羔羊断奶应激期饲喂方式对其生长性能和血清生化指标的影响
2022-07-11史晨迪赵晓雅田沛知陈佳欣段春辉张英杰刘月琴纪守坤
史晨迪 赵晓雅 田沛知 陈佳欣 段春辉 张英杰 刘月琴 纪守坤 严 慧
(河北农业大学动物科技学院,保定 071000)
羔羊成活率及生长发育决定了养殖经济效益。断奶是羔羊生长过程中的关键阶段,母子分离、饲粮转换、社群重组及环境变化等均会对羔羊造成应激,同时羔羊断奶时消化系统发育尚不完善,抵抗力低,发病率高,断奶阶段饲养管理对羔羊成活率及后期生长发育影响较大,断奶阶段是生产中格外关注的重要阶段[1-2]。
羔羊断奶前以母乳为主,皱胃较发达,瘤胃发育尚不完全,研究表明液体饲料长期饲喂会延滞羔羊瘤胃发育,植物饲料有助于刺激胃肠道发育,尤其是粗饲料能够刺激胃肠道发育,维持反刍动物瘤胃正常功能和机体健康[3]。羔羊断奶过渡期内,饲粮结构和组成对消化道的生长发育有重要作用,而全混合日粮(TMR)是将干草、青绿饲料或其他农副产品等粗饲料、精饲料、矿物质及维生素均匀混合调制而成的饲粮,具有低成本、营养均衡、适口性好、饲料利用率高和维持瘤胃内环境稳定等优点,能够满足羔羊生长所需要的营养物质,促进其生长发育,稳定渡过各个应激期。目前养殖场对于羔羊在断奶过渡期饲喂的饲粮各有不同,没有统一的饲粮过渡模式,主要包括代乳品、羔羊料+粗饲料、TMR粉料和羔羊料+TMR粉料等,羔羊腹泻及死亡率较高。在饲料端全面禁抗的背景下,如何有效缓解羔羊断奶应激已成为急需解决的生产问题。因此,本试验基于生产中主要的过渡饲料,分析比较不同饲喂方式对羔羊健康水平的影响,旨在为有效缓解断奶应激及促进羔羊发育提供理论基础。
1 材料与方法
1.1 试验时间与地点
本试验于2020年11月23日至2020年12月18日在河北省张家口市兰海牧业进行。
1.2 试验设计与饲养管理
本试验采用单因素试验设计,选取健康、体重相近的50日龄湖羊羔羊48只(公母各1/2),羔羊在50~60日龄随母哺乳,并补饲羔羊料(购自某饲料公司)。59日龄时(记为第0天),根据羔羊体重[(16.74±2.54) kg]、性别随机分为3组:试验1组(随母哺乳,并补饲羔羊料)、试验2组(60日龄断奶,并饲喂TMR粉料)、试验3组(60日龄断奶,并饲喂50% TMR粉料+50%羔羊料),每组16只。试验期16 d。全混合日粮组成及其与羔羊料营养水平、母乳营养水平分别见表1、表2。
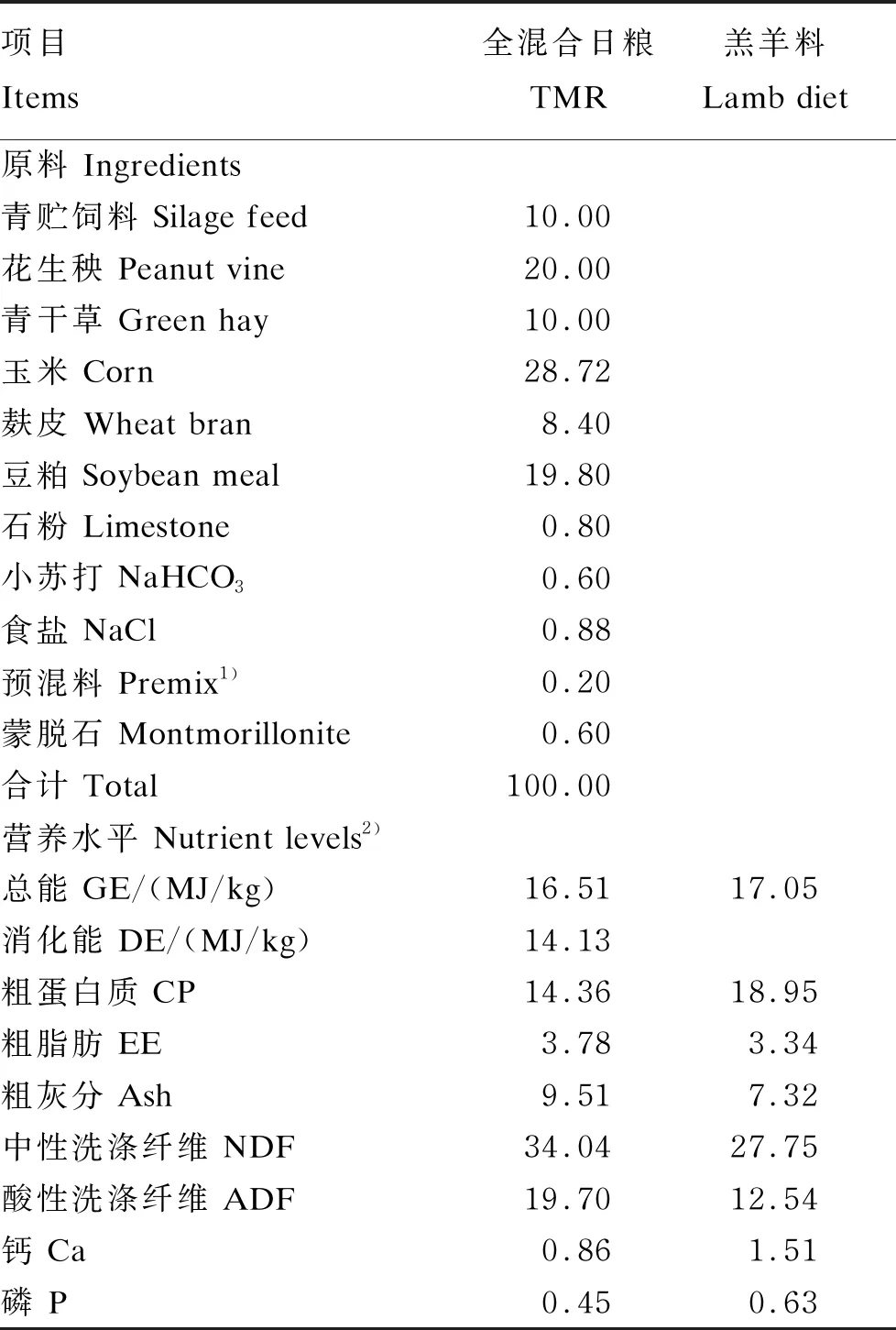
表1 全混合日粮组成及其与羔羊料
试验期间,每周对羊舍地面及四壁进行消毒处理,每天于09:00和16:00各饲喂1次,羔羊自由采食和饮水。
1.3 样品采集
所有羔羊在试验开始及结束时称重,在第0、3、9和15天于晨饲前颈静脉采集血液,离心机3 000×g离心10 min,分离上清液,-20 ℃保存备用。
1.4 测定指标及方法
1.4.1 生长性能
所有羔羊在第0和15天于晨饲前称重并记录,试验2组和3组记录每天饲料投喂量和剩料量。试验1组每天测定羔羊吃奶量(哺乳前后称重),并取奶样进行营养成分测定,同时记录羔羊料的投料量和剩料量。计算平均日增重(ADG)和干物质采食量(DMI)。
ADG=(试验羊末重-试验羊初始重)/试验天数;DMI=(每日投料量-次日剩料量)×干物质含量。
1.4.2 血清生化指标
采用酶联免疫吸附测定(ELISA)法测定血清皮质醇(COR)(检出限:1~400 ng/mL)、促肾上腺皮质激素(ACTH)(检出限:2~600 ng/L)、免疫球蛋白A(IgA)(检出限:0.1~40.0 mg/mL)、免疫球蛋白M(IgM)(检出限:0.2~60.0 mg/mL)、免疫球蛋白G(IgG)(检出限:0.5~100.0 mg/mL)、白细胞介素-1β(IL-1β)(检出限:0.5~200.0 ng/L)、肿瘤坏死因子-α(TNF-α)(检出限:3~900 ng/L)、干扰素-γ(IFN-γ)(检出限:2~600 ng/L)和D-乳酸(D-Lac)(检出限:20~6 000 ng/mL)含量。采用微板法测定血清乳酸脱氢酶(LDH)、谷丙转氨酶(ALT)、谷草转氨酶(AST)活性。采用比色法测定血清肌酸激酶(CK)活性。采用微量酶标法测定血清碱性磷酸酶(ALP)活性(检出限:0~60 金氏单位/dL)。采用重点显色法测定血清内毒素(ET)含量(检出限:0.1~1.0 EU/mL)。采用速率法测定血清二胺氧化酶(DAO)活性(检出限:0~100 U/L)。试剂盒均购自南京建成生物工程研究所。
1.5 统计分析
所有试验数据均用Excel 2010进行初步整理后,采用SPSS 21.0软件对数据进行单因素方差分析,采用Duncan氏法进行多重比较,结果均以“平均值±标准差”表示,显著水平为P<0.05。
2 结 果
2.1 断奶应激期内不同饲喂方式对羔羊生长性能的影响
由表3可知,在第0和15天,3组羔羊BW差异不显著(P>0.05)。在第0~15天,试验1和3组羔羊ADG显著高于试验2组(P<0.05)。在第0~15天,试验1和2组羔羊DMI显著低于试验3组(P<0.05)。
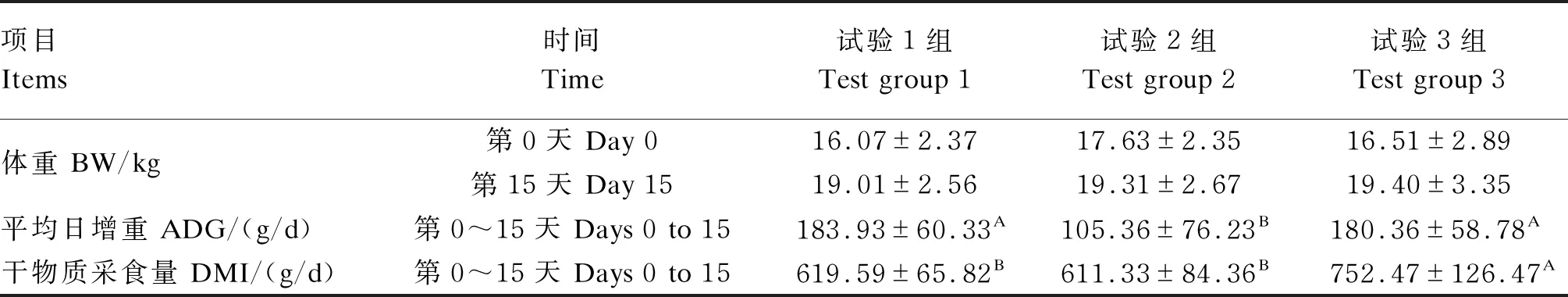
表3 断奶应激期内不同饲喂方式对羔羊生长性能的影响
2.2 羔羊断奶应激期内不同饲喂方式对血清中COR和ACTH含量的影响
由表4可知,不同时间组间点血清中COR和ACTH含量分析结果:在第3天,试验2组血清中COR含量显著高于试验1组(P<0.05),与试验3组差异不显著(P>0.05);在第0、9和15天,3组间血清COR含量差异不显著(P>0.05)。在第0、3、9和15天,3组间血清中ACTH含量差异不显著(P>0.05)。

表4 羔羊断奶应激期内不同饲喂方式对血清COR和ACTH含量的影响
同组在不同时间点血清COR含量的分析结果:在试验1组,不同时间点间血清中COR含量无显著差异(P>0.05)。在试验2组,第3天血清中COR含量显著高于第9、15天(P<0.05),与第0天差异不显著(P>0.05);第0天显著高于第15天(P<0.05),与第9天差异不显著(P>0.05)。在试验3组,第3天血清中COR含量显著高于第15天(P<0.05),第0天分别与第3、9、15天差异不显著(P>0.05),第3天与第9天差异不显著(P>0.05),第9天与第15天差异不显著(P>0.05)。
同组在不同时间点血清ACTH含量的分析结果:在试验1组和3组,血清中ACTH含量在不同时间点间差异不显著(P>0.05)。在试验2组,第3天血清中ACTH含量显著高于第9、15天(P<0.05),与第0天差异不显著(P>0.05);第0天显著高于第15天(P<0.05),与第9天差异不显著(P>0.05)。
2.3 羔羊断奶应激期内不同饲喂方式对血清中酶活性的影响
由表5可知,不同时间点组间血清中酶活性的分析结果:在第0、3和9天,3组间血清中LDH活性差异不显著(P>0.05);在第15天,试验1组血清LDH活性显著高于试验3组(P<0.05),与试验2组差异不显著(P>0.05)。在第0、9和15天,3组间血清中CK活性差异不显著(P>0.05);在第3天,试验2、3组血清CK活性显著高于试验1组(P<0.05)。在第0、3和9天,3组间血清中ALP活性差异不显著(P>0.05);在第15天,试验1组血清ALP活性显著高于试验2组(P<0.05),与试验3组差异不显著(P>0.05)。在第0、3、9和15天,3组间血清中ALT活性差异不显著(P>0.05)。在第3和15天,试验2组血清中AST活性显著高于试验1组(P<0.05),与试验3组差异不显著(P>0.05);在第0和9天,3组间血清AST活性差异不显著(P>0.05)。
同组在不同时间点血清中酶活性的分析结果:在试验1组,各时间点间血清中LDH、CK、ALT和AST活性无显著差异(P>0.05);血清中ALP活性在第15天显著高于第0和3天(P<0.05),与第9天无显著差异(P>0.05)。在试验2组,各时间点间血清中LDH、CK、ALP和ALT活性无显著差异(P>0.05);血清中AST活性在第15天显著高于第3天(P<0.05),与第0和9天无显著差异(P>0.05)。在试验3组,各时间点间血清中LDH、CK、ALT和AST活性无显著差异(P>0.05);血清中ALP活性在第9天显著高于第3天(P<0.05),与第0和15天无显著差异(P>0.05)。
2.4 羔羊断奶应激期内不同饲喂方式对血清中炎症指标的影响
由表6可知,不同时间点组间血清中炎症指标的分析结果:在第3天,试验2、3组血清中IL-1β含量显著高于试验1组(P<0.05);在第0、9和15天,3组间血清IL-1β含量差异不显著(P>0.05)。在第0和9天,3组间血清中TNF-α含量差异不显著(P>0.05);在第3天,试验2、3组血清TNF-α含量显著高于试验1组(P<0.05);在第15天,试验1组血清TNF-α含量显著高于试验2组(P<0.05),与试验3组差异不显著(P>0.05)。在第0、3和15天,3组间血清中IFN-γ含量差异不显著(P>0.05);在第9天,试验2组血清IFN-γ含量显著高于试验1组(P<0.05),与试验3组差异不显著(P>0.05)。

表6 羔羊断奶应激期内不同饲喂方式对血清中炎症指标的影响
2.5 羔羊断奶应激期内不同饲喂方式对血清中肠道屏障指标的影响
由表7可知,不同时间点组间血清中肠道屏障指标的分析结果:在第3天,试验2组血清中D-乳酸含量显著高于试验1组(P<0.05),与试验3组差异不显著(P>0.05);在第9天,试验1组血清D-乳酸含量显著高于试验3组(P<0.05),与试验2组差异不显著(P>0.05);在第0和15天,3组间血清D-乳酸含量差异不显著(P>0.05)。在第0天,3组间羔羊血清中DAO活性差异不显著(P>0.05);在第3天,试验2、3组血清DAO活性显著高于试验1组(P<0.05);在第9天,试验1、2组血清DAO活性显著高于试验3组(P<0.05);在第15天,试验2组血清DAO活性显著高于试验1、3组(P<0.05)。在第0和15天,3组间羔羊血清中ET含量差异不显著(P>0.05);在第3天,试验2组血清ET含量显著高于试验1、3组(P<0.05);在第9天,试验3组血清ET含量显著高于试验1、2组(P<0.05)。

表7 羔羊断奶应激期内不同饲喂方式对血清中肠道屏障指标的影响
2.6 羔羊断奶应激期内不同饲喂方式对血清中免疫球蛋白含量的影响
由表8可知,不同时间点组间血清中IgA、IgG、IgM含量分析结果:在第0和3天,3组间血清中IgA含量差异不显著(P>0.05);在第9天,试验1组血清中IgA含量显著高于试验2组(P<0.05),与试验3组差异不显著(P>0.05);在第15天,试验1组血清中IgA含量显著高于试验2、3组(P<0.05)。在第0、3和9天,3组间血清中IgG含量差异不显著(P>0.05);在第15天,试验1组血清IgG含量显著高于试验2组(P<0.05),与试验3组差异不显著(P>0.05)。在第0和3天,3组间血清中IgM含量差异不显著(P>0.05);在第9天,试验1组血清中IgM含量显著高于试验2组(P<0.05),与试验3组差异不显著(P>0.05);在第15天,试验1、3组血清中IgM含量显著高于试验2组(P<0.05)。

表8 羔羊断奶应激期内不同饲喂方式对血清中免疫球蛋白含量的影响
同组在不同时间点血清中IgA、IgG、IgM含量的分析结果:在试验1组,各时间点间血清中IgA和IgG含量差异不显著(P>0.05);第15天血清中IgM含量显著低于第0天(P<0.05),与第3、9天差异不显著(P>0.05)。在试验2组,第0、3天血清中IgA含量显著高于第9、15天(P<0.05);第15天血清中IgG含量显著低于第0、3天(P<0.05),与第9天无显著差异(P>0.05);第9、15天血清中IgM含量显著低于第0天(P<0.05),与第3天无差异(P>0.05)。在试验3组,血清中IgG、IgM含量在不同时间点间差异不显著(P>0.05);第15天血清中IgA含量显著低于第0、3天(P<0.05),与第9天无显著差异(P>0.05)。
3 讨 论
3.1 断奶应激期内不同饲喂方式对羔羊生长性能的影响
断奶是羔羊生长过程中的重要阶段,对于羔羊生长性能有重要影响,其中体重是评价羔羊生长发育的重要指标。Chai等[4]研究断奶对湖羊生长性能的影响,发现30日龄断母乳羔羊在断哺乳1~10 d的ADG显著低于随母哺乳组。王世琴[5]研究断奶应激对羔羊生长性能的影响,发现与断奶前7 d相比,49日龄断奶羔羊的采食量在断奶后1~7 d升高,但ADG在断奶后1~7 d显著降低。柴建民等[6]研究发现,30日龄断母乳羔羊的ADG在断母乳1~10 d显著低于未断奶组,但采食量差异不显著。以上研究表明,断奶应激会影响幼畜的生长性能。本试验中,试验1组羔羊在第0~15天的ADG显著高于试验2组,与试验3组差异不显著,说明断奶应激使羔羊的生长性能下降。试验3组羔羊在第0~15天的ADG和DMI显著高于试验2组,试验1组羔羊在第0~15天的DMI显著低于试验3组,说明50% TMR+50%羔羊料能缓解断奶应激,也说明羔羊断奶后饲喂50% TMR+50%羔羊料能更有利于羔羊的健康和生长,可能是因为TMR中性洗涤纤维(NDF)含量远高于羔羊料,过多的NDF会使瘤胃快速充满而减少动物DMI[7],使饲料转化率降低,影响羔羊生长性能。
3.2 断奶应激期内不同饲喂方式对羔羊血清中COR和ACTH含量的影响
动物应激反应是由多种激素参与的过程,其中最重要的是糖皮质激素。糖皮质激素能够影响大多数器官和组织的生物学功能,并维持基础和应激相关的能量稳态。COR是由肾上腺皮质分泌的一种重要糖皮质激素,在动物机体内主要作用是促进组织中蛋白质和脂肪的分解。下丘脑-垂体-肾上腺(HPA)轴激活是应激时COR含量升高的主要调节机制,主要通过激活HPA轴刺激垂体前叶释放ACTH,促进肾上腺皮质分泌COR[8-10]。有研究报道,犊牛断奶应激会造成血清COR含量升高,COR可作为动物应激的生物标志物[11]。范超等[12]研究断奶应激对犊牛血清生化指标的影响,发现犊牛血清中COR含量在断奶后1 d上升,至断奶后14 d恢复正常水平。高艳霞等[13]研究断奶应激对犊牛血液生化、免疫和内分泌的影响,发现犊牛血液中的COR含量在断奶后1、3 d升高,断奶后5 d显著升高,断奶后7、14 d恢复正常水平。Ekiz等[14]研究不同断奶时间(45、75 d)对羔羊血浆COR含量的影响,结果发现2组羔羊的血浆COR含量在断奶后1 d显著升高,在断奶后2 d开始下降,直至恢复正常水平。以上研究表明,断奶应激会导致幼畜血液中COR含量升高,一般在断奶2周内恢复到正常水平。本试验中,试验2、3组羔羊60日龄断奶,断奶对血清中COR和ACTH含量无显著影响,说明断奶前补饲羔羊料可缓解断奶应激。试验2、3组羔羊血清中COR和ACTH的含量在断奶后2周内均呈先升高后降低的趋势,均在第3天达到最高水平,高于第9和15天,说明羔羊应激时血清中COR和ACTH含量变化一致,断奶应激通过激活HPA轴促进ACTH释放以提高COR的分泌[9-10],同时也说明断奶后1周内羔羊的应激较大。试验2、3组羔羊血清中COR和ACTH含量在第15天均低于断奶前,说明羔羊在断奶前(第0天)第1次采血时产生了应激,随着采血次数增多,羔羊对采血行为逐渐适应,应激减小。
试验2组羔羊在断奶后第3天血清中COR含量显著高于试验1组,试验3组与试验1、2组无显著差异,说明在哺乳期补饲羔羊料的基础上,断奶后饲喂50% TMR+50%羔羊料有助于缓解断奶应激。
3.3 断奶应激期内不同饲喂方式对羔羊血清中酶活性的影响
LDH是动物机体糖酵解途径中的重要酶,可催化丙酮酸转化为乳酸,应激条件下,在保证心脏等重要器官供氧的前提下,其他组织极可能在缺氧时采取无氧酵解方式供能,导致血液中LDH活性升高[12]。因此,通常将LDH作为反映机体能量代谢是否正常的应激指标。范超等[12]和高艳霞等[13]研究断奶应激对犊牛血清生化指标的影响,发现犊牛血清中LDH活性在断奶1 d后显著升高,在断奶后7、14 d恢复正常水平,说明断奶应激对犊牛能量代谢造成了影响。柴建民[15]研究断奶日龄对羔羊血清生化指标的影响,发现在10和30日龄断奶的羔羊血清中LDH活性在断奶后较对照组显著升高。本试验中,试验2、3组羔羊在第3、9天血清中LDH活性与试验1组无显著差异,但在第15天均低于试验1组,与以上研究结果不一致,说明在本试验条件下断奶没有影响到羔羊的能量代谢,试验1组血清LDH活性较高可能是因为总摄入能量不足,组织代谢过程中存在糖酵解,也说明随着泌乳期延长母羊乳汁质量下降,仅靠母乳或者补饲羔羊料不能满足羔羊生长发育的需要。试验2、3组羔羊血清LDH活性在断奶后2周内呈先升高后降低趋势,但无显著差异,说明60日龄断奶后饲喂TMR和羔羊料对能量代谢无显著影响。
CK是一种特异性激酶,其功能是催化三磷酸腺苷中的高能磷酸键,转移到肌酸分子生成磷酸肌酸而储存能量,当肌肉因应激受损时CK会迅速释放进入血液[16]。周向梅等[17]研究断奶应激对仔猪血清生化指标的影响,发现在断奶后7 d血清中CK活性升高。卢春莲等[18]研究运输应激对肉牛血液理化指标及免疫指标的影响,发现肉牛血清中CK活性在运输后显著上升。柴建民[15]研究断奶日龄对羔羊血清生化指标的影响,发现在10和30日龄断奶的羔羊血清中CK活性在断奶后较对照组显著升高。以上研究表明,应激会通过影响肌肉营养物质代谢提高血液中CK活性。本研究中,试验2、3组羔羊在断奶后3 d血清中CK活性显著高于试验1组,可能是因为断奶应激影响了肌肉营养物质代谢,导致CK迅速释放入血液中[16],在第9和15天,与试验1组无显著差异,说明在断奶后1周内对羔羊肌肉代谢影响较大。试验2、3组羔羊血清CK活性在断奶后2周内呈先升高后降低趋势,试验2组断奶羔羊在第9天血清CK活性最高,试验3组在断奶后第3天的血清CK活性最高,说明断奶应激期内饲喂50% TMR+50%羔羊料有助于降低应激对肌肉的影响。
ALP广泛存在于肝脏、骨骼等,与肝脏和骨骼的代谢密切相关。有研究表明,血清中的ALP可反映动物对蛋白质和脂类的代谢效率,其活性高低可反映动物的生长速率[19-20]。张庆坤等[21]研究不同断奶日龄对仔猪血清酶活性的影响,发现不同断奶日龄仔猪断奶7 d后血清ALP活性均下降。刘超良等[20]研究发现仔猪血清中ALP活性在断奶后7 d下降,断奶后14 d恢复正常水平。李冲[22]研究断奶应激对羔羊血清生化指标的影响,发现羔羊断奶后7 d血清中ALP活性显著降低。以上研究表明,断奶应激会降低幼畜血液中ALP活性。侯生珍等[23]研究蛋白质水平对早期断奶藏羔羊应激和免疫的影响,发现随着饲粮中蛋白质水平的升高,血清中ALP活性升高。本试验中,断奶影响了羔羊血清中ALP活性,在第15天试验2组的羔羊血清ALP活性显著低于试验1组,可能是因为断奶后全部更换饲粮对羔羊应激较大,在应激期内影响了羔羊生长发育[19-20],试验2组在断奶后3和9 d血清ALP活性与试验1组差异不显著,仅在第15天显著降低,可能是因为断奶应激对生长发育的影响在短期内不明显,也可能是因为TMR蛋白质含量较羔羊料低,蛋白质代谢效率低[24],断奶后长时间饲喂TMR对蛋白质代谢影响较大。试验1组羔羊血清中ALP活性在第0、3天显著低于第15天,可能是因为首次分组采血对羔羊产生了应激,随着应激逐渐缓解,血清ALP活性逐渐上升。试验2组羔羊虽然在各时间点间无显著差异,但血清中ALP活性低于试验3组,可能是因为试验2组内个体差异较大。试验3组羔羊在断奶后第3天血清ALP活性最低,第9天恢复到断奶前水平,说明断奶应激期内饲喂50% TMR+50%羔羊料更有利于羔羊生长发育。
ALT主要存在于肝脏中,AST存在于更多组织中,主要是心脏和骨骼肌,其次是肝脏,可作为检测组织损伤的敏感指标[1]。ALT和AST作为非特异性细胞内功能酶,正常时在血清中的活性较低,但当组织细胞受损或细胞膜通透性升高时,会造成血清中ALT和AST活性升高[1,25]。张庆坤等[21]研究不同断奶日龄对仔猪血清酶活性的影响,结果发现14日龄断奶仔猪断奶后7 d血清中ALT和AST活性显著升高。周向梅等[17]研究仔猪断奶应激对血清生化指标的影响,发现血清AST活性断奶后7 d时比断奶时高,ALT活性变化不显著。范超等[12]研究断奶应激对奶水牛犊牛血清生化指标的影响,发现与断奶前1 d相比,犊牛血清中ALT活性在断奶后1、14 d显著升高,血清AST活性差异不大。以上研究说明,断奶应激会影响幼畜血清中ALT和AST的活性。本试验中,断奶应激对羔羊血清中ALT活性无显著影响,说明羔羊60日龄断奶不会影响血清中ALT活性,与周向梅等[17]研究结果类似。断奶应激显著影响了血清AST活性,且在断奶后15 d羔羊血清中AST活性最高,试验2组羔羊在第15天血清中AST活性最高,显著高于第3天,说明断奶应激对组织的损伤在2周内难以恢复。试验2组羔羊血清中AST活性在第3和15天高于试验1组和3组,说明断奶后饲喂TMR相对于饲喂50% TMR+50%羔羊料对组织损伤较大。
3.4 断奶应激期内不同饲喂方式对羔羊血清中炎症因子指标的影响
IL-1β和TNF-α是由单核巨噬细胞产生的促炎细胞因子,可诱导急性期反应,对中性粒细胞和巨噬细胞有刺激作用,在组织异常合成可导致发热,造成炎症反应[11]。TNF-α与IL-1β具有协同作用,通过降低局部血流速度使血液聚集在渗漏的血管中,从而增加淋巴细胞和巨噬细胞的边缘化,然后白细胞在细胞因子和趋化因子的引导下外渗到炎症组织中[26]。Kim等[27]研究发酵豆粕部分替代豆粕对断奶犊牛应激指标的影响,发现2组犊牛断奶后血清中TNF-α和IL-1β含量均升高,表明断奶作为应激源可能会诱导促炎细胞因子的产生。Kim等[28]研究断奶应激对犊牛血清生化指标的影响,发现犊牛血清TNF-α含量在断奶后3 d显著升高。Zhang等[11]研究发现,羔羊断奶应激会造成血清IL-1β和TNF-α含量显著增加。以上研究表明,断奶应激会导致幼畜发生炎症反应。本试验中,试验2、3组羔羊在断奶后3 d血清中TNF-α和IL-1β含量显著高于试验1组,第9天与试验1组无显著差异,说明断奶会导致羔羊发生炎症反应,且试验2、3组血清TNF-α和IL-1β含量均在第3天最高,说明在断奶后1周内炎症发应最大,与以上研究结果一致。本研究中,试验1组羔羊在第15天血清中TNF-α含量高于试验2、3组,可能是因为试验期间母羊频繁挤奶,造成部分母羊乳房出现炎症,羔羊采食有炎症的母乳造成了血清中TNF-α含量升高。
IFN-γ是由T淋巴细胞和自然杀伤(NK)细胞产生的细胞因子,能维持和促进T淋巴细胞的增值,参与细胞免疫应答,进而调节机体免疫功能,当机体受到刺激时会导致IFN-γ含量升高[29-31]。王莉梅等[32]研究运输应激对安格斯牛血清生化指标的影响,发现运输后的血清IFN-γ含量显著高于运输前。高艳霞等[13]研究断奶应激对犊牛血清生化指标的影响,发现犊牛断奶后血清IFN-γ含量有上升趋势,说明犊牛断奶应激刺激了T淋巴细胞做出应答。以上研究表明,应激会影响到动物的免疫功能。本试验中,试验2组在第9天血清中IFN-γ含量高于试验1、3组,且试验2组在第9、15天的血清IFN-γ含量显著高于第0和3天,说明饲喂TMR比50% TMR+50%羔羊料对羔羊免疫力影响较大,且对免疫力的影响在2周内难以恢复。
3.5 断奶应激期内不同饲喂方式对羔羊血清中肠道屏障指标的影响
D-乳酸是由肠道微生物产生的,哺乳动物不会产生D-乳酸,也不具备代谢D-乳酸的酶,当肠黏膜受损、通透性增加时,肠微生物产生的D-乳酸会进入血液,导致血清中D-乳酸含量增加,故血清D-乳酸含量可反映肠道健康状态[33]。DAO是由肠上皮分泌的细胞酶,存在于肠血管上层的绒毛细胞中,当肠黏膜受损时,DAO被释放进入血液,因此血清DAO活性常被作为肠道完整性的标志物[34-35]。关于断奶应激对幼畜血清中肠道屏障指标的研究主要集中在单胃动物上,反刍动物较少。栾兆双等[33]研究断奶应激对仔猪肠形态、黏膜屏障的影响,结果表明与哺乳组相比,断奶组仔猪血清D-乳酸含量和DAO活性在断奶后1、3 d均显著升高。钱仲仓等[36]研究早期断奶对仔猪应激、肠形态和肠通透性的影响,仔猪断奶后血清D-乳酸含量和DAO活性断奶后1 d迅速增加。Xiao等[37]研究断奶应激对仔猪肠道形态和通透性的影响,结果发现仔猪断奶后3、7 d血清DAO活性显著升高。胡彩虹等[38]研究发现基础饲粮组仔猪断奶后第7、15天血浆中D-乳酸含量和DAO活性显著高于哺乳组。以上研究表明,断奶应激会引起仔猪黏膜屏障受损,使肠通透性增加。本试验中,试验2、3组羔羊在断奶后血清中D-乳酸含量和DAO活性均呈先升高后降低趋势,且均在断奶后第3 天达到最大值,在第15天恢复到正常水平,说明断奶应激造成的肠道损伤在2周内可以恢复。试验1组羔羊在第9天血清中D-乳酸含量和DAO活性高于试验3组,可能是因为试验期间母羊频繁挤奶,部分母羊乳房出现炎症,且挤奶应激影响了母羊胃肠道健康,羔羊采食有炎症的母乳造成血清D-乳酸含量和DAO活性升高。试验2组羔羊在第3天血清中D-乳酸含量和DAO活性显著高于试验1组,在第15天血清DAO活性显著高于试验1、3组,说明断奶后饲喂TMR相较于50% TMR+50%羔羊料对羔羊肠道损伤较大,使肠道通透性变大。
ET是革兰阴性菌细胞壁的脂多糖成分,大量存于机体肠腔内,对宿主是有害的,肠道是抵御ET的第1道防线,正常情况下ET难以大量进入血循环,当肠道屏障功能障碍时,大量ET可通过受损屏障进入肝脏,当超过肝脏代谢能力时,ET会进入血循环,因此血液中ET含量可作为检测肠道屏障完整性的指标[39-40]。庞敏[41]研究发现仔猪断奶前期血清ET含量显著升高,说明断奶应激损伤了肠道黏膜。张志浩[42]研究氧化应激对肉鸡肠道黏膜屏障功能的影响,结果表明氧化应激使血浆中ET含量显著增加。本试验中,试验2、3组羔羊在断奶后血清中ET含量呈先升高后降低趋势,均在第15天血清中ET含量恢复到正常水平,说明断奶应激造成的肠道损伤在2周内可以恢复。试验2组血清ET含量在第3天显著高于其他2组,且在第3天达到最高水平;试验3组在第9天显著高于其他2组,且在第9天达到最高水平,说明断奶后饲喂TMR对肠道损伤较大,饲喂50% TMR+50%羔羊料对肠道损伤较缓和。
3.6 断奶应激期内不同饲喂方式对羔羊血清中免疫球蛋白含量的影响
免疫球蛋白含量在一定程度上能够反映幼龄反刍动物的免疫能力,主要包括IgG、IgA、IgM,其中以IgG含量最高,在机体免疫调节中起主要作用[43-44]。高艳霞等[13]研究断奶应激对犊牛血清免疫的影响,发现断奶后血清中免疫球蛋白含量均有不同程度的下降,表明断奶应激导致犊牛免疫力降低。赵会利等[45]研究发现断奶应激导致犊牛血清IgG含量有所下降。王莉梅等[32]研究运输应激对安格斯牛血清免疫球蛋白含量的影响,发现运输后血清中IgG、IgA含量显著降低,IgM含量呈降低趋势。以上研究结果说明,应激会导致幼畜血液中免疫球蛋白含量下降。本试验中,试验2、3组羔羊血清中IgG、IgA、IgM含量均在断奶后15 d达到最低,说明羔羊断奶后2周内免疫力未得到较好恢复,由此在断奶应激期内提高羔羊饲养管理水平对缓解断奶应激、促进羔羊生长发育至关重要。试验2组羔羊血清IgA和IgM含量在第9和15天显著低于试验1组,试验3组在第9天与试验1组无显著差异,说明断奶后饲喂TMR对羔羊免疫力影响较大,断奶后饲喂50% TMR+50%羔羊料有助于缓解断奶应激,对羔羊免疫力影响不大,可能是因为TMR中NDF含量高于颗粒料,较高的NDF含量会使瘤胃快速充满,减少营养成分的摄入,影响动物的生长发育,进而可能会影响自身免疫力。
4 结 论
断奶应激期内不同饲喂方式对羔羊断奶应激影响较大,在哺乳期补饲羔羊料的基础上,断奶后饲喂50% TMR+50%羔羊料可以缓解断奶应激,更有利于羔羊健康和生长发育。
