈现出显著正相关关系(P<0.05),与CR呈显著负相关关系(P<0.05),与CP呈现出负相关关系,其相关系数为-0.954,硝态氮含量与CL呈显著正相关关系(P<0.05),其相关系数为0.958。

表6 土壤有机碳矿化特征与土壤理化性质的相关性Table 6 Correlation between soil organic carbon mineralization characteristics and soil physical and chemical properties
3 讨论
3.1 覆盖作物对茶园土壤理化性质的影响
在C0处理中,土壤中养分含量较低,在种植覆盖作物后,土壤铵态氮、硝态氮含量相较于对照组平均增加8.65%、3.1%,微生物量碳、氮平均增加20.60%、7.8%,使得土壤理化性状也得到提升。有研究表明,种植覆盖作物首先能够增加蒸腾作用,通过减少土壤含水量而减少NO3--N淋溶,从而起到固定氮的效果(Weinert et al.,2002;Quemada et al.,2013),另外种植覆盖作物在土地上增加了生物量,水分利用效率提高,作物氮吸收增加,也减少了NO3--N流失(Vázquez et al.,2005)。另外,有研究表明,覆盖作物能够将接近等比的碳氮分配至地上和地下部分(De Notaris et al.,2020),进入地下后被微生物所利用可增加土壤酶活性(周丽霞等,2007);已有研究表明果园种植覆盖作物每年固定氮素相当于化肥施用氮的 27%—30%的回收率(Ovalle et al.,2010)。
随覆盖作物种类增加,本研究中表层土壤铵态氮、硝态氮、微生物量碳、氮均先增后减。除微生物量氮以外,其余均在C2取得最大值,并与其他模式差异显著,这与王明亮等(2020a)、王明亮,(2020)的研究结论相一致。在C2模式中一年生和多年生禾本科和豆科植物相互搭配,在多年生植物生长之时,一年生植物在死亡后进入土壤进而促进微生物活动,可进一步改善土壤条件。与C1相比,C2多播种了一种禾本科(早熟禾)和多年生豆科作物(红三叶),红三叶相对于白三叶生长速度更快,生物量更大,氮捕获能力强,因此有利于自身和其他覆盖作物的生长。C3中白三叶对波斯菊有化感抑制作用(冯益民等,2018),能限制波斯菊生长,同时菊科内存在对远缘科植物的抑制作用(Kil et al.,1987),可能会抑制其他覆盖作物的生长。
3.2 覆盖作物多样性对茶园土壤有机碳矿化特征的影响
在471 d培养过程中,4种模式土壤有机碳矿化速率均随时间呈“先增后降”的变化。与其他研究在培养刚开始时矿化速率最大所不同的是,在本研究中出现了初始矿化作用滞后的现象(Ellert et al.,1988):除C0在63 d达到最大值外,其余均在56 d达到最大值,这可能与微生物采取的适应策略相关,培养初期微生物进行了重组或适应(Brunner et al.,1984),在培养开始,一部分土壤有机质被仅能分解新鲜有机物的r-策略者所分解,在此过程中r-策略者释放出细胞外酶,初始r-策略者数量增长,在新鲜有机物分解的最后阶段,K-策略者对于聚合有机物进行解聚,并在此过程中释放有机质分解酶,促进土壤有机质的分解(Fontaine et al.,2003),导致最大矿化速率滞后,这可能是本研究培养过程中矿化速率在前期先减后增至最大值,之后减小的原因。
4种模式整个过程平均矿化速率相对大小为C1>C0>C2>C3,差异显著(P<0.05),但累计矿化量差异并不显著。在初期(1—63 d)及后期(63—471 d),4种模式平均矿化速率表现为C1>C0>C2>C3,初期4种模式的平均矿化速率较后期高30.30%。在初期,可能是由于土壤中包含覆盖作物和茶树根系分泌物及各种凋落物等易分解的物质,加之初始N、P、K浓度较高,使得微生物活性逐渐增加(刘义平,2011;徐阳春等,2002),矿化速度较快(郭剑芬等,2006),出现了矿化速率最大值,这也可能是3种含覆盖作物的模式最大值提前7 d的原因。在消耗完最初的易分解化合物之后,微生物开始缓慢分解根系木质素,导致后期土壤有机碳的矿化速率下降(Li et al.,2020),反应趋于平缓。在本试验中发现C0、C1在第7—337天矿化速率相对较大,整体位于 C2、C3上部,C2、C3相对较小,位于下部,C0、C1与C2、C3之间差异显著(P<0.05),表明随覆盖作物种类增多,矿化反应越“温和”。
随覆盖作物种植种类的增加,潜在矿化势先减后增,矿化作用不断增强,这可能与覆盖作物植物根系对于养分及微生物量的富集效应有关。菊科优于豆科,其次是禾本科(杨阳等,2015),本实验发现随覆盖作物多样性增加,土壤有机碳的固存能力也在增强,表现为惰性碳库含量不断增加,这可能与C3含有更多种类的禾本科和菊科植物有关。高纤维素使其更易固存(Redin et al.,2014),同时禾本科植物发达的根系,能够固存土壤中活性碳及总碳含量(Rosolem et al.,2016)。这与汪洋等(2020)的2种覆盖作物矿化作用最强结果不同,本研究中随着覆盖作物种类增多,矿化作用增强,即 8种覆盖作物矿化作用最强,这可能与土壤深度及培养周期差异所致。
浅层土壤与覆盖作物植物根系作用频繁,更易接受覆盖作物物质投入,相比之下,深层土壤理化性质变化及土壤有机碳固存更为缓慢(表7),因此本试验选取 0—30 cm表层土壤,但有研究表明微生物活性可能受底层土壤氮、磷的控制(Meyer et al.,2008),未来还应开展更深层(1 m土壤剖面)的土壤有机碳矿化及微生物活性变化动态研究,以便更准确全面的分析覆盖作物对茶园土壤固碳减排的影响。
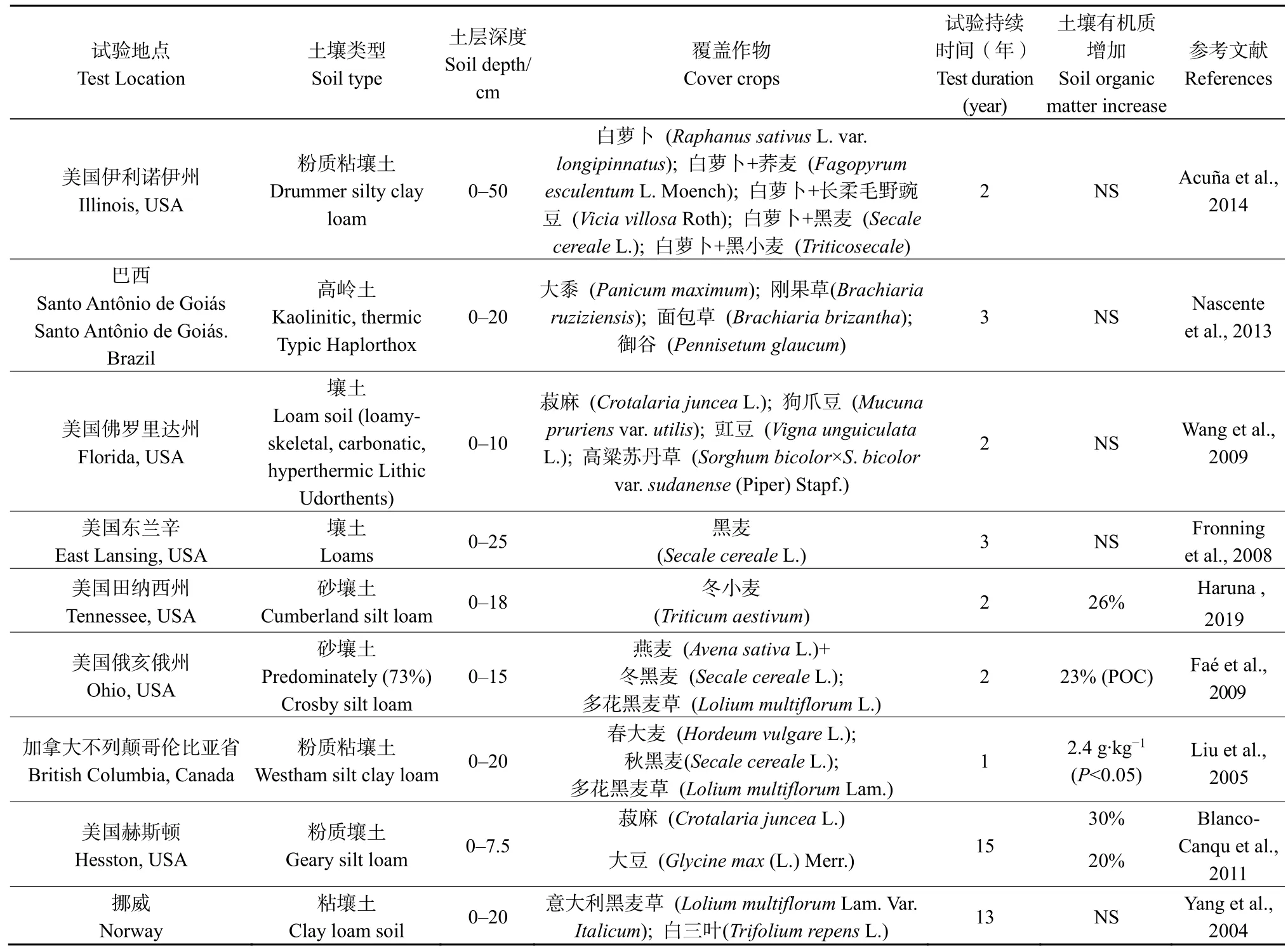
表7 覆盖作物对土壤有机碳影响的效果Table 7 Effects of cover crops on soil organic carbon
4 结论
(1)在本试验中,随覆盖作物多样性的增加,表层土壤(0—30 cm)铵态氮、硝态氮、微生物量碳、微生物量氮含量先增后减,除微生物量氮外,其余均在C2出现最大值,表明过度增加覆盖作物多样性并不能起到更好改善土壤理化性质的效果。
(2)在试验中4种模式均出现了表层土壤矿化滞后现象,C0在63 d达到最高矿化速率,其他模式均在56 d达到最高矿化速率。
(3)在短时间内,随覆盖作物多样性的增加,该坡地茶园表层土壤惰性有机碳含量增加,但矿化作用在增强。
(4)在本试验中,C2能够显著改善表层土壤理化性质,同时也能够在一定程度上促进表层土壤有机碳固存,为该坡地茶园最佳的覆盖作物方案,在农业生产实践中,若既需增加土壤有机碳含量又需实现土壤性质的改良,可适当设置一定的覆盖作物种植种类,以增加土壤惰性碳库含量,进而最大程度地改善土壤养分并实现固碳减排的目的。
