阴山北麓农牧交错区退耕地草地生态系统碳交换及水分利用效率
2022-08-03王赟博赵清格赵萌莉
王赟博,孙 宇,赵清格,张 彬,赵萌莉
内蒙古农业大学草原与资源环境学院草地资源教育部重点实验室, 呼和浩特 010018
陆地生态系统碳交换是全球碳循环的重要组成部分。作为占陆地面积25%的草地生态系统,其巨大的碳库极易受到人类活动的影响而发生碳源-碳汇的转变[1—4]。特别是处于我国阴山北麓农牧交错带的荒漠草地,由于常年缺乏降水,植被稀疏,加之多年的盲目开垦和过度放牧,草地植被破坏严重,碳固持等生态系统功能显著下降,是我国主要的生态脆弱地区之一[5—6]。因此,合理的退耕还草,恢复草地原有植被和功能便成为了改善当地生态环境,维持碳汇潜力的关键措施[7—9]。针对草地的退耕恢复演替,以往的研究大多集中在一定恢复时间后地上植被组成变化以及土壤环境因素变化等方面,对于退耕地生态系统碳交换及其恢复过程的研究则较为缺乏[4,10—12]。这无疑会增加对未来荒漠草地碳汇潜力的预测的不确定性。随着我国土地利用政策的改变,内蒙古阴山北麓退耕还草面积也会进一步增加[13],急需理清在植被、土壤以及气候等诸多复杂关系因素干扰下荒漠草地生态系统碳交换和水分利用效率对退耕恢复的响应规律[14]。因此,本研究主要通过测量不同弃耕恢复阶段荒漠草地植被组成、生物量、生态系统群体光合以及水分利用效率,最终阐明弃耕荒漠草地生态系统碳循环的恢复过程及其调控机制,为在阴山北麓农牧交错区的退耕还草政策的实施以及该区域生态功能恢复情况评估提供理论依据。
20世纪90年代起,我国提出了保护生态脆弱区基础生态功能和价值的政策,随后西部地区大范围实施退耕还草工程,产生了大面积的退耕恢复草地。1999—2019年,全国有大约3400万公顷不适于耕作的农田被人为废弃[15],且由于经费等原因其中多数退耕地只能通过缓慢的自然群落演替恢复其草地原有的生态结构和功能。对于退耕地生态功能的恢复效果和过程以往的研究已多有报道,多数认为退耕还草会显著改变当地的植被组成,提高植物多样性和生产力,并最终刺激生态系统的碳交换及碳固持能力[11,16—19]。Heikkinen等针对前苏联遗留的弃耕地研究发现弃耕20年后草地土壤碳库提高了大概1.5—1.8倍[20]。Paustian甚至认为莫斯科周边的弃耕地所吸收固定的碳足以抵消城市发展所造成的碳排放增量[7]。相类似的结果在我国黄土区以及甘肃等地也有发现[21—22]。这说明退耕地已成为区域生态系统中重要的碳汇,在碳循环以及碳平衡过程中扮演着关键的作用[14]。此外,由于荒漠草原长期处于水分限制状态,退耕地恢复过程中植被对于水分的吸收和利用策略在种间竞争以及植物物种演替中的意义就显得极其重要[23—24]。通常多年生植物往往有更高的水分利用效率以及耐旱能力[25]。因此,随着退耕恢复地群落由一、二年生先锋植物向多年生植物演替[26],群落的水分利用效率也必将受到影响。而植物水分利用效率改变导致的土壤水分可利用性变化以及不同地区及年际间降水的变化使得退耕还草对草地生态系统水分利用效率的影响机制更加复杂[21]。
为了进一步理解内蒙古阴山北麓农牧交错区退耕还草对恢复草地生态系统碳循环的影响,本实验在2020年8月对内蒙古武川县上秃亥乡周围不同弃耕年限恢复草地生态系统碳交换、水分利用效率以及其他相关指标进行了实地测量。并试图回答如下问题:(1)生态系统CO2交换和水分利用效率对不同退耕年限自然恢复处理的响应规律;(2)退耕恢复草地生态系统CO2交换和水分利用效率改变的主要影响因子。
1 材料与方法
1.1 研究区概况
实验地位于内蒙古阴山北麓的武川县上秃亥乡(41°10′59″N,111°18′6″E,海拔1604 m),该区域为武川县退耕还草主要试点区域之一。该区域农田和天然草地呈斑块状交错分布,属于典型的农牧交错区域(图1)。自2002年起该区域分多次组织进行了退耕还草工程,多数退耕地位于坡度大于15°的山坡地带。
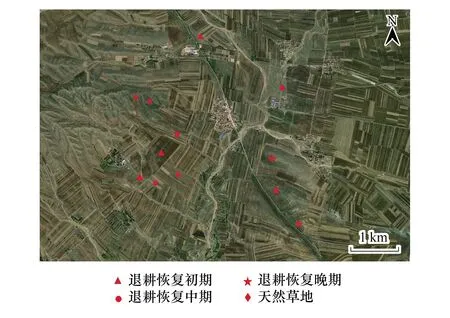
图1 研究地点的地理位置图Fig.1 Geographic location of the study site
实验地所在区域属典型温带干旱半干旱季风气候,常年干旱少雨,年均温度2.5 ℃,年均降水250—360 mm,春秋季降水偏少,降水主要集中于6—9月[13];土壤类型为栗钙土,该区主要天然草地类型为荒漠草原,除禾本科植物外,该地区还有大量菊科植物,豆科植物相对比例极少。除短花针茅(Stipabreviflora)之外的优势种和常见种包括:糙隐子草(Cleistogenessquarrosa)、冷蒿(Artemisiafrigida)、点地梅(Androsaceumbellata)和栉叶蒿(Neopallasiapetinata)等。
1.2 实验设计及样地选择
通过走访调查当地农户以及查阅武川县近20年的退耕还林还草记录等文件,按照上秃亥乡周围退耕还草斑块退耕起始时间将其分为退耕恢复初期(PR,2013—2014年)、退耕恢复中期(MR,2007—2008年)和退耕恢复晚期(AR,2002—2003年)3种退耕恢复梯度(图2)。2019年8月上旬在当地面积较大、连续性较好的退耕地中选取土质基本相同,植被生长良好,且无明显放牧或其他人为干扰痕迹的3种退耕恢复梯度样地各3块样地;同时,选取3块与所选退耕地坡度和土壤等较为一致的天然草地样地作为对照样地。在每个样地典型地段坡度一致的阳坡向随机选取1个10 m×10 m的区域作为实验小区,共得实验小区12块。

图2 不同退耕恢复年限样地实景Fig.2 Photos of the sample plot
1.3 植被调查
实验于2020年8月上旬生物量高峰期时进行,在各样地内随机选取5个1 m×1 m的样方,调查每个样方内的植被物种组成及分种盖度情况;然后将样方中各植物物种分种齐地面刈割,迅速带回实验室称量各物种鲜重。放入烘箱105℃杀青2 h,后70℃烘干至恒重并称量其干重作为各物种地上生物量(AGB)。通过公式(1)计算各物种植物地上组织含水量(PWC)。
PWC=(FW-DW)/FW
(1)
PWC代表植物地上组织含水量,FW代表植物地上组织鲜重,DW代表植物地上组织干重。我们将调查到的所有植物种类按其生活史特性分为多年生植物和一、二年生植物,并分别计算其各自的AGB和PWC。
1.4 生态系统碳交换及水分利用效率
在本实验中,生态系统碳交换主要由净生态系统CO2交换(net ecosystem CO2exchange,NEE),总生态系统生产(gross ecosystem productivity,GEP)和生态系统呼吸(ecosystem respiration,ER)组成。弃耕地NEE和ER通过Li- 6400XT光合仪(LI- 6400, LiCor Inc., Lincoln, NE, USA)配合自制光合箱(50 cm×50 cm×50 cm)在野外实地测量取得,GEP则通过公式(2)计算得出[27]:
GEP=NEE+ER
(2)
实验测量选择在2020年8月上旬无风晴朗的天气下进行,测量时间安排在每天的上午7点到11点。在每个实验样地中的典型斑块内随机选择3个样点,分别对该样点植被生态系统CO2交换相关指标(NEE和ER)以及生态系统水分蒸腾指标(ET)进行测量。通过公式(2)和(3)计算分别计算GEP和水分利用效率(WUE)。
WUE=NEE/ET
(3)
将以上指标在每个样地中3个样点测量值的平均值作为该实验样地的平均生态系统碳交换和平均水分利用效率。
本实验生态系统碳交换测量时间和方法与国内外多数相同或类似实验基本一致。期间地表空气温度变化在19.8—26.3℃之间,同一样地土壤水分变化不显著。尽管温度存在显著变化,但由于其均在多数旱生草本植物适宜的光合温度范围内(19—28℃),因此实验期间空气温度和土壤水分变化对实验结果的影响极小。
土壤10 cm土层含水量(SM)使用便携式土壤水分速测仪(HH2/WET, Delta-T Devices Ltd, Burwell, Cambridge, United Kingdom)随生态系统碳交换测量一同进行。
1.5 数据处理
实验采用单因素方差分析(one-way ANOVA)对不同恢复阶段退耕地的NEE、GEP、ER、WUE、PWC、SM和AGB进行差异显著性检验,并通过Duncon法进行事后检验。通过Pearson相关性分析,得到各主要解释变量与目标指标的相关程度,明晰生态系统CO2交换和水分利用效率变化的主要驱动因素。
2 结果与分析
2.1 退耕地土壤含水量及植物含水量的变化
退耕恢复初期和中期样地的平均植物地上组织含水量(PWC)显著高于恢复晚期和天然草地样地(P<0.05)。同时,各恢复阶段样地间的多年生植物或一、二年生植物的PWC都未有显著的差异(P>0.05),而在所有处理样地中多年生植物均显著低于一、二年生植物的PWC(图3)。此外,表层土壤含水量随弃耕地恢复时间增加而呈先减小后增大的趋势,但均显著小于天然草地斑块,且土壤含水量最小值出现在退耕恢复中期样地(图3)。
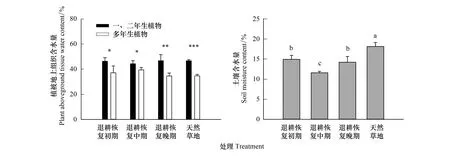
图3 各样地多年生和一、二年生植被地上组织含水量以及土壤含水量差异Fig.3 The mean of annuals and perennial aboveground tissue water content and soil moisture content*、**和***分别代表在P<0.05、P<0.01和P<0.001水平上存在显著差异; 不同小写字母代表各退耕恢复阶段样地间土壤含水量在P<0.05水平上存在显著差异
2.2 退耕地物种组成及地上生物量的变化
本研究样地调查样方中共出现38种植物,其中天然草地包含物种数最多(24种),退耕恢复初期处理最少(14种),物种数量随退耕恢复时间延长呈增加趋势(表1)。短花针茅、栉叶蒿、草麻黄和二裂委陵菜等4种植物在所有处理样地中都有分布。同时,尽管各样地所包含的一、二年生植物种类并不一致,但所有处理中一、二年生植物物种数量差异并不显著(P>0.05)(NG:5种、AR:4种、MR:6种、PR:5种);而多年生植物物种数在恢复晚期和天然草地样地(NG:19种、AR:19种)要远高于其他两个处理样地(MR:12种、PR:9种)。

表1 各样地植被物种分布及生活型分类Table 1 The distribution and life type of species in sample sites
地上生物量(AGB)随恢复时间延长呈增加趋势,退耕恢复晚期样地与天然草地地上生物量已无明显差异(P>0.05),但均显著高于退耕恢复中期和早期样地(P<0.05)(图4)。不同恢复阶段样地一、二年生及多年生植物地上生物量有极显著的差异(P<0.05):弃耕地恢复时间延长会显著降低一、二年生植物地上生物量,但均高于天然草地(P<0.05);而不同弃耕恢复阶段对多年生植物地上生物量的影响则正好相反(图4)。
2.3 退耕地生态系统净CO2交换和水分利用效率的变化
实验结果表明,不同退耕恢复阶段间生态系统碳交换存在显著的差异(P<0.05)。天然草地总生态系统生产(GEP)和生态系统呼吸(ER)均显著高于退耕恢复初期样地(P<0.05),而与退耕恢复中期和晚期样地并无明显差异(P>0.05)(图5)。同时,随退耕地恢复时间延长,退耕地生态系统CO2交换(NEE)存在明显的恢复趋势:与天然草地相比,NEE分别恢复了32.8%(PR),76.5%(MR)和82.0%(AR),退耕恢复晚期样地已基本恢复到天然草地的生态系统碳固定能力水平(图5,P>0.05)。与退耕地NEE结果类似,不同退耕恢复阶段样地水分利用效率(WUE)也呈升高的趋势,退耕恢复初期生态系统WUE显著低于恢复中期和晚期样地,而天然草地生态系统WUE则明显高于所有退耕恢复样地(P<0.05)(图5)。
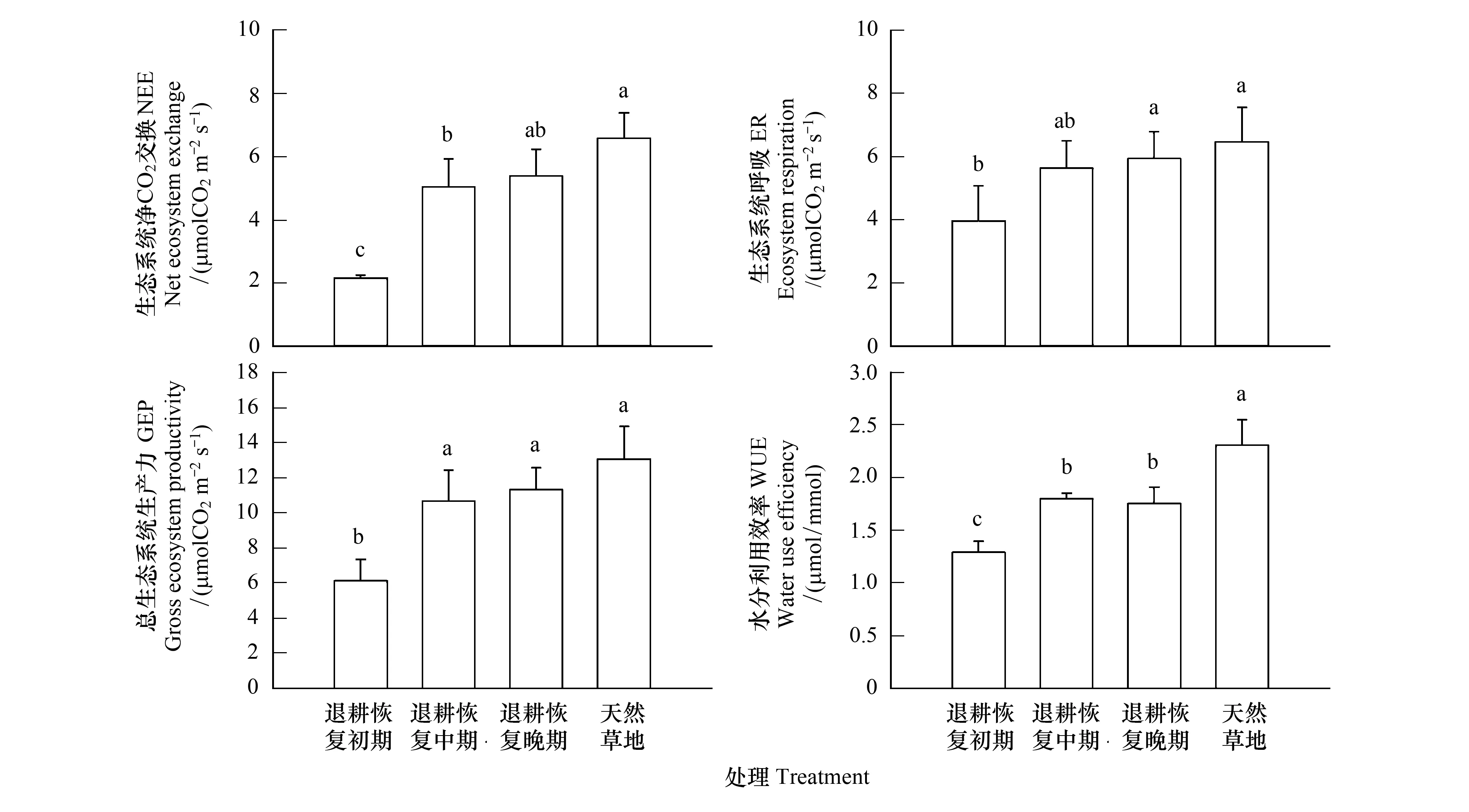
图5 实验处理对生态系统净CO2交换、总生态系统生产、生态系统呼吸和水分利用效率的影响Fig.5 The means of net ecosystem CO2 exchange, ecosystem respiration, gross ecosystem productivity and water use efficiency under experimental treatments
2.4 生态系统净CO2交换、水分利用效率与植被组成的相关性
通过对所有样地的综合分析,实验发现生态系统NEE和ET与样地植被总地上生物量(AGB)极为相关(P<0.01)。这些相关关系可以用一元线性模型很好地描述(图6)。此外,本实验还发现生态系统WUE与一、二年生植物地上生物量呈负相关关系,而与多年生植物地上生物量呈正相关关系(图6)。
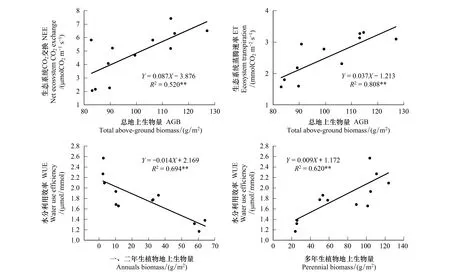
图6 生态系统CO2交换和生态系统蒸腾速率与总地上生物量的相关关系;以及生态系统水分利用效率与一、二年生植被地上生物量和多年生植被地上生物量的相关关系Fig.6 Positive dependence of growing season mean net ecosystem CO2 exchange and ecosystem transpiration on total above-ground biomass. And responses of water use efficiency to annuals biomass and perennial biomass*、**和***分别代表在P<0.05、P<0.01和P<0.001水平上存在显著相关
3 讨论
对于多数干旱半干旱地区退耕恢复草地而言,由于环境水分可利用性的长期缺乏以及由此导致的生态系统营养循环速率的减慢[17,28],其恢复演替周期可能往往需要十几年甚至更长的时间,加之降水、温度等对植被生长和光合过程极为重要的环境因素随年际波动等原因,实际研究中经常难以得到较好的长期观测结果。因此,本实验采用空间序列代替时间序列的方法[9,12]:在同一区域中,选择不同恢复演替阶段的草地,以模拟退耕地恢复演替的过程。并通过选择距离较近,土壤质地和坡度相似,且无人为干扰的样地以减少不同空间差异带来的影响。
3.1 不同恢复演替年限对退耕地生态系统净CO2交换的影响
随着社会经济和人口的发展,不合理开垦等人为干扰事件的增多,我国特别是内蒙古阴山北麓农牧交错地区大量的天然草地生态系统受到了严重的破坏[4,13,29]。这不仅会增加当地草原生态系统水土流失风险,降低植被物种多样性,更显著影响着这些地区的主要生态系统功能(如生态系统碳交换和碳固持能力)的正常实现[12,29—30]。因此,适度的退耕还草成为了保护当地草地生态系统,维持草地正常碳汇功能稳定的重要手段[31]。
通常而言,农田退耕后,随着人为干扰的消失以及农耕植物的减少,当地野生物种会逐渐填补退耕地的空缺生态位并改善土壤营养、水分供给环境,进而促进植物生长,最终影响草地碳交换及其潜在固碳能力[11,16]。以往的研究尽管大都表明退耕恢复处理会显著恢复受破坏草地生态系统的碳循环水平[18,32],但也存在一定的争议,不变甚至降低的实验结果仍有所报道[3]。这些差异的产生可能与实验本身退耕持续时间、当地环境因素以及植被组成的不同有关。例如安慧等在宁夏荒漠草地生态系统恢复的研究中即发现由于退耕初期植物种类以及土壤水分可利用性限制等原因,恢复草地群落植被生物量呈先减少后增加的趋势[17]。而在本实验中发现丰雨年荒漠草地退耕恢复生态系统CO2交换随退耕演替年限增加呈明显的上升趋势,且在恢复晚期群落NEE已与天然草地无明显差异(图5)。相似的现象在一些更为湿润的草地生态系统中也有报道[33]。这一现象可能是退耕处理后植被盖度和生物量的显著增加导致叶面积以及光合单位数量提高的结果。事实上,本实验表明退耕地群落NEE与生物量呈显著的正相关关系(图6)。这些结果表明,至少在水分供给充足的环境下,荒漠草地仅仅通过植被的自然恢复,退耕一定年限后即可对草地生态系统结构和功能产生极为深刻的影响,有效地促进植被生长和光合,并最终恢复草地生态系统的固碳功能。
此外,值得注意的是退耕恢复处理导致的NEE升高主要是因为草地生态系统GEP的恢复速率远高于ER导致的(图5)。由于ER通常与生物量及其活性密切相关,而草地生物量及其活性又极易受到土壤关键营养(如氮和磷素)供给不足的限制,所以在退耕恢复过程中草地可能更倾向于优先恢复其生态系统光合固碳潜力,而非改善土壤营养供给水平及其利用效率。对于绝大多数时间都处于水分胁迫的荒漠草原生态系统而言,这一选择可能有利于在短期环境适宜时植被能够固定更多的碳素,从而为草地生态系统恢复过程提供足够的碳源支持。然而,对于退耕恢复处理影响生态系统碳交换的研究仍处于起步阶段,其内在机理仍不清晰,恢复过程中,弃耕地初始条件变化、年降水量的不同和植被类型差异都有可能对实验结果产生影响。未来需要设计和实施更多包含降水梯度(或年际变化)的更为缜密的实验。
3.2 不同恢复年限对退耕地生态系统水分利用效率的影响
除了生态系统碳交换外,植被水分利用效率(WUE)的变化也在退耕草地生态系统恢复过程中有着至关重要的作用[34]。特别是在荒漠草原中,由于当地高蒸散和低降水的共同作用,植物的生存、生长和繁殖长期处于水分限制状态[35]。因此,植物对水分获取和利用策略的不同可能直接影响着群落物种组成以及退耕地恢复的过程。与实验预期相符,本实验长期弃耕处理极大地影响了草地生态系统WUE的水平,退耕恢复中期和晚期植被WUE显著高于恢复初期,但均显著小于天然草地(图5)。这一现象可能主要与退耕恢复草地中不同生活型植物的组成比例及其水分利用策略差异有关。通常而言,作为典型的r-对策者,一、二年生植物由于其较高的植物含水量,较浅的根系分布和较短的生命周期,在适宜的环境下更倾向于通过提高气孔导度,促进植物叶片CO2和水分交换,从而在短期内获得较高的光合及土壤水分获得能力,但这无疑会极大地降低了植物的WUE水平;而多年生植物则更注重于改善自身对水资源的利用以适应较长生命周期中可能频繁出现的干旱事件[36]。因此,荒漠草原弃耕恢复地一、二年生和多年生植物的生长状况在一定程度上决定了整个生态系统WUE的变化。事实上,在实验中退耕恢复地群落NEE与多年生植物生物量呈正相关,而与一、二年生植物呈负相关的现象印证了这一观点(图6)。同时,不同恢复阶段群落WUE的差异也会导致土壤水分的可利用性的改变。我们的实验发现在WUE较低的退耕恢复地的土壤含水量也显著低于天然草地(图3),这无疑会影响其植物的生存和生长,并提高更为耐旱的多年生植物的竞争优势,促进其所占比例的增加。本实验的这些发现凸显了退耕地群落生态系统功能恢复过程的复杂性,以及植被恢复及演替在这一过程中的重要作用。
3.3 年际降水格局差异对不同恢复年限草地生态系统NEE和WUE变化规律的影响
降水格局变化是草地植物群落结构和功能的主要影响因素之一[27]。特别是对于大多数干旱缺水的草地生态系统,由于年际间生长季降水量和降水分布差异导致的土壤水分可利用性变化往往会显著影响生态系统植被生长、碳交换和水分利用效率[37—38]。一般而言,由于长期处于水分限制状态,降水增加可以通过改变植被盖度、叶面积和叶片气孔开合程度等刺激荒漠草地植物生产力和NEE的提高,但过多的降水往往会抑制群落植被水分利用效率[39—40]。然而,草地不同物种对水分变化的喜好和适应性差异,不同植物物候期与环境水热供需耦合程度差异等因素的存在使我们对不同弃耕恢复年限草地群落响应降水格局变化的判断变得更为复杂和不确定[40]。以往大量研究认为:一、二年生植物对降水改变的敏感性要显著高于多年生植物;而生长季降水分布也可能对当年草地植被组成和生长情况产生影响,草地生长季前期(5月—6月)的干旱事件会严重抑制多年生草本的生长,并最终导致水分供给改善后一、二年生植物的爆发性生长以及其占总体生物量比例的提高[36,37,41—43]。在本实验中,实验期间尽管生长季降水略高于当地多年平均降水量(249.2 mm),但生长季前期降水偏低,这一降水模式可能有利于弃耕恢复草地植被盖度的提高以及一、二年生植物的生长。因此,实验基于单一生长季测量的实验结果可能对不同恢复年限草地生态系统(特别是一、二年生植被较多的恢复前期和恢复中期)NEE的恢复程度存在一定的高估,而低估了这些样地的原本的WUE水平。由于本实验用空间序列代替时间序列的实验方法以及实验人力和设备条件等因素限制,本文缺乏包含对实验样地相关指标多年持续监测的数据,由此得出的结果可能会在总降水偏少或生长季前期降水偏多的年份下对草地NEE和WUE的估计造成一定偏差,但对弃耕恢复草地功能变化的总体趋势判断影响不大,本实验结果对荒漠弃耕草地恢复过程研究仍具有很强的理论和实践意义。未来仍需设计和实施更为完善,实验周期更长的相关实验。
4 结论
内蒙古阴山北麓农牧交错区农田退耕还草后,草地生态系统CO2交换得到了明显的恢复,特别是在植被水分限制较轻的年份,退耕恢复晚期群落NEE已基本恢复到天然草地的正常水平,其中草地恢复演替过程中地上生物量及由此导致的群落盖度的显著增加是一个主要原因。同时,草地生态系统WUE也随退耕恢复演替的进行而呈提高的趋势,这可能主要归功于群落演替引起的多年生与一、二年生牧草比例的变化,但是17年的时间似乎并不足以完全恢复其原有程度。因此,内蒙古荒漠草原地区退耕恢复草地不同生态系统功能的恢复过程可能并非是同时性的,草地生态系统可能优先选择提高植被群落对碳的固定潜力,而减缓对生态系统水分和养分的供给和利用能力的修复。
