过氧化氢酶对生长前期乌苏里貉生长性能、消化代谢、血清生化和免疫指标的影响
2022-07-25田建兴魏梦凡张海华
■刘 欣 刘 云 李 虎 田建兴 刘 日 魏梦凡 张海华*
(1.河北科技师范学院河北省特色动物种质资源挖掘与创新重点实验室,河北 秦皇岛 066600;2.辽宁威兰生物技术有限责任公司,辽宁 沈阳110000)
乌苏里貉作为我国三大主要毛皮动物之一,具有投资低、回本快的特点。据报道乌苏里貉在生长期特别是生长前期有必要加强幼貉钙、磷和蛋白质营养,以促进其骨骼和肌肉充分发育[1-2]。但是幼貉在快速发育的同时,体内产生很多的自由基,以及肠道系统的不完善,对饲粮中脂肪、蛋白质吸收不充分,易受到氧自由基的影响。氧自由基可对核酸、蛋白质等生物大分子进行攻击破坏,致使机体受到损伤[3-5]。
过氧化氢酶(catalase,CAT)是一种酶类清除剂,普遍存在于能呼吸的生物体内,主要存在于植物的叶绿体、线粒体、内质网以及动物的肝和红细胞中,其酶促活性为机体提供了抗氧化防御机理,与超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)共同组成了生物体内活性氧防御系统。前期研究表明,CAT 可以清除体内的自由基,对生长发育[6-8]、延缓衰老[9-10]、抗病[11-15]等均有很好的效果。目前,CAT 已作为饲料酶制剂应用于动物生产中,并对生长性能、营养物质消化率等方面有较好的促进作用[16-17],但在乌苏里貉上的应用报道较少。
本试验以乌苏里貉为研究对象,通过饲粮中添加不同水平的CAT,研究其对生长前期乌苏里貉生长性能、营养物质消化率、血清生化指标、血清免疫指标的影响,确定CAT 的添加水平,为CAT 在貉生长养殖过程中提供理论参考。
1 材料与方法
1.1 试验材料、试验动物及分组
在河北华夏新农科技股份公司昌黎莫各庄养殖场选取健康、体重相近的(60±5)d 的雄性乌苏里貉60 只,将其分成4 组(1~4 组),每组15 只。各组间初重(IBW)差异不显著(P>0.05)。本试验中使用的CAT由辽宁威兰生物技术有限公司提供。
1.2 试验设计及试验饲粮
采用单因素随机试验设计,分为对照组和试验组。对照组(1组)为基础饲粮,试验组是在基础饲粮中分别添加CAT 0.10%(2组)、0.15%(3组)、0.20%(4组)。
参考NRC(1982)[18]以及生产试验中饲喂效果较好的配方配制生长前期乌苏里貉的基础饲粮,其组成和营养水平见表1。

表1 基础饲粮组成及营养水平(风干基础)
1.3 饲养管理
试验开始前对食盆和笼舍进行详细消毒,按常规免疫程序对动物进行免疫。预试期7 d,正试期82 d,饲养试验于2020年7月5日-2020年10月5日。采用单笼饲养,每天7:30 和15:30 定时定量饲喂,让貉自由饮水,定期清洁笼舍。
1.4 消化代谢试验和血清制备
代谢试验于2020 年9 月3 日-2020 年9 月5 日进行,每组中分别选出采食与排便正常的貉各8 只,采用全收粪法收集粪尿。在每个收集桶内加入20 mL的10%的硫酸进行固氮,每天记录貉的采食量。3 d之后将全部的粪样和尿样分别收集。将收集的尿样混匀并过滤,放置在-20 ℃保存备用,粪样称重后按鲜重的5%喷洒10%的硫酸,65 ℃烘干至恒重,粉碎过40目筛,制成风干样本,保存待测。
生长前期试验结束后每组随机选取8只貉,在空腹时后肢静脉采血,用5 mL的促凝管收集血液,于离心机内4 000 r/min离心10 min,放置于-80 ℃保存,备用。
1.5 测定指标及方法
试验开始和结束分别称貉的空腹体重作为初重(IBW)和末重(FBW),计算平均日增重(ADG)、料重比(F/G)。统计试验过程中的总采食量,并计算相应的平均日采食量(ADFI)。代谢试验结束后,干物质(DM)、粪和尿中粗蛋白(CP)、粗脂肪(EE)均参照张丽英[19]方法测定,并计算营养物质消化率以及氮代谢相关指标。血清生化、免疫指标参照说明书通过VITAL-E全自动生化分析仪测定。
1.6 数据处理
试验数据用软件SPSS 24 进行单因素方差分析(one-way ANOVA,Duncan’s 和Dunnett’s T3 多重比较检验),试验结果差异显著用P<0.05来表示,差异极显著用P<0.01来表示,表中结果用“Mean±SD”的形式来表示。
2 结果与分析
2.1 CAT对生长前期乌苏里貉生长性能的影响(见表2)
由表2 可知,在试验结束时,2 组FBW 最大,2 组FBW、ADG 均显著高于1 组(P<0.05),F/G 显著低于1组(P<0.05);2 组、3 组、4 组的FBW、ADG 呈现下降趋势,F/G呈上升趋势,但没有显著差异(P>0.05)。
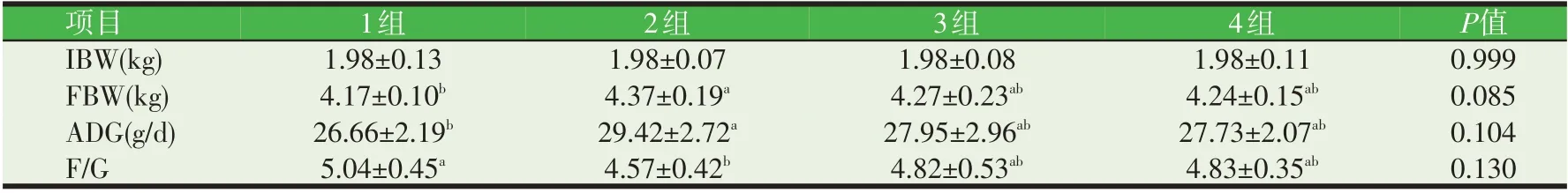
表2 CAT对生长前期乌苏里貉生长性能的影响
2.2 CAT 对生长前期乌苏里貉营养物质消化率的影响(见表3)
由表3 可知,各组间的ADFI、DM 消化率差异不显著(P>0.05);2 组的CP 消化率极显著高于1 组(P<0.01);EE 消化率方面,2 组与1 组、4 组之间差异极显著(P<0.01),3组和2组有显著差异(P<0.05)。

表3 CAT对生长前期乌苏里貉营养物质消化率的影响
2.3 CAT对生长前期乌苏里貉氮代谢的影响(见表4)
由表4可知,1组和2组在食入氮方面有显著差异(P<0.05);各组间的粪氮、尿氮、氮沉积均没有显著差异(P>0.05),但2、3、4组的尿氮含量均低于1组,氮沉积量均高于1组,其中2组氮沉积含量最高。

表4 CAT对生长前期乌苏里貉氮代谢的影响(g/d)
2.4 CAT 对生长前期乌苏里貉血清生化指标的影响(见表5)
由表5可知,1组的UREA显著高于试验2、3、4组(P<0.05);试验1、4 组的ALP 与2、3 组有极显著差异(P<0.01);1组的AST与其他几组均有极显著差异(P<0.01),试验3组与4组有极显著差异(P<0.01),与2组有显著差异(P<0.05);3 组血清中ALT 含量与1 组差异显著(P<0.05),2 组与其他组之间均没有显著性差异(P>0.05);血清中CHO、GLU、TG、TP含量各组间均没有显著差异(P>0.05)。

表5 CAT对生长前期乌苏里貉血清生化指标的影响
2.5 CAT 对生长前期乌苏里貉血清免疫指标的影响(见表6)
由表6 可知,3、4 组的IgA 含量与1 组相比差异极显著(P<0.01),3 组与2 组相比差异显著(P<5),1、2 组之间没有显著差异(P>0.05);2、4 组的含量极显著高于其他各组(P<0.01),且1 组与3组有极显著性差异(P<0.01);1 组的LPS 含量极显著高于2、3 组(P<0.01),2、3、4 组之间差异不显著(P>0.05);1 组的TNF-α含量与2、3、4 组之间均有极显著性差异(P<0.01),2 组和4 组之间有显著性差异(P<0.05)。

表6 CAT对生长前期乌苏里貉血清免疫指标的影响
3 讨论
3.1 CAT对生长前期乌苏里貉生长性能的影响
作为第三代酶制剂代表之一的CAT 在细胞中促进H2O2分解,使其不会进一步产生毒性很大的氧自由基,从而保护抗氧化系统,因此具有抑制细胞凋亡、促进机体发育的作用[20]。研究表明,饲粮中添加CAT对动物的生长性能有很好的效果[6,21]。方锐[22]通过在饲料中添加外源性CAT,提高了断奶仔猪的生长性能。研究发现CAT 的分布与动物的生长发育有关[7,23],在幼体时期CAT的活性较高,且CAT在肠道中只能发挥作用,而不能被肠道所吸收。CAT符合诱导契合学说,当活性氧浓度一定时,CAT浓度过高会使与H2O2反应所形成的中间复合物增多,其活性降低[24-25]。本试验结果显示随着CAT的浓度增加,貉体重呈现下降趋势,是因为在CAT增加的同时,氧化还原体系中所形成的中间产物也随之增多,导致CAT的活性降低,反而不利于机体的生长发育。本试验结果表明,在本试验条件下,添加0.1%的CAT提高了生长性能,降低料重比。
3.2 CAT对生长前期乌苏里貉营养物质消化率的影响
营养物质的消化率体现了动物对饲料的利用效率,同时也是评价饲粮好坏的重要指标之一。营养物质的消化吸收是动物调节生理功能的基础,营养物质消化率的提高对提高动物的生长性能有重要的意义。研究表明,添加一定剂量的CAT可以避免肠炎发生,提高了饲料的利用率,改善动物健康状态[26]。
本试验中试验组的ADFI比对照组低、DM消化率比对照组高,但差异不显著,说明在ADFI、DM消化率方面,饲粮中添加CAT 对生长前期乌苏里貉影响较小。当机体受到应激时,肠黏膜屏障易受到氧活性物质的攻击,导致肠黏膜屏障受损严重,而CAT 可以减少肠黏膜的活性氧,保护上皮细胞,减少细胞凋亡,抑制上皮细胞转录因子的活性[20,27]。本试验中CAT提高了EE 的消化率,主要是因为CAT 可以有效消除肠道内多余的氧自由基,保护肠道,改善肠黏膜屏障,防止脂类物质过氧化,从而对脂类物质可以更好地吸收利用与转化,这与方锐[22]的研究结果一致。随着CAT增加,对EE的吸收呈下降趋势。Lee等[12]发现通过增加CAT 活性可以减少蛋白羰基,从而减少对蛋白的氧化。本试验中,饲粮中添加0.1%的CAT 提高了CP 消化率,可能通过减少蛋白质的氧化来提高蛋白质的消化率。试验结果表明,在本试验条件下,CAT 能够显著提高生长前期乌苏里貉对EE 的消化吸收;添加0.1% CAT能极显著提高CP消化率。
3.3 CAT对生长前期乌苏里貉氮代谢的影响
氮代谢反映了貉对CP的利用率情况,机体对CP吸收的多少也影响着动物机体的健康状况。食入氮的多少取决于动物的采食量。机体内自由基可以消耗体内的糖类物质,从而降低机体对氮的吸收[5]。研究发现,CAT可以激活断奶仔猪消化道的消化酶活性,显著提高淀粉酶的活性[16]。而淀粉酶的增加会显著提高对氮的吸收利用,减少粪氮、尿氮的排放,从而减少氮对环境的污染[28]。而本试验中,饲粮中添加CAT能够减少生长前期貉对尿氮的排放,但是没有统计意义。随着CAT的水平上升,体内的氮沉积量呈先下降后上升的趋势,表明一定浓度的CAT限制了对氮的沉积。本试验结果显示,饲粮中添加CAT可以减少对环境的氮污染。
3.4 CAT对生长前期乌苏里貉血清生化指标的影响
UREA 通过动物机体中CP、氨基酸的代谢产生,主要经尿道排出体外,血清中UREA的含量与动物体内的氮沉积和CP 利用率的关系呈负相关[29]。血清ALT 与AST 是反映动物肝脏功能是否正常的常用指标,肝细胞的细胞质含有大量的ALT 和AST,胞浆中其浓度是血清中浓度的1 000~3 000 倍,在肝细胞损伤后,ALT 与AST 通过细胞膜释放进入血清,机体肝细胞受损程度越重,血清中ALT 与AST 含量越高[30]。研究发现自由基生成增多,导致肝组织脂质过氧化损伤[31-32]。本试验中,2、3、4组的UREA较于对照组减少了18.74%、22.57%、19.01%。试验1 组的AST 极显著高于其他几组,结果可能与CAT可以通过清除体内的氧自由基减轻氧自由基对肝细胞内细胞器结构的破坏有关。有研究表明ALP可以改变血管内稳态,对肾功能有一定调节作用[33]。王切等[34]发现,CAT 对肝缺血再灌注中肾的功能受损发挥着一定的保护作用。本试验中通过在饲粮中添加0.10%、0.15%的CAT 能极显著降低血清中ALP的含量,因此对维持正常肾功能有一定保护作用。在本试验条件下,饲粮中添加0.15%的CAT对肝功能、肾功能的保护作用最好。
3.5 CAT对生长前期乌苏里貉血清免疫指标的影响
IgA、IgG 作用在多种免疫机制中,是体液免疫的主要效应因子[35]。王洪军[36]研究表明,添加CAT 可以提高IgA、IgG含量,提高鸡血清新城疫抗体水平,提高肉鸡的免疫力。本试验中并未发现疾病的感染,这表明在本试验条件下,饲粮中添加0.15%和0.20% CAT可以极显著提高血清中IgA 含量;饲粮中添加0.10%和0.20% CAT 可以极显著增加血清中IgG 的含量。由于IgG在幼体抗感染免疫尤其是再次免疫应答中发挥重要作用,因此饲粮中添加0.1%和0.2% CAT 对促进生长前期的乌苏里貉的免疫指标有重要作用。LPS脂多糖,又称“内毒素”,是革兰氏阴性细菌细胞壁外壁的组成成分。LPS是构建肠道炎症较好的激活剂[37-39]。它很难从细胞壁脱落,当细菌死亡时它会通过溶解、破坏细胞来脱落,并通过作用于动物细胞等发挥其毒性。TNF-α是肿瘤坏死因子,可以促进T细胞产生各种炎症因子,进而促进炎症反应的发生。研究表明[40-43],它可促进炎症细胞因子的产生,引发炎症反应。李威等[26]在CAT对断奶仔猪的研究中发现,CAT可以显著降低过敏指数,显著提高抗过敏(炎症)功能,与本试验通过添加CAT可以减轻炎症反应,提高机体的免疫力结果一致。王慧[27]研究表明,添加适量的CAT较高脂组可以显著减少TNF-α含量,对降低LPS也有一定作用。本试验生长前期乌苏里貉的饲粮具有较高的脂肪含量,因此较对照组,其他几组的TNF-α、LPS含量均表现为降低。并且在本试验条件下,相较于浓度低的CAT 来说,CAT 浓度过高可能会导致炎症的发生,影响机体健康。本试验结果显示,饲粮中添加CAT 能够降低TNF-α、LPS 的含量,提高IgG,从而保护体内免疫系统。
4 结论
综上所述,饲粮中添加适量的CAT能提高生长前期乌苏里貉的生长性能、EE 消化率,改善血清指标,提高乌苏里貉机体免疫力。本试验条件下,在生长前期乌苏里貉饲料中添加0.1% CAT,貉的生长性能显著提高。
