喀斯特地区珍贵树种黄枝油杉的种子萌发特性
2022-07-28江海都谢伟玲柴胜丰唐健民蒋运生秦惠珍
江海都, 谢伟玲, 柴胜丰*, 唐健民, 蒋运生, 秦惠珍, 韦 霄
( 1. 桂林理工大学 旅游与风景园林学院, 广西 桂林 541006; 2. 广西壮族自治区中国科学院 广西植物研究所, 广西 桂林 541006 )
种子植物完成受精发育成为种子,并在适宜的条件下萌发为新的幼苗,种子是其生物学特性赖以生存的基础,在植物繁殖育种、生物多样性保护、种群扩散中具有重要的功能(郑光华,2004;Borja et al., 2016)。种子萌发是植物生活史中的一个关键环节,是植物完成种群扩散、占领新界和种群再生极为关键的步骤(杨辉等,2006)。研究植物种子的萌发过程可阐明该物种如何适应生境以及环境因子如何调节种子萌发(Jozef et al., 2002)。种子萌发是植物对外界环境抵抗力最弱的阶段,受温度、光照、土壤水分等环境因子以及自身生理特性的影响(郑道君等,2016;刘雪松,2019),也是植物为适应环境、维持自身繁殖发展而形成的一种生物学特性,具有重要的生态学意义(Liu et al., 2013)。种子萌发与定居受胁迫成为许多物种濒危与分布受限制的关键因素。濒危红树植物红榄李()种子萌发对果皮、温度、盐度和光照等微环境的适应性狭窄是其处于濒危状态的主要原因之一(杨勇等,2016);海南龙血树()种子萌发对果皮、温度、光照强度等微环境的依赖性较强,原有生境破坏导致微环境的改变可能是其处于濒危状态的主要原因之一(郑道君等,2016);合柱金莲木()种子萌发速度慢,萌发不整齐,幼苗生长缓慢,且种子萌发对温度的适应范围狭窄,再加上林下光照不足限制种子萌发,这些是导致该物种自然更新困难的重要因素(柴胜丰等,2010);蛛网萼()和金丝李()种子萌发对温度和水分的要求较高,这与其濒危有很大关系(张丽芳等,2015;张俊杰等,2018)。研究濒危植物的种子萌发特性,探寻种子萌发的限制因素,可为其濒危原因的阐明及保护策略的制定提供依据。
黄枝油杉()为松科油杉属常绿大乔木,是我国宝贵的植物资源(何平,2005),主要分布于广西、湖南、贵州三省(区)交界处的喀斯特石山局部区域,其树形优美,树干笔直,雄伟挺拔,适宜在园林绿化中种植;其抗旱性强,是石灰岩山地造林的优良树种(黄立铨,1982);其木材坚硬,结构精细,纹理平直,是建筑、家具、水利等行业的优良用材树种(蒋柏生等,2008)。由于黄枝油杉幼龄植株生长缓慢,母树结实较少,自然更新能力弱(黄立铨,1982),再加上近年来滥砍滥伐十分严重,野生资源数量迅速下降,已列入《国家重点保护野生植物名录》(国家林业和草原局,农业农村部,2021)。笔者近年来对其野生种群的初步调查表明,在一些小种群,即使在结实大年,其结实植株数量和球果产量均较少,这可能与其濒危有很大关系;而在一些大种群,在结实大年有10%~20%的成年植株结实,且少数植株结实球果数达500个以上,但其种群幼苗数量仍极少,这一方面可能是球果中饱满种子数量少,另一方面生境中的温度、光照、水分、土壤等环境因子可能限制了种子到幼苗转化。因此,对黄枝油杉的种实性状及其种子萌发特性开展研究有利于阐明该物种濒危的原因。目前,关于黄枝油杉的研究主要集中在引物开发(石远婷等,2021)、群落特征(江海都等,2020)、遗传多样性(谢伟玲等,2016,2017)、光合特性(柴胜丰等,2015)、繁殖技术(蒋柏生等,2008)、化学成分(何道航等,2006)等方面,而关于其种实性状及种子萌发特性方面的研究则未见报道。为此,本文拟对黄枝油杉的种实性状和种子萌发特性进行研究,旨在了解其种子形态特征及结种率,探讨温度、光照、土壤含水量、基质、贮藏温度和时间、不同地理种源等因素对种子萌发的影响,进而分析该物种濒危的原因,为其被有效保护和利用提供参考。
1 材料与方法
1.1 供试材料
黄枝油杉球果于10月中旬采自广西桂林市恭城县三江乡(GC)和临桂区沉桥村(LG)两个野生种群,以及桂林市雁山区桂林植物园(YS)迁地保护种群(表1),球果带回实验室自然风干1周。所有鳞片裂开后,将种子拨出,并选择完整饱满的种子作为备用。

表 1 各样地种群概况Table 1 General profiles of sampling sites
1.2 球果及种子形态特征的测定
用于形态特征测定的球果和种子(图1)来自上述3个种群。用游标卡尺和电子天平测量20个球果的长度、直径和重量,并统计每个球果种子总数以及饱满种子数,计算球果结种率。每个球果选5颗饱满种子,测定种鳞长、宽,种子长、宽、厚以及种子重(含种翅)。电子天平测定100粒 × 8组种子重量,计算千粒重。采用烘干法测定种子含水量,取100粒种子在105 ℃下烘干至恒重,计算种子含水量,重复3次。

A. 球果; B. 种鳞; C. 种子。A. Cones; B. Pericarps; C. Seeds.图 1 黄枝油杉球果和种子形态Fig. 1 Cones and seeds morphology of Keteleeria calcarea
1.3 种子萌发试验
选用粒形饱满、粒径均匀的恭城县三江乡种群的种子(不同种源试验除外),在LRH-250-G型光照培养箱中进行试验。试验前种子用0.3% KMnO溶液消毒30 min,再用清水冲洗干净。在种子萌发试验中,用小塑料盒铺2 cm厚的基质,将种子播种在基质表层。播种后喷洒0.5%的多菌灵溶液消毒,此后每半个月喷洒一次,并适时补充水分。每个处理设3个重复,每个重复为50粒种子。种子开始萌发后,每4 d统计一次萌发数量,种子萌发以胚根长度达到种子长度1/2为标准,连续20 d没有种子萌发则试验结束。
1.3.1 温度对种子萌发的影响 培养条件:光照设置为3 000 lx,12 h·d,以沙土(1/2河沙+1/2黏质土壤,用孔径为2 mm的土壤筛过筛后等体积混合,基质经过高温消毒)为萌发基质,设置3个温度梯度:30、25、20 ℃,进行不同温度处理下种子萌发试验。
1.3.2 光照对种子萌发的影响 培养条件:温度25 ℃,不同光照条件分别为持续光照(3 000 lx,24 h·d)、周期性光照(3 000 lx,12 h·d)和持续黑暗(0 lx,24 h·d)。以沙土为萌发基质,研究不同光照处理对种子萌发的影响。
1.3.3 土壤含水量对种子萌发的影响 培养条件:温度25 ℃,光照3 000 lx,12 h·d,分别设置30%、25%、20%、15%和10% 5个不同土壤含水量处理,以沙土为萌发基质,测定不同土壤含水量对种子萌发的影响。在萌发过程中每4 d称量1次,以补充蒸发的水分。
1.3.4 不同基质对种子萌发的影响 培养条件:温度25 ℃,光照3 000 lx,12 h·d,分别以河沙、黏质土壤、沙土、泥炭土、珍珠岩和石山土为萌发基质,研究不同基质类型对种子萌发的影响。
1.3.5 贮藏温度和时间对种子萌发的影响 培养条件:温度25 ℃,光照3 000 lx,12 h·d,沙土为萌发基质。以在-20 ℃、4 ℃、室温(15 ~ 25 ℃)下分别贮藏1、3、6个月的种子为材料,进行萌发试验。-20 ℃条件下的种子置于塑料袋中密封保存,4 ℃条件下的种子置于牛皮纸袋中保存,室温条件下的种子置于纤维袋中保存。
1.3.6 不同地理种源种子的萌发率比较 培养条件:温度25 ℃,光照3 000 lx,12 h·d,沙土为萌发基质,选取GC、LG和YS种群的种子进行萌发试验,测定不同种源种子萌发率的差异。
1.4 数据分析
记录种子萌发开始时间、萌发种子总数及萌发持续时间。萌发开始时间表示从播种到第1粒种子萌发所需要的天数;萌发率为全部萌发种子占播种种子数的百分比;萌发持续时间为从种子萌发开始至萌发结束的时间。运用SPSS 25.0软件检验不同处理间的差异显著性,并进行多重比较(Duncan法),使用Origin 2015软件绘图。
2 结果与分析
2.1 球果及种子形态特征
黄枝油杉果熟期为10月中下旬,球果圆柱形,平均长度(13.90±1.43) cm,平均直径(3.41±0.28) cm;种鳞平均长度(2.80±0.25) cm,平均宽度(2.60±0.17) cm。种子浅棕色,光滑,具长翅[平均长(2.27±0.32) cm],种子平均长度(不含种翅)(1.55±0.15) cm,平均宽度(0.62±0.05) cm,平均厚度(0.46±0.04) cm,平均重量(含种翅)(0.21±0.04) g(表2),种子平均千粒重(214.81±14.76) g,风干种子平均含水量15.28%±1.66%。3个种群球果平均结种率7.45%±6.54%,恭城种群最高,平均可达14.89%±1.82%,临桂种群次之,雁山种群最低(2.58%±1.10%)(表2)。
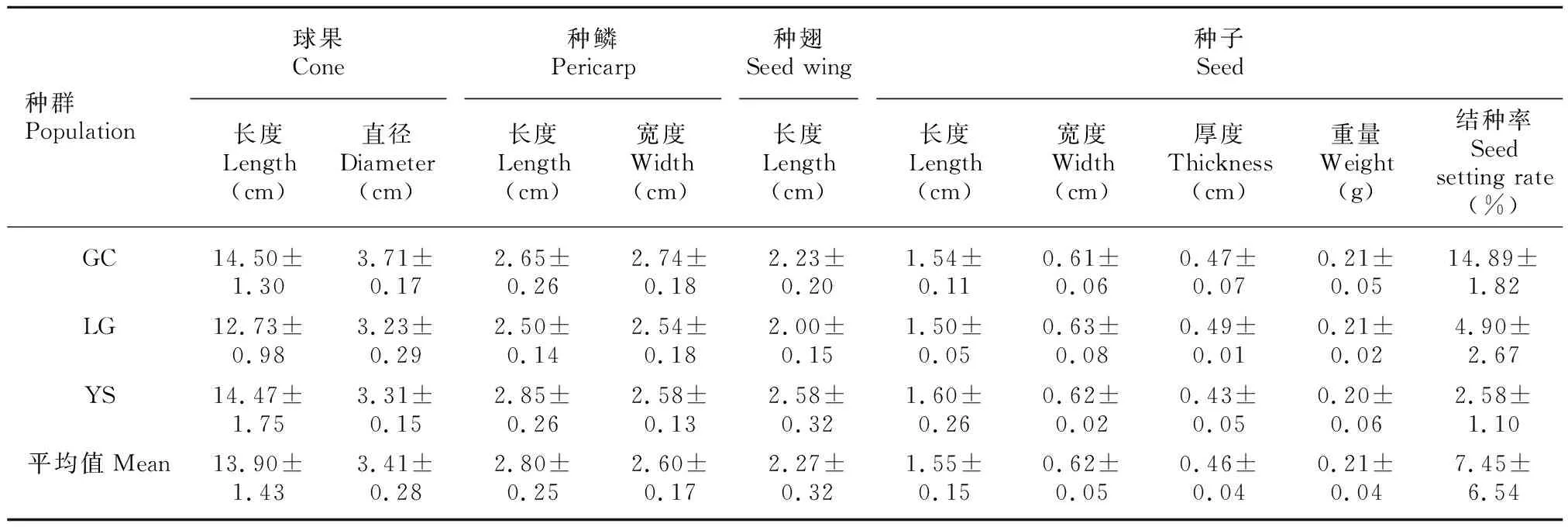
表 2 黄枝油杉球果和种子的形态特征Table 2 Morphological characteristics of cones and seeds of Keteleeria calcarea
2.2 温度对种子萌发的影响
不同温度处理下黄枝油杉种子萌发率有显著性差异(<0.05)。25 ℃条件下的萌发率显著高于20 ℃和30 ℃处理(<0.05)(图2)。30 ℃条件下,种子开始萌发时间和持续时间均较短。黄枝油杉种子萌发对温度有严格的要求,温度过高或过低,均不利于种子的萌发。

图中不同小写字母表示差异显著(P<0.05),下同。Different lowercase letters in the figure indicate significant differences (P<0.05), the same below.图 2 不同温度条件下黄枝油杉的种子萌发进程Fig. 2 Seed germination process of Keteleeria calcarea under different temperatures
2.3 光照条件对种子萌发的影响
不同光照条件下黄枝油杉种子萌发率有显著性差异(<0.05)。周期性光照(3 000 lx,12 h·d)条件下的种子萌发率显著高于持续光照(3 000 lx,24 h·d)和持续黑暗(0 lx,24 h·d)(<0.05)(图3)。在持续光照和持续黑暗下,种子萌发持续时间均较短。持续黑暗条件下,其幼苗呈现淡绿色或白色;持续光照条件下,幼苗长势较差,持续光照和持续黑暗条件处理不利于种子萌发和幼苗生长。

图 3 不同光照条件下黄枝油杉的种子萌发进程Fig. 3 Seed germination process of Keteleeria calcarea under different light conditions
2.4 土壤含水量对种子萌发的影响
不同土壤含水量下,黄枝油杉的种子萌发率没有显著性差异(>0.05)。各处理种子萌发开始时间均为28 d,萌发持续时间在65~88 d之间。在30%、25%和20% 3种土壤含水量条件下,其种子萌发速度要快于15%和10%土壤含水量(图4)。该物种种子萌发对土壤水分要求不严,能适应一定程度的土壤干旱条件。
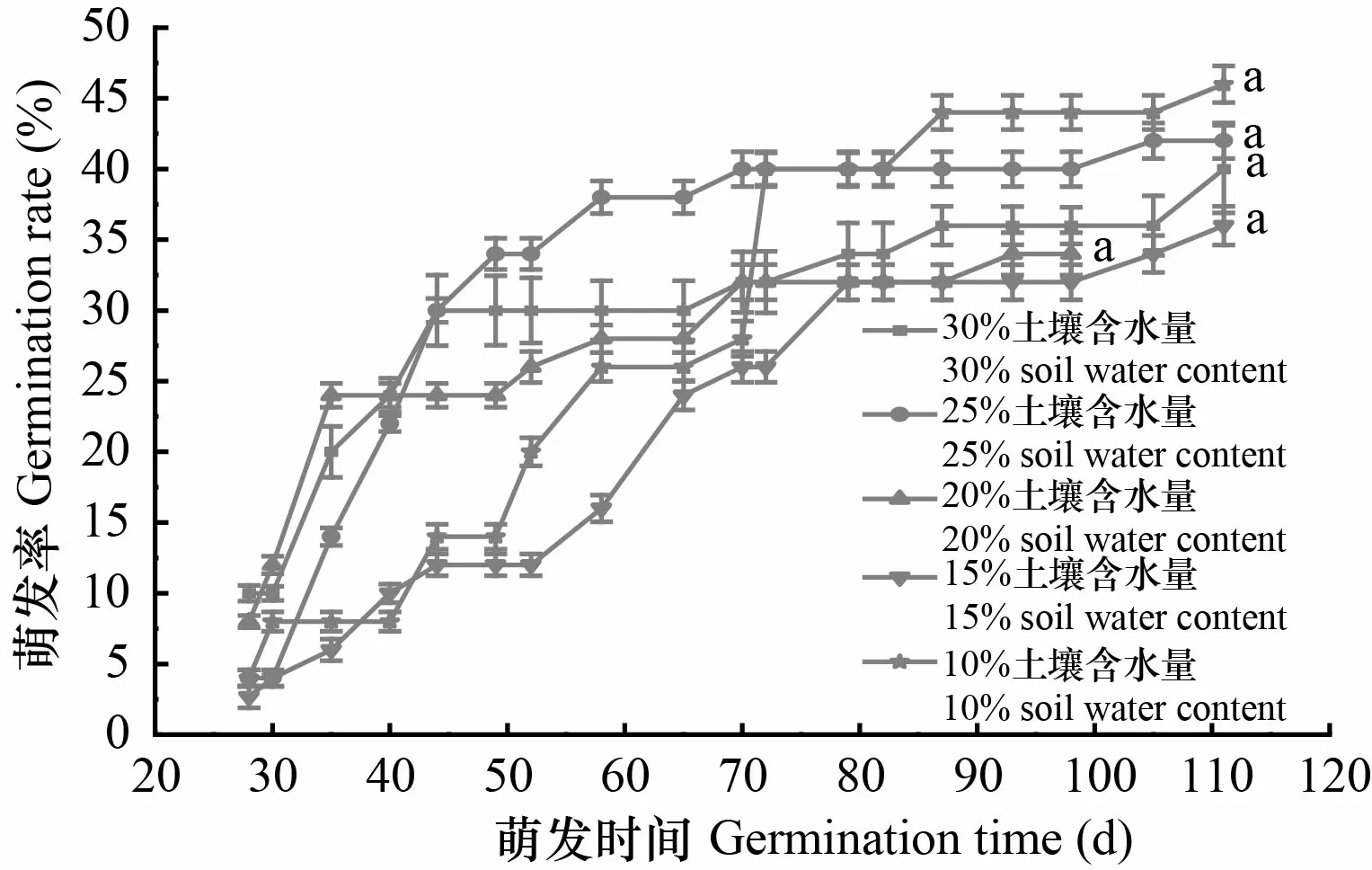
图 4 不同土壤含水量下黄枝油杉的种子萌发进程Fig. 4 Seed germination process of Keteleeria calcarea under different soil water contents
2.5 基质对种子萌发的影响
不同萌发基质对黄枝油杉种子萌发率有显著影响(<0.05)。珍珠岩和泥炭土的种子萌发率显著高于河沙、沙土、石山土和黏质土壤4个处理(<0.05);黏质土壤的种子萌发率最低,显著低于其他处理(<0.05)(图5)。在珍珠岩和泥炭土基质下,种子萌发持续时间明显长于其他处理。可见,黄枝油杉种子萌发对基质有严格的要求。

图 5 不同基质条件下黄枝油杉的种子萌发进程Fig. 5 Seed germination process of Keteleeria calcarea under different substrates
2.6 贮藏温度和时间对种子萌发的影响
不同贮藏温度和贮藏时间对黄枝油杉种子萌发率有显著影响(<0.05)。-20 ℃条件下贮藏种子的萌发率为0;4 ℃和室温(15~25 ℃)下贮藏1个月的种子萌发率分别为38.00%和36.00%,两者无显著性差异(>0.05);4 ℃和室温下贮藏3个月的种子萌发率显著降低(分别为20.00%和12.00%)(<0.05);4 ℃和室温下贮藏6个月的种子萌发率仅为6.00%和2.00%(图6)。可见,零上低温贮藏是更好的贮藏方式,但随着贮藏时间的延长,种子萌发率快速下降。

图 6 不同贮藏温度和时间下黄枝油杉的种子萌发进程Fig. 6 Seed germination process of Keteleeria calcarea under different storage temperatures and time
2.7 地理种源对种子萌发的影响
不同地理种源的黄枝油杉种子萌发率有显著性差异(<0.05)。恭城种群的种子萌发率显著高于临桂种群和雁山种群(<0.05)(图7)。3个地理种源的种子萌发开始时间和持续时间基本一致。

图 7 不同地理种源黄枝油杉的种子的萌发进程Fig. 7 Seed germination process of Keteleeria calcarea under different geographical provenances
3 讨论与结论
3.1 黄枝油杉球果及种子形态特征
种子是种子植物生活史的重要阶段,也是植物成功定居和更新的保障,能保证植物在时空上逃避不利环境(Bu et al., 2007)。种子大小影响植物后代的适合度,决定了幼苗建成和幼苗活力(Kidson & Westoby, 2000)。通常大粒种子比小粒种子能储藏更多的养分,有利于种子萌发和幼苗生长(傅家瑞,1985)。植物遗传因素、资源竞争和环境因子可以使植物产生不同大小的种子(Dech & Maun, 2006)。种子资源的投入有两种策略:一种是生产少量大种子以获得竞争优势,另一种是生产大量小种子以占据更多的生态位(李文良等,2008;呈福山等,2020)。本试验中,黄枝油杉种子平均长度1.55 cm(不含种翅),平均宽度0.62 cm,平均厚度0.46 cm,平均重量0.21 g,稍低于乔木树种的种子平均重量0.328 g(Silvertown, 1982),但与同属的云南油杉(种子重量为0.099 g)(陈美卿等,2010)和江南油杉(种子重量为0.107 g)(杨淼淼等,2020)相比,黄枝油杉的种子重较大,也高于近缘物种秦岭冷杉(千粒重为33.92 g)(赖江山等,2003)和元宝山冷杉(千粒重为16.96 g)(唐润琴等,2001)。恭城种群的球果结种率为14.89%,临桂种群为4.90%,远低于同属广布种江南油杉(49.35%)(何应会等,2017)。可见黄枝油杉更倾向于生产少量大种子的繁殖策略。恭城种群是目前已发现的最大种群,有较多中大径级个体,种群内个体数量超过2 000株,而临桂种群以中小径级个体为主,种群个体数量小于400株(江海都等,2020),种群大小和结构的差异可能是其结种率差异的重要原因。临桂种群仅有5株个体结实,结实数量从几十至二百不等,较低的种子产量是其种群更新障碍的重要原因。恭城种群有约10%的植株生产球果,多者球果数每株有500个以上,可见种子数量并不是该种群更新障碍的主要原因,种子向幼苗转化过程中环境因子的限制可能与其濒危有很大关系。
3.2 环境因子对黄枝油杉种子萌发的影响
温度是决定植物种子能否萌发的因素之一,过高或过低的温度都可能抑制种子萌发,或引起种子休眠或死亡(贺一鸣等,2008;闫兴富等,2016),这与一系列种子萌发参与的酶有关,酶需要在适宜的温度条件下发挥催化作用(张巧仙,2005)。适宜的温度是种子萌发所需的必要条件,掌握适宜种子萌发的温度对指导苗木生产实践具有重要意义(Maria et al., 2003)。本试验结果表明,黄枝油杉种子萌发率在25 ℃显著高于20 ℃和30 ℃,表明该物种种子萌发对温度的要求较为严格。温度过低,种子积温不足,不能在一定时间里积累到种子萌发所需的热量,种子相继出现活力降低、腐烂和发霉等现象;温度过高影响种子内部酶变性、膜的透性以及膜结合的活性导致萌发率降低,这与对江南油杉的研究结果相类似(王鸿彬等,2018a)。
不同物种种子萌发所需的光照条件不同(杨期和等,2001;Ali et al., 2021)。黄枝油杉种子在持续光照和持续黑暗条件下的萌发率显著低于周期性光照,且幼苗生长较差,说明持续光照或黑暗不利于黄枝油杉种子萌发及幼苗生长。在持续黑暗条件下,幼苗不能进行光合作用产生有机物质,原来储存的物质和能量随时间的推移被消耗,导致幼苗无法存活;持续光照可抑制种子萌发,同时也会引起干旱,使种子脱水失去活力,威胁幼苗生长。
土壤水分是影响种子萌发的重要因素。本试验中,黄枝油杉的种子萌发率在10%~30%土壤含水量范围内的差异不显著,但在土壤含水量高的情况下种子发芽较快,表明黄枝油杉对土壤水分没有严格要求,能耐石山干旱环境。这与江南油杉种子萌发对水分的需求明显不同,江南油杉在10%土壤含水量下不能萌发,15%和20%土壤含水量条件下显著低于25%和30%土壤含水量(王鸿彬等,2018b)。黄枝油杉耐干旱环境可能是其长期对喀斯特石山生境的适应。
不同的基质含有不同的营养物质、微生物种类,其紧密性也不同,从而影响种子的萌发、幼苗的生长和存活(Carolina et al., 2005)。本试验中泥炭土和珍珠岩的种子萌发率显著高于其他处理,而黏质土壤的种子萌发率最低。这可能是由于泥炭土和珍珠岩的质地疏松,透气性好,氧气充足,减少了霉变的发生,有利于黄枝油杉种子萌发;黏质土壤的粒间空隙小,透气性和透水性差,种子易霉变,萌发率低。
不同植物种子适宜的贮藏温度和时间存在较大的差异。我国亚热带地区的枫香,阴干后(含水量为10.54%)在室温、4 ℃、-20 ℃和-70 ℃条件下贮藏1 a,种子活力无明显差异,若贮藏2 a以上,则-70 ℃的贮藏效果最佳(裴云霞等,2020);而松科银杉种子活力的保持局限在一定的贮藏时间内,种子不耐过长时间的贮藏,宜随采随播,4 ℃低温苔藓包裹是适宜的保存方式(曹基武等,2010)。温度是影响种子新陈代谢的主要因子,种子在低温环境下呼吸作用很弱,物质和能量的消耗很少,因此胚部细胞可以长期保存其活力(浙江农业大学种子教研组,1987)。本试验中,黄枝油杉种子在室温、4 ℃和-20 ℃ 3种贮藏条件下,4 ℃贮藏相对较好,但贮藏3个月后种子萌发率仍会降低近一半,表明黄枝油杉的种子不耐贮藏,采回后宜及时播种。种子不能在-20 ℃贮藏可能与零下低温导致胞间和胞内水分结冰而产生组织损害有关。
不同地理种源的种子质量及活力因其母株地理位置和生境条件不同而不同,从而导致最终萌发率的差异(魏胜利等,2008;宿昊等,2021)。在本试验中,恭城种群的种子萌发率最高,这可能与该种群种子质量较好有关。该种群中的植株数量超过2 000株,是目前发现的最大野生种群(江海都等,2020),拥有许多大径级个体,结实母株数量和比例是最高的;而临桂种群以中小个体居多,结实植株数量少,种子质量较差。
本研究中的种子萌发试验在光照培养中进行,且种子均铺在长宽高不超过20 cm的小塑料盒内,虽然对种子和基质都进行了消毒处理,但种子萌发环境相对密闭,且萌发时间较长,仍有相当数量的种子发生霉变,导致本研究中黄枝油杉的种子萌发率相对较低。如能做好消毒处理或在通风透光条件好的环境下,将有助于该物种种子萌发率的提升。
3.3 黄枝油杉濒危原因分析及其对保育的启示
一方面,黄枝油杉依赖风媒传粉,且雄球花的花粉具有集中成熟和寿命短的特点,再加上散粉期在4月上旬,阴雨天气较多,若遭遇不利天气影响,将不利于其花粉的传播和胚珠受精,导致结实率低下,这可能是其结种率低的重要原因。在自然生境中,黄枝油杉球果于10月中下旬至11月上旬成熟,种鳞裂开后种子随风飘落到林下,但此时气温逐渐降低,桂林地区11月平均气温低于18 ℃,12月和1月的平均气温低于10 ℃,这将不利于黄枝油杉种子的萌发,再加上种子不耐贮藏,过冬后大部分种子将失去活力,这可能是限制种子向幼苗转化的关键因素。另一方面,黄枝油杉林下凋落物较多,光照不足,可能对种子萌发有一定影响,部分种子虽能萌发,但由于前期生长缓慢,无法与冠层下生长迅速的耐荫阔叶树抗衡,很容易被林中灌木或杂草淘汰。为了使种子落入土壤,同时提升其光照水平,以促进其生根发芽,可在黄枝油杉结果母树周边除草松土,为其自然更新创造条件;同时可对郁闭度较高的林分适当间伐,增强林下透光率,促进幼苗生长。此外,在黄枝油杉球果成熟时,可人工采集球果带回室内,在适宜的萌发条件下繁殖苗木,在合适时机对小种群进行回归引种,以满足种群更新的需要。
综上所述,黄枝油杉球果结种率较低,种子萌发对温度要求严格,且种子不耐贮藏,易失去活力,这与其濒危有很大关系。黄枝油杉野生种群幼苗数量少,种群更新困难,可采取人工抚育和回归引种的方式促进种群更新。
