线粒体和真核生物的起源
2022-07-29编译莫庄非
编译 莫庄非
生命起源后的数十亿年间,地球上唯一的生命体是微小的原始细胞——类似于今天的细菌。不过在15亿年(可能更久)前,发生了一桩了不起的故事:这些原始细胞中的一种古细菌,吞下了另一种古细菌。
被吞噬的细菌没有被消化,而是永久居住对方体内,生物学家称之为内共生体。最终,小胞体完全整合至古生菌大胞体,成为我们今天所熟知的线粒体,即细胞的产能器。
生命起源之后最重大的演化飞跃,是从早期原核生物向更复杂真核生物的转变,而这场飞跃中最重要的一步,是线粒体的获得。
上述内容是你在大多生物教材里都找得到的简介,但要把简介内容展开来细究,故事就变得错综复杂。
过去几年间,新证据的出现对“线粒体在原核-真核转变过程中发挥重要作用”的观点提出了挑战。研究人员曾对第一批真核生物的现代亲属进行基因组测序,结果发现许多意想不到的基因,它们似乎既非宿主之物,也不是来自内共生体。一些科学家指出,这可能意味着第一批真核生物的演化涉及两个以上的“合作伙伴”,而且发生过程比想象的更慢。
其他学者则认为暂时还没理由放弃老理论,获得线粒体可能就是点燃真核大跃进的那把火——亿万年后,植物、动物、脊椎动物以及人类出现了。来自基因组学和细胞生物学的新证据可能有助于解决这一争论,同时也指出了认知复杂细胞起源所需填补的知识空缺。
神秘参与者的额外贡献
过去十年间,巴塞罗那超级计算中心的演化基因组学家托尼·加巴尔东(Toni Gabaldón)和同事利用基因测序技术探索各种真核生物的基因组,其中包括一些早期真核生物的现代近亲,并发现了神秘基因。
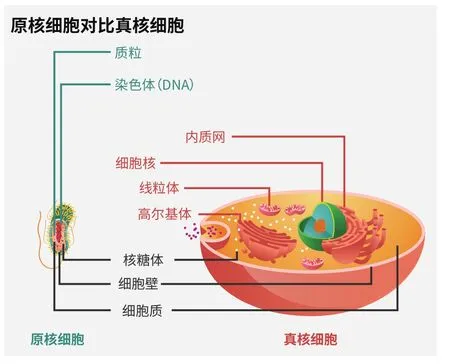
原核细胞往往小而简单,内部结构少;现代植物和动物的真核细胞比之复杂不少,具备许多执行特定功能的细胞器
他们希望找到其谱系可追溯至古生菌宿主或线粒体祖先的基因。线粒体祖先是变形菌门α-变形菌纲的成员。但他们还惊讶地发现了似乎来自多种其他细菌的基因。
加巴尔东和同事做出假设:真核生物的细胞祖先从多种合作伙伴处获得基因,这些伙伴可能曾被收编为内共生菌,之后又丢失了;也可能一直都是自由生活的细菌,通过水平基因转移将自己的一个或多个基因传递给宿主。无论以何种形式,真核“大跃进”都涉及两名以上合伙人。
加巴尔东在2021年的《微生物学年度评论》(Annual Review of Microbiology)中写道:“很明显,还有来自其他参与者的贡献。”
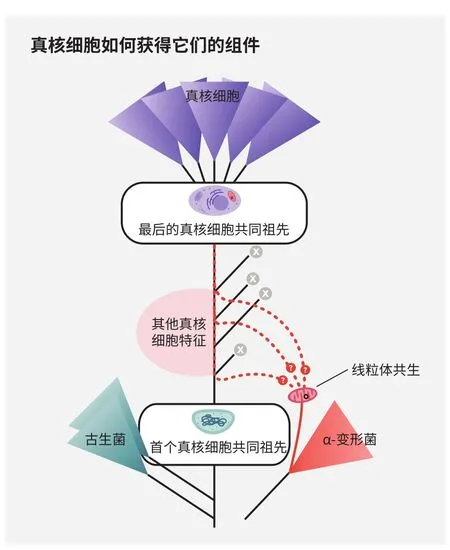
真核细胞起源于被称为古生菌的原始细胞,最终获得复杂特征,例如细胞器,但人们对这段往事的确切顺序所知甚少,许多被认为处于中间阶段的种类现已灭绝。学界仍在争论,因内共生而来的线粒体究竟是否出现于原核-真核转变的早期阶段—这决定了它究竟是演化的关键触发事件,还是后期改良因素
由于年深岁久,很难确切知晓那些古老外来基因的源头。不过亚利桑那州立大学的演化细胞生物学家约翰·麦卡琴(John McCutcheon)表示,有许多更晚出现、结构更松散的内共生体,它们的外源基因起源更易判断。通过类比研究,科学家有望借助它们揭示线粒体和第一批真核生物的演化过程。
细胞伴侣
在粉蚧(又名水蜡虫)的细胞里,生活着由两种细菌组成的内共生体,其中一种“内嵌”于另一种。细菌内共生体已经与粉蚧保持了大约1亿年的伙伴关系。(内共生体可制造水蜡虫无法从饮食中获取的必需氨基酸。)
麦卡琴和同事通过基因组分析发现,粉蚧的代谢途径简直就是一幅基因拼图画:源于虫子自身的基因,拼接上内共生体的核酸序列,或通过环境中其他微生物的水平转移而吸收的遗传片段。
麦卡琴等人表示,粉蚧细胞必须演化出一套运输系统。它能够在曾经独立的生物体之间来回输送蛋白质,允许粉蚧的细胞穿过两组内共生体膜,供“合伙人”使用。
差不多的情况也出现于宝琳虫属(Paulinella)细胞,一种类似变形虫的单细胞真核细胞。宝琳虫属细胞在数千万年前吞并得到内共生体,从而能够在无叶绿体的情况下利用太阳能。德国杜塞尔多夫大学的实验室负责人伊娃·诺瓦克(Eva Nowack)发现,宝琳虫属细胞基因组包含来自内共生体的基因以及通过水平基因转移获得的其他基因。
内共生体从宝琳虫属细胞处“进口”了400多种蛋白质,因此宝琳虫属细胞必须像粉蚧细胞那样演化出复杂的蛋白质运输系统。加拿大达尔豪西大学教授安德鲁·罗杰(Andrew Roger)表示,“这非常令人兴奋,因为它表明输送功能的演化并不像过去想象的那么困难”。

在大约1亿年的时间里,粉蚧及其内共生体已经演化出复杂的系统,可助基因产物往来穿梭,正如线粒体和它的宿主细胞那般
上述例子说明了内共生体与宿主整合的方式,也表明在真核生物演化早期,各种来源的水平基因转移可能非常频繁。
罗杰表示:“很多强有力的证据显示,真核生物中存在水平基因转移,因此真的没理由说它不可能发生于原核生物向真核生物转变的时期。事实上,几乎可以肯定它确实发生了。”
逐步演化假说
加拿大达尔豪西大学的比较基因组学家约翰·阿奇博尔德(John Archibald)解释道,古代宿主可能是逐步获得真核生物特征的,就像购物者把商品逐件加入购物车——它们通过水平基因转移或吞并一系列内共生体,逐渐发展完善出现代真核细胞的全部机制,其中一些新获得基因可能对宿主很有用。
若然,当远古宿主吞噬线粒体祖先时,它可能已经拥有了许多真核生物特征,包括一些细胞器,这意味着线粒体并非真核生物演化的主要驱动力,而是后期改良因素。

上图从左至右分别为阴道毛滴虫(Trichomonas vaginalis)、蓝氏贾第鞭毛虫(Giardia lamblia)和布氏锥虫(Trypanosoma brucei)。它们都被认为是原始真核生物。生物学家对它们以及其他罕见真核生物的基因组进行测序,意外发现一些“无主”基因—似乎既非源于原始真核宿主细胞,也不是来自其线粒体共生体。这意味着可能有其他“合伙”物种参与促成了真核生物起源
不过逐步演化假说也受到不少质疑。首先,前文介绍的这些现代内共生案例,粉蚧和宝琳虫属这类现代真核生物,能否等同于原核-真核转变时期的生物,能否真实反映当时的内共生情况?这是一个很大的疑问。
杜塞尔多夫大学的演化生物学家比尔·马丁(Bill Martin)表示:“那些案例似乎告诉我们,一旦有了真核细胞,就可轻易建立胞内共生体。但对于原-真核细胞来说,情况亦然吗?我们还不知道。”
在马丁看来,祖先们的内共生难度可能大得多,而这不符合真核细胞逐步演化的理论。逐步演化假说认为真核细胞的祖先在吞并线粒体以前就已非常接近真核细胞,能够轻松开展内共生;因此,若原-真核细胞如马丁所言,很难建立胞内共生体,这就意味着它们其实距离真正的真核细胞还很远,逐步演化假说自然站不住脚了。
事实上,有证据显示,关键的真核生物特征是一次性获得的。学习过细胞生物学的人都知道,所有真核生物都具备完全相同的一组细胞器:细胞核、核仁、核糖体、粗糙内质网和光滑内质网、高尔基体、细胞骨架、溶酶体以及中心粒。(植物和其他光合真核生物还具有叶绿体。)
霍华德休斯医学研究所珍妮莉亚研究园区的细胞生物学家詹妮弗·利平科特-施瓦茨(Jennifer Lippincott-Schwartz)表示,标准化的细胞器套件有力地自证——它们几乎都起源于同一时间,如若不然,不同真核生物应有不同细胞器组合。
一些生化证据也支持此观点。宿主古生菌和内共生体细菌属于生命之树的不同分支,使用不同分子构建膜结构。真核细胞器的膜结构并非完全古生菌式的,因此它们不大可能来自宿主。这个情况也意味着,古生菌宿主似乎是一种相对简单的细胞,因为它在内共生体细菌,即线粒体祖先来到后,才演化出其他细胞器。
那么关于前文提到的那些神秘外来基因,有什么说法吗?马丁认为还存在一种可能:所有外来基因或许都与线粒体祖先一起打包进入宿主,然后在15亿年间分散至许多细菌群体。它们的开枝散叶要归功于细菌善于交换基因的特点,却也迷惑、误导了后世学者,让他们以为多个合作伙伴为真核生物起源贡献基因。
马丁还补充道,假使逐步演化论正确,不同真核生物谱系就应当有不同基因集合,这种不同是根本性的,可测量的,但已证明此情况并不存在。
线粒体起源之争还将继续。根据罗杰的说法,“我们还很难找到能用于明确区分这些理论的数据”。不过若有进一步研究揭示一些只具备某组真核细胞器子集的生物,逐步演化假说就可得实证支持。另一方面,如果证据显示简单的古生菌细胞确能获得内共生体,那么“线粒体驱动真核生物形成”的理论将更具说服力。
资料来源 Knowable Magazine
