野生欧洲李光合特性日变化及其与环境因子的相关性分析
2022-07-20曹云冰王亚铜潘佳琪耿文娟牛莹莹
曹云冰,陈 曦,王亚铜,潘佳琪,耿文娟,牛莹莹
(新疆农业大学 园艺学院,新疆 乌鲁木齐 830052)
光合作用是一系列复杂的生理生化过程,影响光合作用的因素主要分为内部因素和外部因素。影响光合作用内部因素主要有叶片的发育与结构(包括叶龄、叶位、叶片结构、气孔、光合色素等)[1]、光合产物的输出速率[2]。影响光合作用外部因素即环境因子主要有光照强度、温度、大气CO2浓度、水分、矿质元素等[3]。光照强度高于或低于植物光合作用所需时,植物净光合速率表现为下降趋势[4],而对于“双峰型”植物,适当的遮阴则有助于缓解植物光合午休现象[5]。CO2浓度在适宜范围内时,净光合速率与CO2浓度成正比,超出适宜范围,则净光合速率与CO2浓度成反比[6]。研究该植物的光合作用与环境因子相关性可以为该植物的移栽和品种选育提供理论依据[7]。
野生欧洲李(Prunus dometicaL.),俗名野酸梅,蔷薇科(Rosaceae),李属(Prunus)植物,为欧洲李野生种。野生欧洲李为中乔木或灌木,树高0.8 ~2.8 m,枝条无刺,暗紫褐色或灰褐色,嫩枝无毛,无棱。叶片椭圆形或宽卵形[8]。该种自1982 年首次被发现到现今,种群数量急剧缩减,目前野外现存415 株。原生分布地自然环境恶劣,树体营养不足,导致其花粉活力低,坐果率低,实生繁殖困难[9-10]。种群规模逐年缩小且种群分布严重破碎化,濒临灭绝[8]。近年来对野生欧洲李的研究多集中于分布环境[11]、植物组织培养[12]、亲缘关系[13]、表型多样性[14]、种子萌发特性及生理结构[15-16]等方面,在光合生理与环境相关性方面的研究未见报道。野生欧洲李作为一种濒危野生果树资源,目前对其研究相对较少,因此对其开展研究具有重要的栽培育种科研价值。目前野生欧洲李在我国处于野生状态,尚未有人工栽培的报道。
本试验以4 个样地、每个样地3 种树龄的野生欧洲李为研究对象,通过测定净光合速率、蒸腾速率、气孔导度等光合生理指标日变化及环境因子日变化,分析和探讨了野生欧洲李净光合速率的日变化特征,及其与环境因子的关系,并通过逐步多元回归与通径分析探讨环境因子与野生欧洲李净光合速率的相关性。希望本研究能揭示野生欧洲李光合生理特性和环境因子对净光合速率的影响规律,为野生欧洲李资源的保护开发及引种栽培提供理论依据和指导。
1 材料与方法
1.1 试验地概况
试验在新疆伊犁新源县阿勒玛勒镇、新疆乌鲁木齐新疆农业大学校园内进行(详情见表1)。
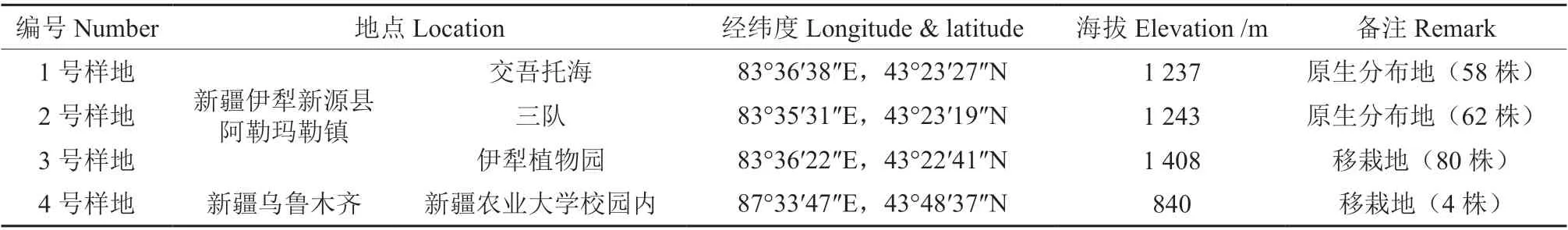
表1 试验地概况†Table 1 Overview of the test site
1.2 试验材料
选取1、2、3、4号样地中幼苗、幼树、成龄株(认定标准见表2)树势良好、无病虫害的野生欧洲李。

表2 认定标准Table 2 Identification standard
1.3 试验方法
于2020 年7 月随机选取晴朗无云的4 天,采用美国LI-COR 公司生产的Li-6400XT 便携式光合仪测量其生理生态指标。选取生长良好的植株,方向一致无遮挡,第5 叶位的新梢成熟健康叶,每株3 个重复,取平均值来分析相关参数。从10:00 测定到20:00,每隔2 h 测试1 次。测试指标有净光合速率(Pn, μmol·m-2·s-1)、蒸 腾 速 率(Tr, mmol·m-2·s-1)、胞 间CO2浓 度(Ci, μmol·mol-1)、 气 孔 导 度(Gs, mol·m-2·s-1)、光合有效辐射(PAR, μmol·m-2·s-1)、大 气CO2浓 度(Ca, μmol·mol-1)、 空 气 相 对 湿度(RH, %)。空气温度(Ta, ℃)。根据公式计算水分利用率(WUE, μmol·mmol-1)和气孔限制值(Ls, μmol·mol-1)。WUE=Pn/Tr,Ls=1-Ci/Ca。
1.3 数据处理
采用Microsoft Excel 2016 软件进行数据初步处理,使用SPSS 21.0 软件对数据进行统计分析,采用LSD 法进行显著性检验(P<0.05),并做线性回归方程。采用origin 2021 软件进行图表绘制。
2 结果与分析
2.1 四个样地环境因子日变化
如图1 所示,4 个样地的Ta日变化曲线为“单峰型”。4、3 号样地于14:00 达到全天的最大值,分别为33.2、31.3 ℃,1、2 号样地于12:00 达到全天的最大值,分别为31.78、38.05 ℃。日气温均值从大到小排列为2 号样地>4 号样地>3 号样地>1 号样地,分别为30.24、30.06、28.28、26.3 ℃。
如图1 所示,4 个样地的RH 日变化曲线均为“V 型”。3、4 号样地于14:00 达到全天的小值,分别为30%、26.30%;1、2 号样地于12:00 达到全天的最小值,分别为24.4%、25.1%。RH 变化日均值从大到小排列为1 号样地>2 号样地>3号样地>4 号样地,分别为58.65%、45.31%、44.41%、29.61%。表明原生分布地1、2 号样地的RH 要大于移栽地3、4 号样地。
如图1 所示,4 个样地的Ca日变化曲线呈现逐渐递减趋势。10:00 为全天的最大值,Ca变化日均值从大到小排列为4 号样>3 号样地>1 号样地>2 号样地,分别为414.824、408.715、405.137、403.359 μmol·mol-1。表明移栽地3、4 号样地的Ca均大于原生分布地1、2 号样地。
如图1 所示,4 个样地的PAR 日变化曲线为“单峰型”。PAR 变化日均值从大到小排列为4 号样地>3 号样地>2 号样地>1 号样地,在14:00 达到全天的最大值,分别为1 776.871、1 729.012、1 595.505、1 476.293 μmol·m-2·s-1。表明移栽地3、4 号样地的PAR 均大于原生分布地1、2 号样地。
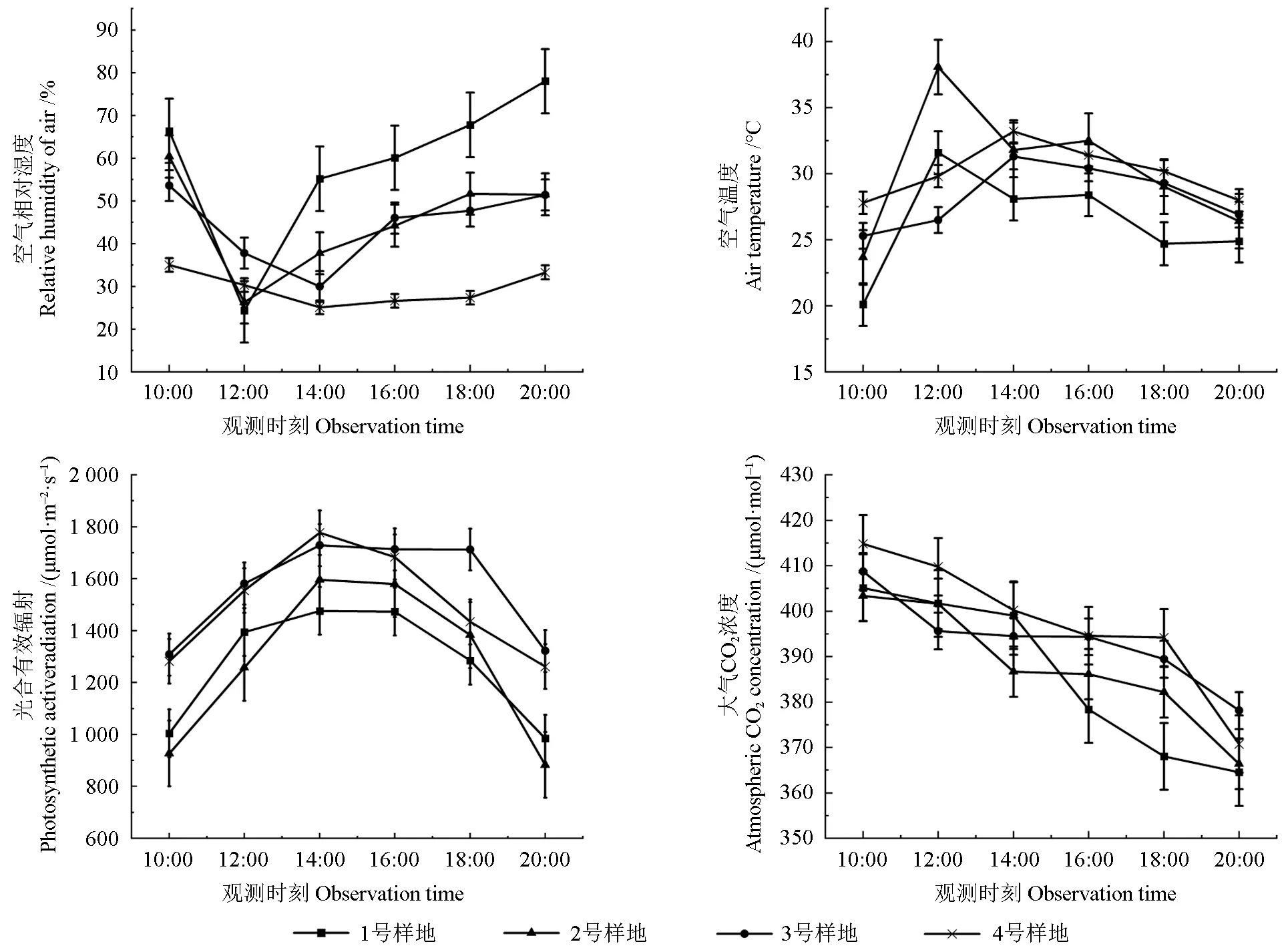
图1 环境因子日变化Fig.1Diurnalvariationofenvironmentalfactors
综上,野生欧洲李的原生分布地相较于移栽样地有温差大、空气相对湿度高、大气CO2浓度低、光合有效辐射不高的环境特点。
2.2 野生欧洲李光合作用日变化
2.2.1 不同树龄野生欧洲李净光合速率(Pn)的日变化
由图2 可知,4 个样地不同树龄的野生欧洲李Pn日变化曲线多为不对称“双峰型”。1 ~3 号样地的Pn日变化曲线为“双峰型”,并且存在明显的光合午休现象。第一个波峰出现在12:00,第二个波峰出现在18:00—16:00,第二个波峰明显小于第一个波峰。4 号样地的Pn日变化曲线为“单峰型”,主要是由于13:00 过后该植株处于遮阴状态,Pn急速下降,14:00 为全天最小值,仅为0.590 μmol·m-2·s-1,后续虽有所回升但仍不超过2.50 μmol·m-2·s-1。幼苗Pn日变化曲线中第二个波峰明显小于第一个,4 号样地表现更为明显,说明幼苗受光合午休的影响较大,并且较难恢复到原先的光合水平。4 号样地的幼树、幼苗Pn午间光合午休明显低于其余3个样地,波峰明显高于其余3 个样地,推测4 号样地午间高温高光强的环境对野生欧洲李光系统影响较大。
原生分布地1、2 号样地的Pn日变化均值大于移栽地3、4 号样地。4 号样地的日变化差值明显大于其余样地。野生欧洲李Pn地区日变化均值为2 号样地>1 号样地>3 号样地>4 号样地,分别为11.865、9.936、8.283、7.011 μmol·m-2·s-1。对比同一样地不同树龄的Pn发现,1、3、4 号样地不同树龄的野生欧洲李Pn日均值从大到小排列为幼树>成龄株>幼苗,2 号样地树龄与Pn日均值成正比。
2.2.2 野生欧洲李气孔导度(Gs)的日变化
由图2 可知,野生欧洲李Gs日变化曲线与Pn变化相似,都为“双峰型”,在12:00 达到第一个波峰,在16:00 或18:00 达到第二个波峰;4 号样地的幼苗野生欧洲李Gs日变化曲线与Pn相似,第二个波峰明显小于第一个,14:00 的4 号样地幼树、幼苗Gs明显低于其余3 个样地,两个波峰明显高于其余3 个样地。
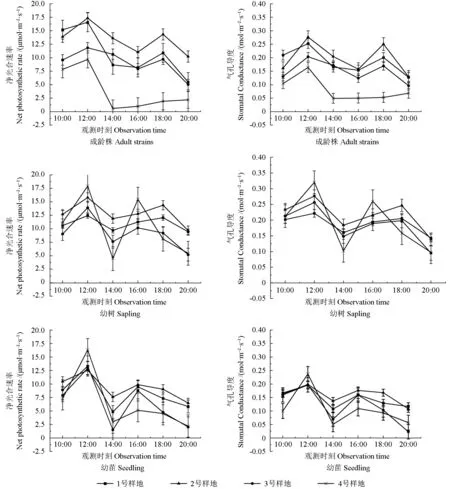
图2 不同样地不同树龄野生欧洲李净光合速率(Pn)、气孔导度(Gs)日变化Fig. 2 Diurnal variation of net photosynthetic rate (Pn), stomatal conductance (Gs) at different ages of P. dometica L.in different distribution areas
原生分布地1、2 号样地野生欧洲李Gs日变化均值大于移栽地3、4 号样地。4 个样地叶片Gs日变化均值从大到小排列为2 号样地>1 号样地>3 号样地>4 号样地,分别为0.190、0.172、0.150、0.127 mol·m-2·s-1。分别对比4 个样地不同树龄的Gs日变化均值发现,1、2、3 号样地的不同树龄Gs日变化均值由大到小排列为幼树>成龄株>幼苗,4 号样地的不同树龄Gs日变化均值由大到小排列为幼树>幼苗>成龄株。
2.2.3 野生欧洲李蒸腾速率(Tr)的日变化
由图3 可知,4 个样地不同树龄的野生欧洲李Tr日变化曲线多为“倒N 型”。1 ~3 号样地的Tr日变化曲线为“倒N 型”,呈“下降—上升—下降”趋势,12:00 或14:00 达到波谷,16:00 或18:00 达到波峰。4 号样地成龄株Tr日变化曲线为逐渐下降的曲线,10:00 为全天的最大值。2 号样地的成龄株16:00—18:00 的Tr明显大于其余3 个样地,这是由于其12:00 后处于遮阴状态,所以在午间仍维持较高的蒸腾速率。
原生分布地1、2 号样地的Tr日变化均值大于移栽地3、4 号样地。4 个样地的野生欧洲李Tr地区日变化均值为2 号样地>1 号样地>3 号样地>4 号 样 地,分 别 为5.790、5.376、4.943、3.940 mmol·m-2·s-1。对 比 同 一 样 地 不 同 树 龄 的Tr日均值发现,1、4 号样地不同树龄的Tr日均值从大到小排列为幼树>幼苗>成龄株;2、3 号样地不同树龄的Tr日均值从大到小排列为幼树>成龄株>幼苗。
2.2.4 野生欧洲李水分利用效率(WUE)的日变化
由图3 可知,野生欧洲李WUE 日变化曲线与Tr日变化曲线相反,多为“N 型”,呈“上升—下降—上升”趋势。“N 型”在12:00 或14:00 达到波峰。2 号样地成龄株、幼苗12:00—14:00 的WUE 明显大于其余3 个样地,4 号样地的成龄株WUE 日变化曲线与Pn日变化曲线相似。
图3 不同样地不同树龄野生欧洲李蒸腾速率(Tr)、水分利用率(WUE)日变化Fig. 3 Diurnal variation of transpiration rate (Tr), water use efficiency (WUE) at different ages of P. dometica L.in different distribution areas
原生分布地1、2 号样地野生欧洲李WUE 日变化均值大于移栽地3、4 号样地。4 个样地叶片WUE 日变化均值从大到小排列为2 号样地>1 号样地>3 号样地>4 号样地,分别为2.583、2.340、1.943、1.773 μmol·mmol-1。对比4 个样地不同树龄的WUE 日变化均值,发现2、3 号样地的不同树龄WUE 日变化均值由大到小排列为成龄株>幼树>幼苗,1 号样地的不同树龄WUE 日变化均值由大到小排列为成龄株>幼苗>幼树,4 号样地不同树龄WUE 日变化均值由大到小排列为幼树>幼苗>成龄株。
2.2.3 野生欧洲李胞间CO2 浓度(Ci)的日变化
由图4 可知,4 个样地不同树龄的野生欧洲李Ci日变化曲线多为“U 型”。“U 型”在10:00—12:00 的Ci急速下降,清晨叶片光合系统开始工作,Ci降低,18:00—20:00 又迅速上升,净光合速率迅速下降,Ci上升;4 号样地成龄株“单峰型”在14:00 达到全天最大值。
原生分布地与移栽地的Ci日变化均值无明显差异。4 个样地的野生欧洲李Ci日变化均值从大到小排列为4 号样地>1 号样地>3 号样地>2 号样地,分别为283.610、271.451、268.115、260.537 μmol·m-2。对比4 个样地不同树龄的Ci日变化均值,1、3 号样地不同树龄Ci日变化均值与树龄成反比,2 号样地的Ci日变化均值从大到小排序为幼树>幼苗>成龄株,与4 号样地相反。
2.2.4 野生欧洲李气孔限制值(Ls)的日变化
由图4 可知,Ls与Ci日变化呈相反的趋势,多为“单峰型”。在14:00、16:00、18:00 达到全天的最大值。1 号样地幼树、幼苗波峰明显高于其余几个样地。4 号样地成龄株Ls日变化曲线与Pn相似。
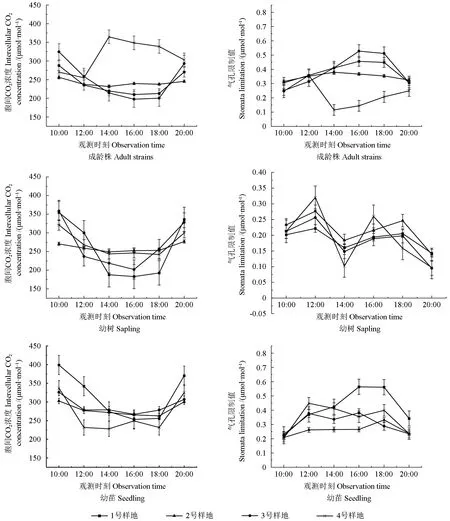
图4 不同样地不同树龄野生欧洲李胞间CO2(Ci)、气孔限制值(Ls)日变化Fig. 4 Diurnal variation of intercellular CO2 concentration (Ci), stomatal inhibition value (Ls) at different ages of P. dometica L. in different distribution area
4 个样地的Ls日变化均值差异不明显。4 个样地野生欧洲李的Ls日变化均值从大到小排列为1 号样地>3 号样地>2 号样地>4 号样地,分别为0.400、0.350、0.316、0.305 μmol·mol-1。对比4个样地不同树龄的Ls日变化均值,发现2、3 号样地的不同树龄Ls日变化均值与树龄成正比,1、4号样地的同树龄Ls日变化均值排列顺序为幼树>幼苗>成龄株。
2.3 野生欧洲李净光合速率Pn 与其生理生态因子的关系
2.3.1 简单相关性分析
影响光合作用的因素有很多,通过对野生欧洲李净光合速率Pn与其生理生态因子进行简单的相关性分析,结果(表2)发现Ci与树龄呈显著正相关关系,Pn与Ls、Ca呈显著正相关关系,与Tr、Gs、WUE 呈极显著正相关关系,与Ci呈极显著负相关关系。Tr与Gs、RH 呈极显著正相关关系,与Ls呈显著正相关关系。Ci与Ls、Gs、Ta极显著负相关关系,与WUE 呈显著负相关关系。Ls与Gs、WUE 呈显著正相关关系,与PAR 呈极显著正相关关系。Gs与WUE 呈及显著正相关关系,与Ca呈显著正相关关系。WUE 与Ta呈显著正相关。Ta与PAR 呈极显著正相关关系,与RH 呈极显著负相关关系。RH 与PAR 呈极显著正相关关系,与Ca呈极显著负相关关系。Ca与PAR 呈显著性正相关。
2.3.2 多元线性回归分析
多元逐步回归分析是多元统计分析的一种,使用多元逐步回归分析可以反映出野生欧洲李Pn与其生理生态因子间的相关性,对不同样地、不同树龄野生欧洲李的Pn进行正态检验,根据Kolmogorov-Smirnov 分析结果sig.=0.200 >0.05,即Pn服从正态分布,具有线性关系,将生理生态 因 子Tr、Ci、Ls、Gs、WUE、Ta、RH、Ca、PAR 用X1、X2、X3、X4、X5、X6、X7、X8、X9表示,对野生欧洲李净光合速率Pn(Y)及各影响因子(X1-X9)分别进行逐步多元回归分析,得到最优回归方程为:Y=-2.826+57.363X4+0.027X8-0.014X2-0.003X9。
(R2=0.904,复相关系数R=0.951,标准估计误差S=1.263,F=178.796,P<0.05)
对野生欧洲李净光合速率(Y)回归方程进行方差分析和偏相关分析得知因变量和自变量之间均达到极显著或显著水平,表明该线性关系成立。从表3 的偏相关结果可知各因子与Pn无显著相关,影响野生欧洲李叶片净光合速率Pn变化的主要因子为Gs,并且Gs与Pn为正相关关系。偏相关分析结果与线性回归方程相吻合。
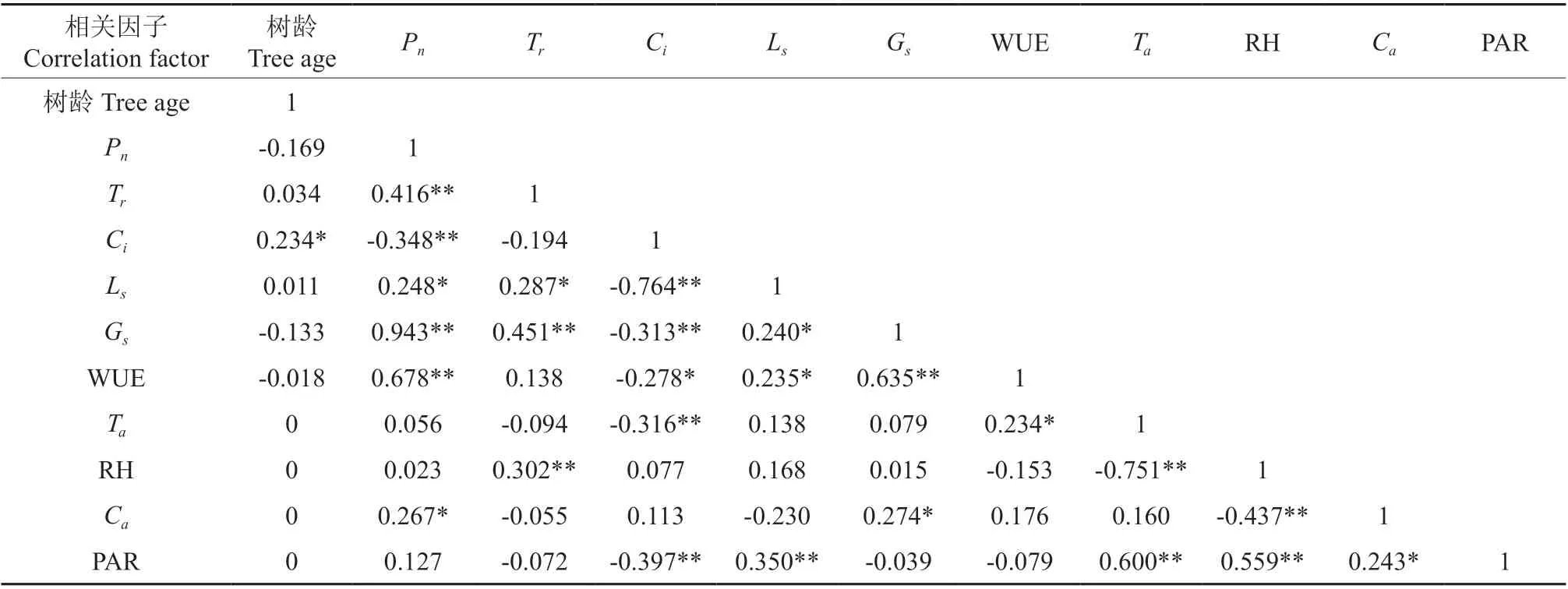
表3 野生欧洲李净光合速率Pn与生理生态因子间的关系矩阵†Table 3 Relationship matrix between net photosynthetic rate Pn and ecophysiological factors of P. dometica L.

表4 野生欧洲李净光合速率(Pn)与各因子的偏相关分析Table 4 Partial correlation analysis between net photosynthetic rate (Pn) and various factors of P. dometica L.
2.3.3 通径分析
通径分析能反映出生理生态因子对野生欧洲李Pn的影响及其复杂的相互作用关系(表5)。从直接作用绝对值从大到小看,各生理生态因子对野生欧洲李Pn的影响顺序Gs>Ci>Ca>PAR。Gs、PAR 直接通径系数为正值,证明其对野生欧洲李Pn有促进作用,Ci、Ca直接通径系数为负数,说明其对野生欧洲李Pn有抑制作用,通过分析各因子间的间接通径系数发现,Ci是通过Gs对野生欧洲李Pn的间接作用最大的生理因子。Ca是通过Gs对野生欧洲李Pn的间接作用最大的生态因子。
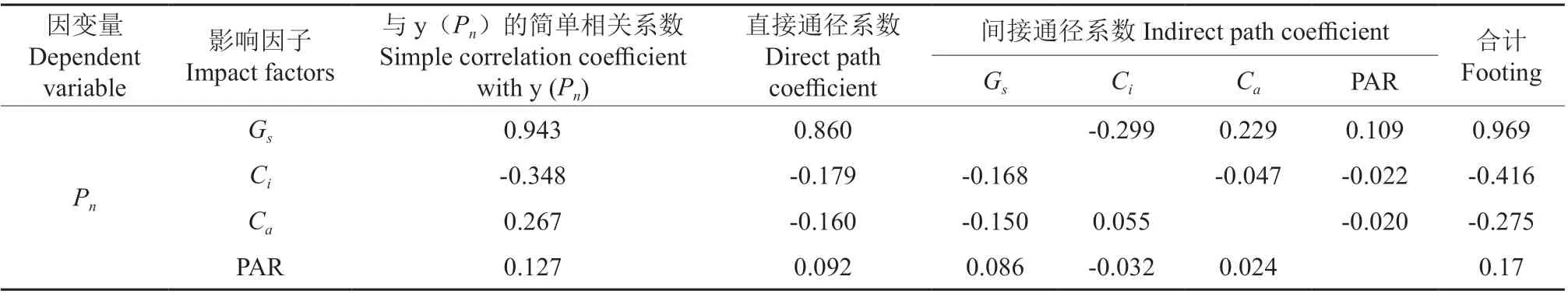
表5 野生欧洲李净光合速率(Pn)与其主要影响因子间的通径分析Table 5 Path analysis between net photosynthetic rate (Pn) and its main influencing factors of P. dometica L.
3 讨 论
3.1 野生欧洲李环境因子分析
野生欧洲李原生分布地1、2 号样地的环境特征为高RH,日温差大,PAR 不高等特点,移栽地3、4 号样地有RH 低,PAR 高,Ca高的环境特点。通过对不同树龄野生欧洲李样地环境因子进行简单相关性分析发现,Ta与RH 显著负相关,与PAR 呈正相关,与Ca呈负相关。随着时间的变化,Ta、RH、PAR、Ca也产生相应的变化。
3.2 野生欧洲李光合特性分析
植物光合作用是一个复杂的生理生化过程,衡量光合作用强弱的指标是净光合速率[17]。本研究发现,野生欧洲李的Pn日变化曲线为“双峰型”,存在较为明显的光合午休现象。光合午休现象广泛存在于山椒子[18]、葡萄[18]、酥李[19]等植物上。产生光合午休的原因有两种,一种是气孔因素,即中午高温,水分蒸发快,气孔部分关闭,Gs、Ci下降,从而导致午间光合速率下降[20]。另一种是非气孔因素,即中午光强、温度过高以及干旱胁迫导致Rubisco 酶活性降低,损伤细胞器,关闭气孔,Gs下降,Ci上升从而导致午间光合作用受抑制[21]。本研究结果表明,在午间,除4 号样地成龄株野生欧洲李外,其余样地、树龄的野生欧洲李Pn处于波谷即光合午休时,Ci、Gs同时下降,说明野生欧洲李光合午休为气孔因素;4 号样地成龄株野生欧洲李Ci随Pn的下降而上升,说明其光合午休为非气孔因素。
气孔是植物叶片与外界环境交换气体的主要通道。气孔导度受多方面因子的影响,对比野生欧洲李原生分布地以及移栽地的环境因子,发现1、2 号样地的Ca、PAR 日变化均值均大于3、4 号样地,1、2 号样地的RH 日变化均值小于3、4 号样地。1、2 号样地的Gs日变化均值大于3、4 号样地。除4 号样地成龄株野生欧洲李外,其余样地、树龄的野生欧洲李Gs日变化进程与Pn相似,均为“双峰型”,其峰值在12:00 和18:00 或16:00 出现,峰谷在14:00 或16:00 出现,可能是由于午间光强和温度过高导致气孔关闭,Gs降低,Ci降低从而引起WUE 下降。
通过对比4 个样地的成龄株、幼树、幼苗野生欧洲李的各项生理生态指标发现,随着树龄的增加,野生欧洲李的Pn与Gs均呈先增大再减小的趋势,Pn与野生欧洲李树龄呈负相关关系。原生分布地1、2 号样地的Pn与Gs日均值均大于移栽地的3、4 号样地,这与王婉妮[22]的研究结果相同,随着干旱程度的加深,葡萄砧木的Pn与Gs呈逐渐下降的趋势。
3.3 野生欧洲李净光合速率与环境因子的相关性分析
野生欧洲李Pn及其生理生态因子进行简单相关性分析发现,野生欧洲李Pn的变化与多个因子具有相关性,是受多个因子共同调控的复杂生理过程[23]。通过偏相关分析发现,野生欧洲李Gs与Pn呈正相关关系。多元线性回归分析和通径分析结果表明,影响野生欧洲李Pn变化的主要生理生态因子有Gs、Ci、Ca、PAR。本研究得出Gs是野生欧洲李Pn的主要正相关因子,Ca、PAR 与Pn呈正相关关系,Ci与Pn呈负相关关系。此外通径分析可以看出,Gs无论是直接正效应还是间接效应都是最大的,这表明在野生欧洲李光合作用过程中Gs为主要影响因子,而Ci与Ca通过其相互作用共同影响其Pn。对野生欧洲李Pn影响最大的生理因子为Gs,且Ca、PAR 对野生欧洲李Pn均具有积极作用,Ci对野生欧洲李Pn具有消极作用。
4 结 论
本试验通过研究野生欧洲李4 个样地、不同树龄Pn及其生理生态因子的日变化进程及其相关性,发现野生欧洲李的Pn日变化曲线为“双峰型”,光合午休因素为气孔限制性因素;野生欧洲李Pn与树龄呈负相关关系,与生理因子Gs正相关,与Ci负相关,与环境因子Ca、PAR 正相关。
通过多元线性回归分析,得出各生理生态因子与野生欧洲李Pn的线性回归方程Y=-2.826+57.363X4+0.027X8-0.014X2-0.003X9(R2=0.904,复相关系数R=0.951,标准估计误差S=1.263F=178.796P<0.05),影响野生欧洲李Pn变化的主要影响因子为Gs。为了保证野生欧洲李移栽的成活率,建议在移栽后增施二氧化碳气肥,并且在午间适当遮阴来减轻因外界高温高光强引起的气孔关闭现象。本研究探究了野生欧洲李不同树龄原生分布地与移栽地的光合特性日变化,虽然能反映野生欧洲李光合特性与环境因子的相关性,但要综合考虑野生欧洲李的环境适应性则需结合叶绿素荧光参数、生理生化指标、叶片形态结构结构、土壤相对含水量、土壤温度等指标进行综合评判。此外,本研究基于前期研究通过对比4 个样地的野生欧洲李成龄株、幼树、幼苗的净光合速率发现1、3、4 号样地的野生欧洲李存在早衰现象。因此,利用基因组学和分子手段挖掘早衰欧洲李中调控衰老的基因 并利用转基因方法确定对应基因是下一步的研究方向。
